Разные виды зубов и их особенности

Что такое зубы?
Образования, находящиеся в альвеолярном отростке челюсти, сформированные из твердых тканей, называются зубами. Вопреки расхожему мнению, они состоят не из кости, а намного более прочных компонентов. Помимо основной функции – пережевывать пищу, они выполняют еще и целый ряд других. Например, без зубов наша речь становится непонятной, возникают многочисленные дефекты. Также зубы опосредованно помогают социализации, ведь улыбка трактуется членами общества как настроенность на диалог, открытость и дружелюбие.
Однако без дробления пищи в ротовой полости нашим пищеварительным органам было бы сложно ее переработать.
В зависимости от нагрузки, которая приходится на каждый зуб, со временем они эволюционировали до нынешних форм.
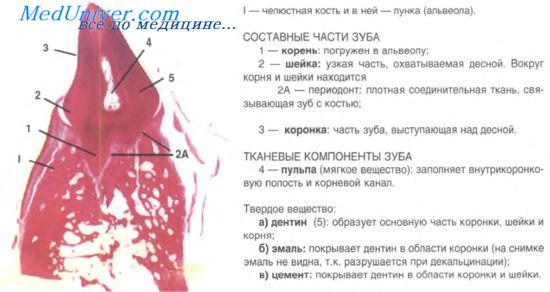
Составные компоненты зуба
Каждый зуб, вне зависимости от функционального назначения, состоит из следующих тканей:
Анатомическое строение предполагает условное разделение зуба на:
Корень находится глубоко в челюсти, его прикрывает десна. Местоположение шейки – между корнем и коронкой. И наконец, та часть, которую мы и называем зубом, в профессиональной среде имеет терминологическое определение – коронка. Она покрыта эмалью – самой прочной тканью организма – и именно на нее припадает основная функциональная нагрузка.
У коронки различают несколько типов поверхностей:
Зуб держится в десне за счет периодонта, цемента корня, десны и стенки зубной альвеолы. Все это составляет зубной орган, который вместе с участком челюсти, в котором находится непосредственно зуб, получил название зубочелюстного сегмента.
Классификация видов зубов
Правильное количество зубов у взрослого – 32. Стоматологи их делят на 4 группы, каждая из которых имеет свое название, форму, строение и функции.
Такой разновидности зубов, как глазные, в профессиональной среде не выделяют. Это исключительно бытовое название, которое, скорее всего, происходит из-за того, что малейшая их патология вызывает сильную боль, которая «стреляет» в глаз. При лечении приходится ставить особенно сильную анестезию, чтобы процедура проходила безболезненно.
Зубы молочные и коренные
В отличие от многих представителей животного мира, у которых зубы меняются несколько раз в течение жизни, у человека этот процесс происходит один раз – в возрасте от 5 до 12 лет.
Критерии нумерации зубов
Стоматологи всего мира давным-давно пронумеровали все наши зубы для облегчения работы. Считать взрослые зубы начинают со средины верхней челюсти влево и вправо. Это так называемая система Виола, именно ее традиционно используют в наших широтах.
Для молочных существует своя нумерация – от левого верхнего по часовой стрелке.
Для лицевых хирургов и ортодонтов создана квадратно-цифровая система.
Создана и так называемая универсальная система, при которой зубы получают буквенное, цифровое обозначение и ориентировку относительно сегмента в челюсти.
Аномалии в росте зубов
Стоматологической практике известны самые разнообразные отклонения, связанные с ростом зубов. Случается, что у человека появляется больше зубов, чем предусмотрено природой. Это называется полиодонтией. Также отмечены случаи отсутствия замены молочного зуба на постоянный.
Лишние зубы чаще всего приходится удалять, поскольку они нарушают правильный зубной ряд. Если их оставить, возможен целый ряд осложнений. К ним относятся:
Помимо этой неправильности, довольно часто встречается макродентия, то есть зубы более крупного размера, чем должны быть. Также известны случаи микродентии, то есть зубы меньшего размера, чем нужно по стилистическим показателям этого человека.
Может быть изменена форма коронки зуба, когда вырастают перекрученные, бочкообразные, и других аномальных форм зубы. Отдельные из них может исправить только длительное лечение у ортодонта.
Что такое жевательно-речевой аппарат
Чтобы человек мог разговаривать и жевать, эволюция создала целую систему, слаженная работа которой и обеспечивает выполнение названных функций. Эта система состоит из:
Все эти органы связаны между собой и какие-либо отклонения в работе одних нарушают слаженные действия других.
Прикус – взаимодействие между зубами верхней и нижней челюсти
Выполнение жевательно-речевым аппаратом своих функций затруднено, а то и невозможно при нарушениях окклюзии. Это понятие означает смыкание верхних и нижних зубов. Правильными окклюзиями считаются следующие:
Нарушения прикуса – довольно частая аномалия. Ее появление обусловлено генетически, влиянием вирусных инфекций, патологиями, которые перенесла женщина во время беременности.
В норме верхние зубы на 1/3 прикрывают нижние. При правильном прикусе нагрузка, возникающая при жевании, распределяется равномерно между всеми зубами. Также люди с нормативным прикусом правильно выговаривают все звуки. Однако случаются и следующие сбои:
Здоровье зубов – это здоровье всего организма
Стоматологи всего мира давно пришли к выводу, что болезни зубов приводят к самым различным недугам, в остальном казалось бы, организма. Например, если больные зубы, неправильный прикус не позволяют качественно разжевать пищу – возникают заболевания желудочно-кишечного тракта. Помимо этого, возможно появление:
Очевидно, что заболевания зубов имеют крайне серьезные последствия, что делает актуальным вопрос посещения специалиста соответствующего профиля раз в полгода, проведение профилактических мероприятий и получение надлежащего лечения.
Что такое дифференцированные зубы
Зубы участвуют в механической обработке пищи: плоские резцы и клыки конической формы откусывают пищу, малые и большие коренные зубы с коронками кубической формы и жевательными буграми перетирают ее при еде. Зубы имеют важное значение для артикуляции.
Гисто- и органогенез зубов. У человека различают две смены зубов — выпадающие, или молочные (20), и постоянные (32). Развитие молочных зубов начинается в конце 2-го месяца эмбриогенеза. В это время эпителий ротовой полости врастает в виде зубной пластинки в подлежащую мезенхиму. На передней поверхности зубной пластинки возникают эпителиальные зубные почки по числу закладок зубов, вокруг которых возникает уплотнение мезенхимных клеток — зубные мешочки.
Взаимодействие двух эмбриональных зачатков приводит к изменению формы зубной пластинки — она постепенно трансформируется в структуру в виде бокала, внутри которого концентрируются мезенхимные клетки в форме сосочка. Последний оказывает индуктивное влияние на дифференцировку клеток эпителиального зубного бокала, в котором топографически выделяются внутренний и наружный эмалевый эпителий и клетки промежуточного слоя. Внутренний эмалевый эпителий обращен к мезенхимному сосочку, наружный образует «стенку» зубного бокала и тонкой клеточной ножкой остается некоторое время связанным с эпителием ротовой полости; клетки промежуточного слоя располагаются между двумя первыми, приобретают звездчатую форму и оттесняются друг от друга накапливающейся здесь жидкостью.
Внутренний эмалевый эпителий отделяется от мезенхимного сосочка базальной мембраной. Его клетки дифференцируются в энамелобласты (амелобласты) — клетки-образователи эмали. Образование базальной мембраны индуцирует дифференцировку расположенных рядом мезенхимных клеток в одонтобласты (дентинобласты). Последние, в свою очередь, влияют на развитие энамелобластов.
Энамелобласты имеют вытянутую цилиндрическую форму, в них постепенно происходит перемещение ядер из базальной части клеток в апикальную, поскольку в базальных частях клеток формируются эмалевые призмы, слой которых составляет эмаль зуба. Начинается кальцинация эмали. Каждый энамелобласт вырабатывает одну эмалевую призму.
Клетки, прилежащие к энамелобластам, — одонтобласты — начинают секретировать дентин во встречном с образованием эмали направлении. По мере развития зуба массы эмали и дентина увеличиваются, и ряды клеток удаляются друг от друга. Энамелобласты при этом отодвигаются наружу, а одонтобласты — внутрь развивающегося зуба. К моменту прорезывания молочных зубов ядросодержащие части энамелобластов редуцируются, остаются лишь тесно прилежащие друг к другу эмалевые призмы, покрытые кутикулой, образованной остатками промежуточных звездчатых клеток и наружного эмалевого эпителия. Последние постепенно уменьшаются в размерах и дегенерируют; клеточная ножка, связывающая зачаток зуба с эпителием полости рта, фрагментируется и полностью исчезает.
Зубной зачаток оказывается погруженным в костную ткань челюсти. Формирование эмали и дентина распространяется от вершины будущего зуба к боковым поверхностям. Клетки зубного мешочка дифференцируются в цементобласты, и незадолго до прорезывания зуба в области будущих корней формируют цемент. Клетки центральных участков мезенхимных сосочков образуют пульпу зуба — внутреннюю рыхлую соединительную ткань зуба, богатую сосудами. Из клеток наружного слоя мезенхимного зубного мешочка образуется зубная связка (периодонт), соединяющая зуб с альвеолой челюсти. Зубные альвеолы формируются из окружающей зубные зачатки мезенхимы параллельно с формированием зубов. Таким образом, в составе зуба эмаль имеет эпителиальную природу.
Все остальные части зуба (дентин, цемент, пульпа зуба), а также связочный аппарат — это производные мезенхимы.
Закладка постоянных зубов происходит на 4-5-м месяцах эмбриогенеза, когда от зубной пластинки начинают формироваться вторые эмалевые зачатки. Развитие их происходит принципиально так же, как и молочных зубов.
Что такое дифференцированные зубы
BMP — костный морфогенетический белок
DPSC — стволовые клетки пульпы взрослого зуба
ED — эмбриональный день
ERM — эпителиальная сеть Малассе
FGF — фактор роста фибробластов
Hedgehog — название сигнального пути
HERS — эпителиальное корневое влагалище Гертвига
iPS-клетки — индуцированные плюрипотентные стволовые клетки
PDLSC — стволовые клетки периодонтальной связки
SCAP — стволовые клетки апикального сосочка
SHED — стволовые клетки молочных зубов
Shh — sonic hedgehog, ген, кодирующий белок семейства hedgehog
TGFβ — трансформирующий фактор роста β
TNF — фактор некроза опухолей
WNT — белок сигнального пути
Зубы обеспечивают важные аспекты здоровья и качества жизни индивида, такие как пережевывание и глотание пищи, а также воспроизведение речи. Утрата зуба в результате кариеса и его осложнений, заболеваний пародонта или травматического повреждения приводит к нарушению этих функций и ухудшению эстетического вида. Для устранения последствий, вызванных потерей зуба, используют заместительную терапию с помощью искусственных коронок или зубных имплантатов.
Принципиально новые и более эффективные возможности для восстановления утраченного или поврежденного зуба могут дать технологии регенеративной стоматологии. Регенеративная стоматология предполагает выращивание зуба с помощью эмбриональных, дентальных или индуцированных плюрипотентных стволовых (iPS) клеток или «конструирование» зуба с помощью матриц, имитирующих экстраклеточный матрикс, и предшественников зубных клеток [1—7]. Однако пока эти технологии еще далеки от воспроизведения функционально и эстетически полноценного зуба. Вместе с тем становится все более очевидным, что вырастить полноценный зуб можно только по законам естественного одонтогенеза. В настоящем обзоре будут рассмотрены клеточные и молекулярные механизмы естественного развития зуба.
Зубной ряд и анатомия зубов
Существует четыре типа зубов: резцы, клыки, премоляры и моляры. Резцы откусывают, клыки размельчают, а премоляры и моляры перетирают пищу. Человек имеет зубы всех типов, а мыши только резцы и моляры (рис. 1) [8]. Люди имеют два поколения зубов: временные или «молочные» и постоянные. В отличие от людей у рептилий, рыб и амфибий зубы могут заменяться большое количество раз [9].
 Рис. 1.Зубнойрядчеловека (а) имыши (б). Четыре типа зубов: резцы, клыки, премоляры и моляры. Адаптирован с рисунка из Jussila and Thesleff, 2012.
Рис. 1.Зубнойрядчеловека (а) имыши (б). Четыре типа зубов: резцы, клыки, премоляры и моляры. Адаптирован с рисунка из Jussila and Thesleff, 2012.
Зуб состоит из коронки, шейки и корня (рис. 2). Коронка — это видимая часть зуба, которая покрыта эмалью до цементно-эмалевого соединения или «шейки» зуба. Корень расположен ниже шейки зуба и покрыт цементом. Под эмалью и цементом находится дентин, который формирует внутреннюю пульпарную камеру в коронке и каналы в корнях. Резцы, клыки и премоляры, кроме первых премоляров верхней челюсти, имеют один корень, верхние первые премоляры и нижние моляры — два, а верхние моляры — три корня. Гистологически в зубе различают четыре вида тканей: эмаль, дентин, цемент и пульпа.
 Рис. 2. Анатомия зуба. 1 — зуб; 2 — эмаль; 3 — дентин; 4 — пульпа; 5 — пульповая камера; 6 — корневая пульпа; 7 — цемент; 8 — коронка; 9 — бугор; 10 — борозда; 11 — шейка; 12 — корень; 13 — фуркация; 14 — верхушка корня; 15 — апикальное отверстие; 16 — десневой желобок; 17 — периодонт; 18 — десна; 19 — свободная; 20 — маргинальная; 21 — альвеолярная; 22 — периодонтальная связка; 23 — альвеола; 24 — сосуды и нервы; 25 — дентальные; 26 — периодонтальные; 27 — альвеолярные. Автор — И.Ю. Малышев.
Рис. 2. Анатомия зуба. 1 — зуб; 2 — эмаль; 3 — дентин; 4 — пульпа; 5 — пульповая камера; 6 — корневая пульпа; 7 — цемент; 8 — коронка; 9 — бугор; 10 — борозда; 11 — шейка; 12 — корень; 13 — фуркация; 14 — верхушка корня; 15 — апикальное отверстие; 16 — десневой желобок; 17 — периодонт; 18 — десна; 19 — свободная; 20 — маргинальная; 21 — альвеолярная; 22 — периодонтальная связка; 23 — альвеола; 24 — сосуды и нервы; 25 — дентальные; 26 — периодонтальные; 27 — альвеолярные. Автор — И.Ю. Малышев.
Эмаль — самая твердая ткань в организме человека. Она состоит из 96% минеральных веществ и 4% воды и органических веществ. Основной элемент эмали — минерал гидроксиапатит. Благодаря этому эмаль обеспечивает механическую прочность и защищает зуб от действия кислот, находящихся в пище, и продуктов жизнедеятельности микроорганизмов. В отличие от дентина в эмали отсутствуют коллагеновые волокна, но присутствуют уникальные белки амелогенин и энамелин. Эмаль формируют амелобласты. В ходе одонтогенеза амелобласты покрывают поверхность зуба, синтезируя продукты эмали. В результате происходит минерализация эмали и кристаллизация кальциево-фосфатных соединений с образованием кристаллической решетки. Однако перед прорезыванием зуба эти клетки подвергаются апоптозу. Поэтому эмаль взрослого зуба не содержит клеток, является нежизнеспособной, лишенной чувствительности и не обладает способностью к регенерации или замещению после повреждения [10].
Дентин мягче и менее минерализован по сравнению с эмалью. Дентин состоит из 70% неорганических веществ, 20% органических и 10% воды. В состав дентина входят белки (коллаген I типа, дентин фосфопротеин и дентин сиалопротеин) [11]. Дентин формирует основу зуба и служит опорой для эмали. Дентин секретируется одонтобластами пульпы. Дентин имеет каналы диаметром от 2,5 мкм до 900 нм, которые расходятся от пульпарной камеры к цементу или эмалевой границе. В эти каналы проникают отростки одонтобластов. В отличие от эмали дентин является чувствительной тканью и обладает способностью к регенерации. Это связано с тем, что в ответ на механические повреждения одонтобласты или мезенхимальные клетки пульпы зуба могут дополнительно откладывать дентин [10].
Цемент мягче, чем дентин и эмаль. Цемент состоит из 45% минеральных компонентов, 33% органических веществ и 22% воды. Он обеспечивает прикрепление периодонтальной связки к зубу. Цемент секретируется цементобластами. По химическому составу и структуре цемент напоминает минерализованную грубоволокнистую соединительную ткань, это наименее минерализованная твердая ткань зуба.
Пульпа — центральная часть зуба, которая окружена дентином, заполнена мягкой соединительной тканью и содержит фибробласты, макрофаги, Т-лимфоциты, а также одонтобласты, которые располагаются на границе между пульпой и дентином. Через отверстия в апексе корня в зуб поступают кровеносные сосуды и нервы. Пульпа зуба выполняет важные функции, такие как продукция дентина, питание дентина и опора для нервных отростков и кровеносных сосудов, которые обеспечивает чувствительность и питание зуба.
Зубы прикрепляются и удерживаются в челюсти при помощи комплекса специализированных тканей, которые совокупно называются пародонтом. Пародонт состоит из цемента (также является частью зуба), периодонтальной связки, костной альвеолы и десны. Периодонтальная связка связывает корень зуба и костную альвеолу. Периодонтальная связка состоит из коллагеновых волокон и содержит остеобласты, остеокласты, фибробласты, макрофаги, цементобласты и эпителиальные островки Малассе (ERM) (также являются частью зуба). Периодонтальная связка также участвует в образовании и резорбции альвеолярной кости, в формировании чувства давления и в прорезывании зуба. Альвеолярная кость — это часть челюстной кости, которая формирует «лунку» вокруг корня зуба. Альвеолярную кость образуют остеобласты, а разрушают остеокласты. И, наконец, десна — это часть слизистой оболочки полости рта и самый периферический отдел пародонта, который покрывает кость. Различают следующие части десны: свободную десну и прикрепленную. Свободная десна покрывает пришеечный участок зуба. Граница между свободной и прикрепленной десной проходит на уровне эмалево-цементного соединения. Прикрепленная десна — это часть десны, которая сращена с надкостницей альвеолярной кости и корневым цементом.
Морфогенез и клеточная дифференцировка в течение роста зуба
Зуб является эктодермальным органом, т. е. развивается из тканей, сформированных зародышевым листком эктодермы. Клетки эктодермы, делясь и дифференцируясь, образуют две структуры, необходимые для развития зуба: дентальный эпителий и нервный гребень. Нервный гребень позже преобразуется в мезенхиму. Развитие зуба инициируется и регулируется взаимодействием между эпителием и мезенхимой [4, 7, 12, 13]. Самым ранним морфологическим признаком формирования зуба является образование первичной зубной (дентальной) пластинки (ламины), подковообразной полосы утолщения эпителия, которая отмечает будущий зубной ряд [14—17]. Процесс развития зуба делят на стадии плакоды, почки, колпачка, колокола и развития корня (рис. 3) [8]. Наиболее детально эти стадии изучены на экспериментальных мышах.
 Рис. 3. Стадии развития зуба: стадия плакоды, стадия почки, стадия колпачка, стадия колокола, стадия развития корня и прорезывания. Формирование зуба начинается на месте дентальной ламины, которая состоит из инвагинированного эпителия, образовавшегося из незрелого ротового эпителия, и мезенхимальных клеток, образовавшихся из клеток нейронального креста. У мышей на ED10−11 из дентальной ламины формируется зачаток зуба (стадия плакоды). На ED13.5—14.5 на стадии колпачка формируется первичный эмалевый узел, а на ED16.5—18.5 на стадии колокола — вторичные эмалевые узлы, которые формируют бугорки коронки зуба. На этой же стадии эпителиальные и мезенхимальные клетки зародыша зуба дифференцируются в клетки, формирующие ткани зуба, а именно амелобласты, одонтобласты и клетки дентального фолликула. Амелобласты и одонтобласты продуцируют эмаль и дентин соответственно границе между эпителием и мезенхимой. Клетки дентального фолликула дифференцируются в клетки тканей периодонта, такие как периодонтальная связка, цемент и альвеолярная кость. Адаптирован с рисунка из Jussila and Thesleff, 2012.
Рис. 3. Стадии развития зуба: стадия плакоды, стадия почки, стадия колпачка, стадия колокола, стадия развития корня и прорезывания. Формирование зуба начинается на месте дентальной ламины, которая состоит из инвагинированного эпителия, образовавшегося из незрелого ротового эпителия, и мезенхимальных клеток, образовавшихся из клеток нейронального креста. У мышей на ED10−11 из дентальной ламины формируется зачаток зуба (стадия плакоды). На ED13.5—14.5 на стадии колпачка формируется первичный эмалевый узел, а на ED16.5—18.5 на стадии колокола — вторичные эмалевые узлы, которые формируют бугорки коронки зуба. На этой же стадии эпителиальные и мезенхимальные клетки зародыша зуба дифференцируются в клетки, формирующие ткани зуба, а именно амелобласты, одонтобласты и клетки дентального фолликула. Амелобласты и одонтобласты продуцируют эмаль и дентин соответственно границе между эпителием и мезенхимой. Клетки дентального фолликула дифференцируются в клетки тканей периодонта, такие как периодонтальная связка, цемент и альвеолярная кость. Адаптирован с рисунка из Jussila and Thesleff, 2012.
Cтадия плакоды характеризуется появлением на дентальной ламине локального утолщения эпителия, называемого плакодой. У мышей плакода формируется на 10—11-й день эмбрионального развития (ED10−11). На ED11,5 плакода погружается в мезенхиму и трансформируется в так называемую почку, которая дает рост одному зубу.
На стадии почки дентальный эпителий разделяется на два клеточных пула: периферические базальные клетки, контактирующие с базальной мембраной, и центрально расположенные клетки звездчатого ретикулума (см. рис. 3). Эти два вида клеток будут формировать эпителиальный компонент ниши стволовых клеток в растущих зубах. На этой стадии мезенхима начинает конденсироваться, окружает эпителиальную почку и приобретает форму полумесяца.
На стадии колпачка, которая у мышей начинается на ED13.5−14.5, дентальная мезенхима продолжает конденсироваться вокруг почки и разделяется на два клеточных пула: центральный зубной сосочек и периферический зубной фолликул. Зубной сосочек позже будет окружен зубным эпителием и даст начало пульпе зуба и одонтобластам, а периферический зубной фолликул даст начало цементобластам, остеобластам и фибробластам. Цементобласты будут формировать цемент зуба, остеобласты — альвеолярную кость, а фибробласты — периодонтальную связку.
Во время стадии колпачка и дальше на стадии колокола, которая у мышей начинается на ED16.5—18.5, формируется коронка зуба. Этот процесс регулируют первичные и вторичные эмалевые узлы. Эмалевые узлы определяют места бугров зуба [18]. Форма коронки фиксируется благодаря тому, что на эпителиально-мезенхимальной границе эпителиальные клетки дифференцируются в амелобласты, а мезенхимальные — в одонтобласты. Амелобласты секретируют продукты эмали, а одонтобласты — дентина.
На стадиях колпачка и колокола эпителий уже начинает окружать подлежащую мезенхиму и формировать так называемые цервикальные петли (см. рис. 3). Слой базальных эпителиальных клеток петли, ограничивающий зубной сосочек, назвали внутренним эмалевым эпителием. Внутренний эмалевый эпителий дифференцируется в амелобласты. Часть базальных эпителиальных клеток петли, контактирующая с зубным фолликулом, назвали наружным эмалевым эпителием. Центр петли заполнен клетками звездчатого ретикулума. Цервикальные петли сохраняются в постоянно растущих резцах грызунов и образуют ниши взрослых стволовых клеток [19—21], которые обеспечивают рост зубов.
Стадия развития корня начинается после образования коронки зуба, когда клетки внутреннего эмалевого эпителия перестают дифференцироваться в амелобласты. На этой стадии в непостоянно растущих зубах (включая зубы человека) цервикальные петли модифицируются. Центральная часть петли исчезает, оставляя двойной слой базального эпителия, известного как эпителиальное корневое влагалище Гертвига (HERS). HERS пролиферирует и мигрирует вглубь мезенхимы, индуцируя дифференцировку одонтобластов, формирующих дентин корня. HERS имеет ограниченный потенциал роста, который определят длину корня. После формирования дентиновой основы корня происходит дезинтеграция HERS с образованием эпителиальной сети Малассе (ERM). Это позволяет клеткам зубного фолликула контактировать с дентином и дифференцироваться в цементобласты, продуцирующие цемент на поверхности корня. Из клеток зубного фолликула, как указано выше, также образуются фибробласты и остеобласты, которые формируют периодонтальную связку и альвеолярную кость соответственно [22].
Одонтогенная компетентность и сигнальные центры развития зуба
Формирование зуба регулирует последовательный процесс реципрокных взаимодействий между эпителиальными и мезенхимальными клетками (см. рис. 3, 4). Чтобы из этих клеток развился именно зуб, а не другая эктодермальная структура (слюнные железы или крипты кишечника), эти клетки должны обладать одонтогенной компетентностью, т. е. способностью формировать зуб. Несмотря на то, что уже определены более 200 генов, вовлеченных в развитие зуба (http://bite-it.helsinki.fi), генетическая основа одонтогенности до конца не понятна. Показано, что в механизмах одонтогенности и развития зуба важную роль играют эпителиальные сигнальные центры. Известны четыре вида таких центров: дентальная пластинка, плакода, первичные и вторичные эмалевые узлы. Сигнальные центры активируют Eda-, TGFβ-, BMP-, WNT-, FGF- и Hedgehog-зависимые пути, которые обеспечивают взаимодействие между эпителием и мезенхимой (рис. 4) [23—25]. Активность передачи сигнала по этим путям контролируют модуляторы, такие как ингибиторы ВМР (фоллистатин и эктодин/Sostdc1) [26, 27] и ингибиторы FGF (Sprouty) [28].
 Рис. 4. Последовательный и взаимный сигналинг между эпителием и мезенхимой регулирует экспрессию специфических факторов. Эта схема иллюстрируют пошаговый процесс с повторным использованием одних и тех же сигнальных путей. В этой схеме указаны не все пути и отсутствуют модуляторы сигналинга. Аббревиатуры со всеми заглавными буквами обозначают белки (BMP-4 и др.), а с первой прописной и остальными строчными — гены (Pitz-2 и др.). Автор И.Ю. Малышев.
Рис. 4. Последовательный и взаимный сигналинг между эпителием и мезенхимой регулирует экспрессию специфических факторов. Эта схема иллюстрируют пошаговый процесс с повторным использованием одних и тех же сигнальных путей. В этой схеме указаны не все пути и отсутствуют модуляторы сигналинга. Аббревиатуры со всеми заглавными буквами обозначают белки (BMP-4 и др.), а с первой прописной и остальными строчными — гены (Pitz-2 и др.). Автор И.Ю. Малышев.
В эпителии дентальной пластинки млекопитающих обнаружена экспрессия генов Shh и Pitx2 [29—32]. Shh необходим для инициации роста зуба у мышей, а также для дифференцировки дентального эпителия в амелобласты [33—35], а Pitx2 необходим для развития зубов и у мышей, и у людей [36, 37]. В дентальной пластинке также экспрессируются лиганды WNT [38], которые регулируют экспрессию FGF8 [39]. Доказано, что WNT играет ключевую роль в формировании плакоды [40]. Кроме того, показано, что в дентальной пластинке в местах, где будут развиваться моляры, увеличивается экспрессия BMP4, а там, где будут резцы — FGF8 [41].
Плакода является вторым сигнальным центром развития зуба. Место плакоды и будущего зуба определяется по увеличению экспрессии генов р21, р63, Msx2, Lef1 и Eda, а в мезенхиме — Lhx6,7,8, Barx1, Msx1,2, Dlx1,2, Pax9, Gli2 и молекул FGF, BMP и activin [4, 12, 23, 42].
Наиболее важными факторами формирования плакоды являются ген фактора транскрипции р63 и EDA (эктодисплазин) [17]. При нарушении функции р63 нарушается функционирование BMP-, FGF-, Notch- и EDA-зависимых сигнальных путей и плакода зубов не развивается [43]. EDA также относят к ключевым регуляторам развития плакод и эмалевых узлов зуба [17, 44]. Мишенями EDA-сигналинга являются важные сигнальные пути развития зуба в плакоде и в эмалевых узлах, такие как SHH-, FGF20-, DKK4-, CTGF- и Follistatin-зависимые пути. Мутации в гене Eda приводят к отсутствию третьих моляров или резцов и нарушению формы бугорков моляров [45]. Напротив, у мышей, у которых Eda экспрессируется в чрезмерных количествах, развиваются дополнительные зубы.
Плакода продуцирует молекулы, которые регулируют дальнейшее развитие зуба (см. рис. 4) [17]. Так, например, BMP, выделяемый плакодой, контролирует размер и количество резцов у мышей [46, 47].
Интересным оказалось, что эпителий на стадиях дентальной пластинки и плакоды обладает, а мезенхима не обладает одонтогенной компетентностью (см. рис. 4) [48, 49]. Однако эта одонтогенная компетентность эпителия после стадии плакоды у мышей теряется и переходит к мезенхиме [1, 50]. Поэтому на стадиях почки, колпачка и колокола уже не эпителий, а дентальная мезенхима способствует развитию зуба при комбинации даже с незубным эпителием [51, 52].
Перед стадией почки BMP из эпителиального сигнального центра плакоды индуцирует экспрессию BMP4 в мезенхиме. Увеличение экспрессии BMP4 в мезенхиме коррелирует со сдвигом одонтогенного потенциала от эпителия к дентальной мезенхиме [53] и индукцией в мезенхиме генов факторов транскрипции Msx1, Pax9 [53, 54], Lhx6,7, Barx1, Dlx1,2, Runx2, Gli2,3 [23, 55] (см. рис. 4). Показано, что активность этих, предположительно одонтогенных, факторов также регулируют FGF-зависимые пути [23]. Ингибирование хотя бы одного из этих факторов транскрипции приводит к остановке роста зуба на стадии плакоды или почки.
Во время перехода от стадии почки к стадии колпачка, эпителиальные FGF индуцируют в мезенхиме аctivin, FGF3 и FGF10, которые в свою очередь действуют реципрокно на эпителий [56, 57], стимулируя формирование эмалевых узлов на стадиях колпачка и колокола (см. рис. 4).
Первичный эмалевый узел — следующий, третий, эпителиальный сигнальный центр, реализует одонтогенный эффект мезенхимы. Первичный эмалевый узел выделяет WNT, FGF, BMP и SHH [35, 39, 58, 59]. FGF из эмалевого узла увеличивает в мезенхиме активность Runx2 и продукцию FGF3, которые в свою очередь влияют на эпителий, замыкая реципрокный сигналинг в механизме регуляции развития зуба [28].
На стадии колокола сигнальные молекулы первичного эмалевого узла стимулируют формирование вторичных эмалевых узлов, которые в свою очередь определяют расположение бугров на поверхности моляров [18, 60, 61] (см. рис. 3, 4). Происходит это благодаря тому, что клетки эмалевого узла выделяют FGF, который стимулирует окружающий эпителий к пролиферации и одновременно экспрессируют ингибитор клеточного цикла р21 и утрачивают рецепторы к FGF, и потому сами клетки эмалевого узла не делятся [62, 63].
Несмотря на многочисленные исследования, проблема понимания одонтогенной компетентности все же остается. Она состоит в том, что до сих пор так и не обнаружены одонтогенно-специфические молекулы, а все описанные выше гены, молекулы и факторы транскрипции участвуют не только в одонтогенезе, но и в развитии других тканей. При этом, однако, был установлен важный факт, что одонтогенной компетентностью обладают только клетки из ротовой полости [39, 59].
Естественные источники и сигнальные пути дифференцировки клеток зуба
Модулирование сигнальных путей, вовлеченных в эпителиально-мезенхимальные взаимодействия, например с помощью EDA, может привести к появлению дополнительных зубов [28, 44, 64], а модулирование FGF-, аctivin- и BMP-зависимых систем — к постоянному откладыванию эмали в резцах [65]. Это означает, что описанные выше сигнальные пути активно вовлечены в дифференцировку клеток, продуцирующих компоненты тканей зуба.
Дифференцировка клеток зуба начинается на стадии колокола (рис. 5). Одонтобласты дифференцируются из клеток зубного сосочка, цементобласты из клеток зубного фолликула, а амелобласты из эпителия. Эти клетки ответственны за образование и отложение дентина, цемента и эмали соответственно.
 Рис. 5. Источники и сигнальные пути дифференцировки клеток зуба. Автор И.Ю. Малышев.
Рис. 5. Источники и сигнальные пути дифференцировки клеток зуба. Автор И.Ю. Малышев.
Сигнальные молекулы из внутреннего эмалевого эпителия, такие как TGFb и BMP, начинают стимулировать клетки мезенхимы к дифференцировке в одонтобласты [66]. Нарушение TGF-β/BMP сигнального пути в зубном сосочке предупреждает дифференцировку одонтобластов и отложение дентина [67]. Сигнальные молекулы из эмалевых узлов, например WNT10b, также важны для дифференцировки одонтобластов [68]. Показано, что WNT10b регулирует экспрессию белков дентина [69, 70]. Базальная мембрана, выполняя функцию резервуара сигнальных молекул [66, 71], также вовлечена в дифференцировку одонтобластов.
Дифференцированные одонтобласты начинают посылать BMP2,4 и TGFb обратно к эпителию. В результате клетки эпителия дифференцируются в амелобласты [26, 72]. SHH из эпителиальных клеток дополнительно способствует дифференцировке и созреванию амелобластов [34, 35]. К факторам, регулирующим амелобласты, также относятся WNT3, EDA и Follistatin [23]. Дифференцированные амелобласты экспрессируют факторы транскрипции Sp6 и Msx2, которые участвуют в амелогенезе у мышей [23,24] благодаря активации амелобластспецифических генов ameloblastin, amelogenin, enamelin и Mmp-20 [23].
Очень мало известно о дифференцировке цементобластов. Предполагают, что они происходят из клеток дентального фолликула при контакте с дентином и действии BMP и WNT [73, 74].
Основные морфологические характеристики и клеточные маркеры амелобластов, одонтобластов и цементобластов представлены в таблице.
 Основные морфологические характеристики и маркеры амелобластов, одонтобластов и цементобластов
Основные морфологические характеристики и маркеры амелобластов, одонтобластов и цементобластов
Способность к регенерации и стволовые клетки в зубах млекопитающих
У людей постоянные зубы, которые однократно заменяют молочные, развиваются из дополнительной дентальной пластинки, которая формируется как часть пластинки молочного зуба [14, 75]. В отличие от людей рыбы и рептилии могут регенерировать новые зубы многократно. В механизм замены зуба вовлечены гены Pitx2 и Bmp4 [76], ген ингибитора ВМР/WNT, эктодина (Sostdc1) и ген Axin2, ингибитора обратной связи WNT-зависимого пути [75]. Смену зубов у позвоночных запускает увеличение активности WNT-зависимого пути [75, 77]. Увеличению активности WNT пути способствуют SHH- и BMP-сигнальные пути мезенхимы [77].
У мышей зубы в норме не замещаются. Однако с помощью активации WNT-пути можно стимулировать образование зубов de novo [59]. Другие млекопитающие, кроме мышей, не способны формировать зубы de novo. Это связано с тем, что, когда зуб прорезывается в полость рта, эмалевый эпителий, окружающий коронку зуба, исчезает, и единственный дентальный эпителий сохраняется в ERM. Хотя клетки ERM были выделены [78, 79] и дифференцированы в амелобластоподобные клетки [79], пока не ясно, имеют ли они одонтогенный потенциал.
Исследования показали, что стволовые клетки пульпы взрослого зуба (DPSC), молочных зубов (SHED) [80], апикального сосочка (SCAP) [50, 81—85] и зубного фолликула [1, 50] могут участвовать в регенерации твердых тканей зуба, а стволовые клетки периодонтальной связки (PDLSC) могут быть дифференцированы в одонтобласты, цементобласты и фибробласты и участвовать в продукции дентина и цемента, а также в регенерации периодонтальной связки [1]. Однако пока не доказано, что PDLSC сохранили одонтогенную компетентность.
Основные знания о дентальных стволовых клетках получены в исследованиях ниши стволовых клеток резцов мышей [86, 87]. Резцы грызунов, в отличие от других млекопитающих, растут постоянно и их эмаль расположена асимметрично (рис. 6). Губная сторона резца покрыта эмалью, а язычная только мягким дентином и цементом. Различие в твердости между губной и язычной стороной обеспечивает формирование режущего края. Асимметричность эмали определяется различием размеров ниш эпителиальных стволовых клеток, которые находятся в язычной и губной цервикальных петлях. Цервикальная петля образуется клетками звездчатого ретикулума, окруженными базальным эпителием. Стволовые клетки находятся в звездчатом ретикулуме. Предполагается, что стволовые клетки проникают в эпителий, там пролиферируют и, в зависимости от микроокружения, дифференцируются в амелобласты или корневой эпителий. Губная цервикальная петля значительно больше по сравнению с язычной, поэтому губная сторона резца покрыта эмалью, а язычная нет (см. рис. 6). В поддержании асимметрии ниши стволовых клеток резцов и ингибировании амелобластов важную роль играют гены Sprouty, которые кодируют ингибиторы FGF [88].
 Рис. 6. Цервикальная петля, ниша эпителиальных стволовых клеток постоянно растущего зуба. Стволовые клетки находятся в звездчатом ретикулуме, давая рост клеткам, которые проникают в базальный слой эпителия. В эпителиальном слое потомки стволовых клеток пролиферируют и мигрируют в сторону полости рта и там дифференцируются в амелобласты, которые откладывают эмалевый матрикс. Адаптирован с рисунка из I. Thesleff, M. Tummers (2008).
Рис. 6. Цервикальная петля, ниша эпителиальных стволовых клеток постоянно растущего зуба. Стволовые клетки находятся в звездчатом ретикулуме, давая рост клеткам, которые проникают в базальный слой эпителия. В эпителиальном слое потомки стволовых клеток пролиферируют и мигрируют в сторону полости рта и там дифференцируются в амелобласты, которые откладывают эмалевый матрикс. Адаптирован с рисунка из I. Thesleff, M. Tummers (2008).
Ниши стволовых клеток зуба, как и ниши других эктодермальных структур [89], окружены мезенхимальной тканью, которая продуцирует регуляторные молекулы. Эти молекулы, например FGF3 и FGF10 необходимы для самообновления, дифференцировки и выживаемости потомства эпителиальных стволовых клеток [86, 90, 91] (рис. 7). Ингибитор TGFβ фоллистатин ограничивает количество стволовых клеток и клеток-предшественников на язычной стороне, и благодаря этому ингибирует дифференцировку амелобластов, обеспечивая асимметрию эмали на резцах [91].
 Рис. 7. Различные фенотипы резцов, формирующиеся в результате модуляции сигнальных путей, регулирующих нишу стволовых клеток. Типичная асимметрия резцов грызунов нарушается, когда формирование эмали либо стимулируется, либо тормозится. Размер ниши стволовых клеток способствует чрезмерному росту резцов у Spry4–/–; Spry2+/– мышей и уменьшению размера резцов у Fgf3–/–; Fgf10+/– мышей. Адаптирован с рисунка из Thesleff I., Tummers M. (2008).
Рис. 7. Различные фенотипы резцов, формирующиеся в результате модуляции сигнальных путей, регулирующих нишу стволовых клеток. Типичная асимметрия резцов грызунов нарушается, когда формирование эмали либо стимулируется, либо тормозится. Размер ниши стволовых клеток способствует чрезмерному росту резцов у Spry4–/–; Spry2+/– мышей и уменьшению размера резцов у Fgf3–/–; Fgf10+/– мышей. Адаптирован с рисунка из Thesleff I., Tummers M. (2008).
В результате модуляции сигнальных путей, регулирующих нишу стволовых клеток, могут формироваться различные фенотипы резцов (см. рис. 7) [21, 26, 88, 91, 93]. Так, увеличение размера ниши стволовых клеток у Spry4–/–; Spry2+/– мышей способствует чрезмерному росту резцов, а снижение размера ниш у FGF-3–/– и FGF10+/– мышей — уменьшению размера резцов. Кроме того, снижение FGF10 приводит к прекращению развития корней у моляров мышей, а добавление FGF10 стимулирует сохранение цервикальных петель и ингибирует образование HERS [94].
Заключение
Изучение механизмов одонтогенеза безусловно имеет большое фундаментальное значение для понимания законов природы развития органов. Наряду с этим становится очевидным, что в хорошем понимании именно этих механизмов спрятан ключ к технологиям выращивания зуба и регенеративной стоматологии. Уже сейчас сформулированы несколько важных для биоинженерии зуба положений, вытекающих из того, что известно о молекулярных и клеточных основах одонтогенеза.
Во-первых, стало понятным, что наиболее успешный результат в выращивании зубов, его эквивалентов или компонентов зубной ткани может быть достигнут при использовании двух (а не одного) типов клеток, и мезенхимальных, и эпителиальных. Причем один из этих типов клеток должен обладать одонтогенным потенциалом. Этот подход уже взят на вооружение научными лабораториями и биотехнологическими компаниями. Он называется методом клеточно-тканевой рекомбинации. В основе метода лежит 3D-культивирование эпителиальных и мезенхимальных клеток в геле для воспроизведения начальных и последующих стадий одонтогенеза [7].
Во-вторых, понимание важности в механизмах естественного одонтогенеза экстраклеточного матрикса, разных факторов роста и разных клеток, а не только специализированных зубных (амелобласты, одонтобласты и цементобласты), привело к разработке тканеинженерного метода. Метод основан на использовании стволовых клеток из разных источников, iPS клеток, клеток, продуцирующих биологически активные вещества, например факторов роста сосудов, натуральных или искусственных матриксов с добавлением наиболее важных для развития зуба регуляторных и сигнальных молекул [7].
Разрабатывается еще несколько других направлений, имитирующих естественное развитие зуба. Нетрудно предсказать, что появление новых фактов и раскрытие новых закономерностей естественного одонтогенеза несомненно даст дополнительный мощный толчок новых технологий регенеративной стоматологии.
Благодарности: авторы благодарны к.м.н. Г.С. Руновой за помощь в редактировании текста и сделанные ценные замечания, а также М.А. Морозовой за техническую помощь в оформлении статьи.
Обзор написан при поддержке Министерства здравоохранения РФ (Государственное задание Министерства здравоохранения РФ № 056−00139−16 от 2 февраля 2016 г., уникальный номер реестровой записи 110 401 000 000 000 000 071 021 02).
Все авторы в равной степени принимали участие в подготовке материала.



