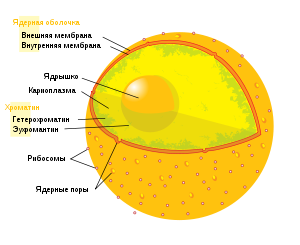
Ядерная оболочка

Ядро (лат. nucleus), органелла эукариотической клетки, содержащая генетическую информацию в форме молекул ДНК. В ядре происходит репликация — удвоение молекул ДНК, а также транскрипция — синтез молекул РНК на матрице ДНК. В ядре же синтезированные молекулы РНК подвергаются ряду модификаций, после чего выходят в цитоплазму. Сборка рибосом также происходит в ядре, в специальных образованиях, называемых ядрышками.
Содержание
Тонкая структура клеточного ядра
Хроматин
Огромная длина молекул ДНК эукариот предопределила появление специальных механизмов хранения, репликации и реализации генетического материала. Хроматином называют молекулы хромосомной ДНК в комплексе со специфическими белками, необходимыми для осуществления этих процессов. Основную массу составляют «белки хранения», так называемые гистоны. Из этих белков построены нуклеосомы, структуры на которые намотаны нити молекул ДНК. Нуклеосомы располагаются довольно регулярно, так что образующаяся структура напоминает бусы. Нуклеосома состоит из белков четырех типов: H2A, H2B, H3 и H4. В одну нуклеосому входят по два белка каждого типа — всего восемь белков. Гистон H1, более крупный чем другие гистоны, связывается с ДНК в месте ее входа на нуклеосому. Нуклеосома вместе с H1 называется хроматосомой.
Нить ДНК с нуклеосомами образует нерегулярную соленоид-подобную структуру толщиной около 30 нанометров, так называемую 30 нм фибриллу. Дальнейшая упаковка этой фибриллы может иметь различную плотность. Если хроматин упакован плотно, его называют конденсированным или гетерохроматином, он хорошо видим под микроскопом. ДНК находящаяся в гетерохроматине не транскрибируется, обычно это состояние характерно для незначащих или молчащих участков. В интерфазе гетерохроматин обычно располагается по периферии ядра (пристеночный гетерохроматин). Полная конденсация хромосом происходит перед делением клетки. Если хроматин упакован неплотно, его называют эу- или интерхроматином. Этот вид хроматина гораздо менее плотный при наблюдении под микроскопом и обычно характеризуется наличием транскрипционной активности. Плотность упаковки хроматина во многом определяется модификациями гистонов — ацетилированием и фосфориллированием.
Считается, что в ядре существуют так называемые функциональные домены хроматина(ДНК одного домена содержит приблизительно 30 тысяч пар оснований), то есть каждый участок хромосомы имеет собственную «территорию». К сожалению вопрос пространственного распределения хроматина в ядре изучен пока недостаточно. Известно, что теломерные (концевые) и центромерные (отвечающие за связывание сестринских хроматид в митозе) участки хромосом закреплены на белках ядерной ламины.
От цитоплазмы ядро отделено ядерной оболочкой, образованной за счёт расширения и слияния друг с другом цистерн эндоплазматической сети таким образом, что у ядра образовались двойные стенки за счёт окружающих его узких компартментов. Полость ядерной оболочки называется люменом или перинуклеарным пространством. Внутренняя поверхность ядерной оболочки подстилается ядерной ламиной, жёсткой белковой структурой, образованной белками-ламинами, к которой прикреплены нити хромосомной ДНК. Ламины прикрепляются к внутренней мембране ядерной оболочки при помощи заякоренных в ней трансмембранных белков — рецепторов ламинов. В некоторых местах внутренняя и внешняя мембраны ядерной оболочки сливаются и образуют так называемые ядерные поры, через которые происходит материальный обмен между ядром и цитоплазмой. Пора не является дыркой в ядре, а имеет сложную структуру, организованную несколькими десятками специализированных белков — нуклеопоринов. Под электронным микроскопом она видна как восемь связанных между собой белковых гранул с внешней и столько же с внутренней стороны ядерной оболочки.
![]()
Ядрышко
Ядрышко находится внутри ядра, и не имеет собственной мембранной оболочки, однако хорошо различимо под световым и электронным микроскопом. Основной функцией ядрышка является синтез рибосом. В геноме клетки имеются специльные участки, так называемые ядрышковые организаторы, содержащие гены рибосомной РНК (рРНК), вокруг которых и формируются ядрышки. В ядрышке происходит синтез рРНК РНК полимеразой I, ее созревание, сборка рибосомных субчастиц. В ядрышке локализуются белки, принимающие участие в этих процессах. Некоторые из этих белков имеют специальную последовательность — сигнал ядрышковой локализации (NoLS, от англ. Nucleolus Localization Signal). Следует отметить, самая высокая концентрация белка в клетке наблюдается именно в ядрышке. В этих структурах было локализовано около 600 видов различных белков, причем считается, что лишь небольшая их часть действительно необходима для осуществления ядрышковых функций, а остальные попадают туда неспецифически.
Под электронным микроскопом в ядрышке выделяют несколько субкомпартментов. Так называемые Фибриллярные центры окружены участками плотного фибриллярного компонента, где и происходит синтез рРНК. Снаружи от плотного фибриллярного компонента расположен гранулярный компонент, представляющий собой скопление созревающих рибосомных субчастиц.
Ядерный матрикс
Эволюционное значение клеточного ядра
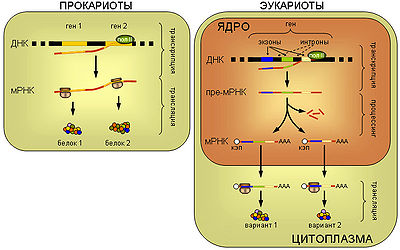
Основное функциональное отличие клеток эукариот от клеток прокариот заключается в пространственном разграничении процессов транскрипции (синтеза матричной РНК) и трансляции (синтеза белка рибосомой), что дает в распоряжение эукариотической клетки новые инструменты регуляции биосинтеза и контроля качества мРНК.
В то время, как у прокариот мРНК начинает транслироваться еще до завершения ее синтеза РНК-полимеразой, мРНК эукариот претерпевает значительные модификации (так называемый процессинг), после чего экспортируется через ядерные поры в цитоплазму, и только после этого может вступить в трансляцию. Процессинг мРНК включает несколько элементов.
Из предшественника мРНК (пре-мРНК) в ходе процесса, называемого сплайсингом вырезаются интроны — незначащие участки, а значащие участки — экзоны соединяются друг с другом. Причем экзоны одной и той же пре-мРНК могут быть соединены несколькими разными способами (альтернативный сплайсинг), так что один предшественник может превращаться в зрелые мРНК нескольких разных видов. Таким образом, один ген может кодировать сразу несколько белков.
Кроме того, интрон-экзонная структура генома, практически невозможная у прокариот (т.к. рибосомы смогут транслировать незрелые мРНК), дает эукариотам определенную эволюционную мобильность. Учитывая протяженность интронных участков, рекомбинация между двумя генами зачастую сводится к обмену экзонами. Благодаря тому, что экзоны часто соответствуют функциональным доменам белка, участки получившегося в результате рекомбинации «гибрида», зачастую сохраняют свои функции. В то же время у прокариот рекомбинация между генами невозможна без разрыва в значащей части, что безусловно уменьшает шансы на то, что получившийся белок будет функционален.
Строение ядерной оболочки
Строение ядерной оболочки

Клеточное ЯДРО
1. Общая характеристика интерфазного ядра|ядра. Функции ядра|ядра
2. Ядерная оболочка, строение и функциональное значение
3. Строение и функции хроматина и хромосом
1. Общая характеристика интерфазного ядра|ядра
Ядро — это важнейшая составная часть клетки, которая имеется практически во всех клетках многоклеточных организмов. Большинство клеток имеет одно ядро, но бывают двуядерные и многоядерные клетки (например, поперечно-полосатые мышечные волокна|волокна). Двуядерность и многоядерность обусловлены функциональными особенностями или патологическим состоянием клеток. Форма и размеры ядра|ядра очень изменчивы и зависят от вида организма, типа, возраста и функционального состояния клетки. В среднем объем|объём ядра|ядра составляет приблизительно 10% от общего объёма клетки. Чаще всего ядро имеет округлую или овальную форму размером от 3 до 10 мкм в диаметре. Минимальный размер ядра|ядра составляет 1 мкм (у некоторых простейших), максимальный — 1 мм (яйцеклетки некоторых рыб и земноводных). В некоторых случаях наблюдается зависимость формы ядра|ядра от формы клетки. Ядро обычно занимает центральное положение, но в дифференцированных клетках может быть смещено к периферийному участку клетки. В ядре сосредоточена практически вся ДНК эукариотической клетки.
Основными функциями ядра|ядра являются:
1) Хранение и передача генетической информации;
2) Регуляция синтеза белка|белка, обмена веществ и энергии в клетке.
Таким образом, ядро является не только вместилищем генетического материала, но и местом, где этот материал функционирует и воспроизводится. Поэтому нарушение любой|любой из этих функций приведёт к гибели клетки. Всё|Все это указывает на ведущее значение ядерных структур в процессах синтеза нуклеиновых кислот и белков.
Одним из первых учёных продемонстрировавших роль ядра|ядра в жизнедеятельности клетки был немецкий биолог Хаммерлинг. В качестве экспериментального объекта Хаммерлинг использовал крупные одноклеточные морские водоросли Acetobularia mediterranea и А. crenulata. Эти близкородственные виды хорошо отличаются друг от друга по форме «шляпки». В основании стебелька находится ядро. В одних экспериментах шляпку отделяли от нижней части стебелька. В результате было установлено, что для нормального развития шляпки необходимо ядро. В других экспериментах стебелёк с ядром одного вида водоросли соединялся со стебельком без ядра|ядра другого вида. У образовавшихся химер всегда развивалась шляпка, типичная для того вида, которому принадлежало ядро.
Общий план строения интерфазного ядра|ядра одинаков у всех клеток. Ядро состоит из ядерной оболочки, хроматина, ядрышек, ядерного белкового матрикса и кариоплазмы (нуклеоплазмы). Эти компоненты встречаются практически во всех неделящихся клетках эукариотических одно- и многоклеточных организмов.
2. Ядерная оболочка, строение и функциональное значение
Ядерно-цитоплазматический транспорт
Процесс транслокации субстрата через ядерную пору (для случая|случая импорта) состоит из нескольких стадий. На первой стадии транспортирующийся комплекс заякоривается на обращённой в цитоплазму фибрилле. Затем фибрилла сгибается и перемещает комплекс ко входу в канал ядерной поры|поры. Происходит собственно транслокация и освобождение комплекса в кариоплазму. Известен и обратный процесс — перенос веществ из ядра|ядра в цитоплазму. Это в первую очередь касается транспорта РНК синтезируемого исключительно в ядре. Также существует другой путь переноса веществ из ядра|ядра в цитоплазму. Он связан с образованием выростов ядерной оболочки, которые могут отделяться от ядра|ядра в виде вакуолей, а затем содержимое их изливается или выбрасывается в цитоплазму.
Таким образом, обмен веществ между ядром и цитоплазмой осуществляется двумя основными путями: через поры|поры и путём отшнуровывания.
Функции ядерной оболочки:
1. Барьерная. Эта функция заключается в отделении содержимого ядра|ядра от цитоплазмы. В результате оказываются пространственно разобщёнными процессы синтеза РНК/ДНК от синтеза белка|белка.
3. Организующая. Одной из основных функций ядерной оболочки является её участие в создании внутриядерного порядка.
3. Строение и функции хроматина и хромосом
Наследственный материал может находиться в ядре клетки в двух структурно-функциональных состояниях:
1. Хроматин. Это деконденсированное, метаболически активное состояние, предназначенное для обеспечения процессов транскрипции и редупликации в интерфазе.
2. Хромосомы. Это максимально конденсированное, компактное, метаболически неактивное состояние, предназначенное для распределения и транспортировки генетического материала в дочерние клетки.
Хроматин. В ядре клеток выявляются зоны плотного вещества, которые хорошо окрашиваются основными красителями. Эти структуры получили название «хроматин» (от греч. «хромо» – цвет, краска). Хроматин интерфазных ядер представляет собой хромосомы, находящиеся в деконденсированном состоянии. Степень деконденсации хромосом может быть различной. Зоны полной деконденсации называются эухроматином. При неполной деконденсации в интерфазном ядре видны участки конденсированного хроматина, называемого гетерохроматином. Степень деконденсации хроматина в интерфазе отражает функциональную нагрузку этой структуры. Чем «диффузнее» распределён хроматин в интерфазном ядре, тем интенсивнее в нём синтетические процессы. Уменьшение синтеза РНК в клетках обычно сопровождается увеличением зон конденсированного хроматина. Максимальная конденсация конденсированного хроматина достигается во время митотического деления клеток. В этот период хромосомы не выполняют никаких синтетических функций.
В химическом отношении хроматин состоит из ДНК (30-45%), гистонов (30-50%), негистонных белков (4-33%) и небольшого количества РНК. ДНК эукариотических хромосом представляет собой линейные молекулы, состоящие из тандемно (друг за другом|другом) расположенных репликонов разного размера. Средний размер репликона около 30 мкм. Репликоны — участки ДНК, которые синтезируются как независимые единицы. Репликоны имеют начальную и терминальную точки синтеза ДНК. РНК представляет собой всё|все известные клеточные типы РНК, находящиеся в процессе синтеза или созревания. Гистоны синтезируются на полисомах в цитоплазме, причём этот синтез начинается несколько раньше редупликации ДНК. Синтезированные гистоны мигрируют из цитоплазмы в ядро, где и связываются с участками ДНК.
В структурном отношении хроматин представляет собой нитчатые комплексные|комплексные молекулы дезоксирибонуклеопротеида (ДНП), которые состоят из ДНК, ассоциированной с гистонами. Хроматиновая нить представляет собой двойную спираль ДНК, окружающую гистоновый стержень. Она состоит из повторяющихся единиц – нуклеосом. Количество нуклеосом огромно.
Хромосомы (от. греч. хромо и сома|сома) — это органоиды клеточного ядра|ядра, являющиеся носителями генов и определяющие наследственные свойства клеток и организмов.
Хромосомы представляют собой палочковидные структуры разной длины|длины с довольно постоянной толщиной. У них имеется зона первичной перетяжки, которая делит хромосому на два плеча. Хромосомы с равными называют метацентрическими, с плечами неодинаковой длины|длины — субметацентрическими. Хромосомы с очень коротким, почти незаметным вторым плечом называются акроцентрическими.
В области первичной перетяжки находится центромера, представляющая собой пластинчатую структуру в виде диска. К центромере прикрепляются пучки микротрубочек митотического веретена|веретёна, идущие в направлении к центриолям. Эти пучки микротрубочек принимают участие в движении хромосом к полюсам|полюсам клетки при митозе. Некоторые хромосомы имеют вторичную перетяжку. Последняя обычно расположена вблизи дистального конца хромосомы и отделяет маленький участок, спутник. Вторичные перетяжки называют ядрышковыми организаторами. Здесь локализована ДНК, ответственная за синтез р-РНК. Плечи хромосом оканчиваются теломерами, конечными участками. Теломерные концы хромосом не способны соединяться с другими хромосомами или их фрагментами. В отличие от них разорванные концы хромосом могут присоединяться к таким же разорванным концам других хромосом.
Размеры хромосом у разных организмов варьируют в широких пределах. Так, длина хромосом может колебаться от 0,2 до 50 мкм. Самые мелкие хромосомы обнаруживаются у некоторых простейших, грибов. Наиболее длинные — у некоторых прямокрылых насекомых, у амфибий и у лилейных. Длина хромосом человека находится в пределах 1,5-10 мкм.
Число хромосом у различных объектов также значительно колеблется, но характерно для каждого вида животных или растений. У некоторых радиолярий число хромосом достигает 1000-1600. Рекордсменом среди растений по числу хромосом (около 500) является папоротник ужовник, 308 хромосом у тутового дерева. Наименьшее количество хромосом (2 на диплоидный набор) наблюдается у малярийного плазмодия, лошадиной аскариды. У человека число хромосом составляет 46, у шимпанзе, таракана и перца – 48, плодовая мушка дрозофила – 8, домашняя муха – 12, сазана – 104, ели и сосны|сосны – 24, голубя|голубя — 80.
Кариотип (от греч. Карион — ядро, ядро ореха, операторы — образец, форма) — совокупность признаков хромосомного набора (число, размер, форма хромосом), характерные|характерные для того или иного вида.
Особи разного пола (особенно у животных) одного и того же вида могут различаться по числу хромосом (различие чаще всего на одну хромосому). Даже у близких видов хромосомные наборы отличаются друг от друга или по числу хромосом, или по величине хотя бы одной или нескольких хромосом. Следовательно, структура кариотипа может быть таксономическим признаком.
Во второй половине 20 века|века в практику хромосомного анализа стали внедряться методы дифференциального окрашивания хромосом. Считается, что способность отдельных участков хромосом к окрашиванию связана с их химическими различиями.
Ядрышко (нуклеола) — обязательный компонент клеточного ядра|ядра эукариотных организмов. Однако имеются некоторые исключения. Так ядрышки отсутствуют в высокоспециализированных клетках, в частности в некоторых клетках крови. Ядрышко представляет собой плотное тельце|тельце округлой формы величиной 1-5 мкм. В отличие от цитоплазматических органоидов ядрышко не имеет мембраны, которая окружала бы его содержимое. Размер ядрышка отражает степень его функциональной активности, которая широко варьирует в различных клетках. Ядрышко является производным хромосомы. В состав ядрышка входят белок|белок, РНК и ДНК. Концентрация РНК в ядрышках всегда выше концентрации РНК в других компонентах клетки. Так концентрация РНК в ядрышке может быть в 2-8 раз выше, чем в ядре, и в 1-3 раза выше, чем в цитоплазме. Благодаря высокому содержанию РНК, ядрышки хорошо окрашиваются основными красителями. ДНК в ядрышке образует большие|большие петли|петли, которые носят название «ядрышковые организаторы». От них зависит образование и количество ядрышек в клетках. Ядрышко неоднородно по своему строению. В нём выявляются два основных компонента: гранулярный и фибриллярный. Диаметр гранул около 15-20 нм, толщина фибрилл – 6-8 нм. Фибриллярный компонент может быть сосредоточен в центральной части ядрышка, а гранулярный — по периферии. Часто гранулярный компонент образует нитчатые структуры — нуклеолонемы толщиной около 0, 2 мкм. Фибриллярный компонент ядрышек представляет собой рибонуклеопротеидные тяжи предшественников рибосом, а гранулы — созревающие субъединицы рибосом. Функция ядрышка заключается в образовании рибосомных РНК (рРНК) и рибосом, на которых происходит синтез полипептидных цепей в цитоплазме. Механизм о
разования рибосом следующий: на ДНК ядрышкового организатора образуется предшественник рРНК, который в зоне ядрышка одевается белком. В зоне ядрышка происходит сборка субъединиц рибосом. В активно функционирующих ядрышках синтезируется 1500-3000 рибосом в минуту. Рибосомы из ядрышка через поры|поры в ядерной оболочке поступают на мембраны эндоплазматической сети. Количество и образование ядрышек связано с активностью ядрышковых организаторов. Изменения числа|числа ядрышек могут происходить за счёт слияния ядрышек или при сдвигах в хромосомном балансе клетки. Обычно в ядрах содержится несколько ядрышек. В ядрах некоторых клеток (ооциты тритонов) содержится большое количество ядрышек. Это явление получило название амплификации. Оно заключается в организации систем управления качеством, что происходит сверхрепликация зоны ядрышкового организатора, многочисленные копии отходят от хромосом и становятся дополнительно работающими ядрышками. Такой процесс необходим для накопления огромного количества рибосом на яйцеклетку. Благодаря этому обеспечивается развитие эмбриона на ранних стадиях даже при отсутствии синтеза новых рибосом. Сверхчисленные ядрышки после созревания яйцевой клетки исчезают.
Судьба ядрышка при делении клеток. По мере затухания синтеза р-РНК в профазе происходит разрыхление ядрышка и выход готовых рибосом в кариоплазму, а затем и в цитоплазму. При конденсации хромосом фибриллярный компонент ядрышка и часть гранул тесно ассоциируют с их поверхностью, образуя основу матрикса митотических хромосом. Этот фибриллярно-гранулярный материал переносится хромосомами в дочерние клетки. В ранней телофазе по мере деконденсации хромосом происходит высвобождение компонентов матрикса. Его фибриллярная часть начинает собираться в мелкие многочисленные ассоциаты — предъядрышки, которые могут объединяться друг с другом. По мере возобновления синтеза РНК предъядрышки превращаются в нормально функционирующие ядрышки.
Ядерный белковый матрикс (ядерный скелет) — каркасная внутриядерная система, которая служит для поддержания общей структуры интерфазного ядра|ядра объединения всех ядерных компонентов. Представляет собой нерастворимый материал, остающийся в ядре после биохимических экстракций. Он не имеет чёткой морфологической структуры и состоит на 98% из белков.
Строение ядерной оболочки

Ядерная оболочка строение,функции,роль ядра|ядра при взаимодействии с цитоплазмой.
Роль ядра|ядра:Ядро осуществляет две группы общих функций: одну, связанную собственно с хранением генетической информации, другую — с её реализацией, с обеспечением синтеза белка|белка.
В первую группу входят процессы, связанные с поддержанием наследственной информации в виде неизменной структуры ДНК. Эти процессы связаны с наличием так называемых репарационных ферментов, ликвидирующих спонтанные повреждения молекулы ДНК (разрыв одной из цепей ДНК, часть радиационных повреждений), что сохраняет строение молекул ДНК практически неизменным в ряду поколений клеток или организмов. Далее, в ядре происходит воспроизведение или редупликация молекул ДНК, что даёт возможность двум клеткам получить совершенно одинаковые и в качественном и в количественном смысле объёмы генетической информации. В ядрах происходят процессы изменения и рекомбинации генетического материала, что наблюдается во время мейоза (кроссинговер). Наконец, ядра|ядра непосредственно участвуют в процессах распределения молекул ДНК при делении клеток.
Другой группой клеточных процессов, обеспечивающихся активностью ядра|ядра, является создание собственно аппарата белкового синтеза. Это не только синтез, транскрипция на молекулах ДНК разных информационных РНК и рибосомных РНК. В ядре эукариотов происходит также образование субъедениц рибосом путём комплексирования синтезированных в ядрышке рибосомных РНК с рибосомными белками|белками, которые синтезируются в цитоплазме и переносятся в ядро.
Таким образом, ядро представляет собой не только вместилище генетического материала, но и место, где этот материал функционирует и воспроизводится. Поэтому выпадение лил нарушение любой|любой из перечисленных выше функций губительно для клетки в целом. Так нарушение репарационных процессов будет приводить к изменению первичной структуры ДНК и автоматически к изменению структуры белков, что непременно скажется на их специфической активности, которая может просто исчезнуть или измениться так, что не будет обеспечивать клеточные функции, в результате чего клетка погибает. Нарушения редупликации ДНК приведут к остановке размножения клеток или к появлению клеток с неполноценным набором генетической информации, что также губительно для клеток. К такому же результату приведёт нарушение процессов распределения генетического материала (молекул ДНК) при делении клеток. Выпадение в результате поражения ядра|ядра или в случае нарушений каких-либо регуляторных процессов синтеза любой|любой формы РНК автоматически приведёт к остановке синтеза белка|белка в клетке или к грубым его нарушениям.
Хаммерлинг показал, что для нормального развития шляпки необходимо ядро. В дальнейших экспериментах, в которых соединяли нижнюю часть, содержащую ядро одного вида с лишённым ядра|ядра стебельком другого вида, у таких химер всегда развивалась шляпка, типичная для того вида, которому принадлежит ядро.
Сам термин ядро впервые был применён Броуном в 1833 г. Для обозначения шаровидных постоянных структур в клетках растений. Позднее|Позднее такую же структуру описали во всех клетках высших организмов.
Ядра|Ядра имеют обычно шаровидную или яйцевидную форму; диаметр первых равен приблизительно 10 мкм, а длина вторых — 20 мкм.
Ядро необходимо для жизни клетки, поскольку именно оно регулирует всю её активность. Связано это с тем, что ядро несёт в себе генетическую (наследственную) информацию, заключённую в ДНК.
Ядерная оболочка
Мембраны ядерной оболочки в морфологическом отношении не отличаются от остальных внутриклеточных мембран: они имеют толщину около 7 нм и состоят из двух осмиофильных слоёв.
Строение ядерной оболочки
Внешняя мембрана ядерной оболочки, непосредственно контактирующая с цитоплазмой клетки, имеет ряд сруктурных особенностей, позволяющих отнести её к собственно мембранной системе эндоплазматического ретикулума. Так, на внешней ядерной мембране обычно располагается большое количество рибосом. У большинства животных и растительных клеток внешняя мембрана ядерной оболочки не представляет собой идеально ровную поверхность — она может образовывать различной величины|величины выпячивания или выросты в сторону цитоплазмы.
Внутренняя мембрана контактирует с хромосомным материалом ядра|ядра (см. Ниже).
Число ядерных пор зависит от метаболической активности клеток: чем выше синтетические процессы в клетках, тем больше пор на единицу поверхности клеточного ядра|ядра.
Химия ядерной оболочки
В составе ядерных оболочек обнаруживаются небольшие количества ДНК (0-8%), РНК (3-9%), но основными химическими компонентами являются липиды (13-35%) и белки|белки (50-75%), что для всех клеточных мембран.
Состав липидов сходен с таковым в мембранах микросом или мембранах эндоплазматической сети. Ядерные оболочки характеризуются относительно низким содержанием холестерина и высоким — фосфолипидов, обогащённых насыщенными жирными кислотами.
Белковый состав мембранных фракций очень сложен. Среди белков обнаружен ряд ферментов, общих с ЭР (например, глюкозо-6-фосфатаза, Mg-зависимая АТФаза, глютамат-дегидрогеназа и др.) не обнаружена РНК-полимераза. Тут выявлены активности многих окислительных ферментов (цитохромоксидазы, НАДН-цитохром-с-редуктазы) и различных цитохромов.
Среди белковых фракций ядерных мембран встречаются основные белки|белки типа гистонов, что объясняется связью участков хроматина с ядерной оболочкой.
Ядерная оболочка и ядерно-цитоплазматический обмен
Ядерная оболочка — система, разграничивающая два основных клеточных отсека: цитоплазму и ядро. Ядерные оболочки полностью проницаемы для ионов, для веществ малого молекулярного веса|веса, таких, как сахара|сахара|сахара, аминокислоты|аминокислоты, нуклеотиды. Считается, что белки|белки молекулярного веса|веса до 70 тыс. И размером не больше 4,5 нм могут свободно диффундировать через оболочку.
Известен и обратный процесс — перенос веществ из ядра|ядра в цитоплазму. Это в первую очередь касается транспорта РНК синтезируещегося исключительно в ядре.
Ещё один путь транспорта веществ из ядра|ядра в цитоплазму связан с образованием выростов ядерной оболочки, которые могут отделяться от ядра|ядра в виде вакуолей, содержимое их затем изливается или выбрасывается в цитоплазму.
Таким образом, из многочисленных свойств и функциональных нагрузок ядерной оболочки следует подчеркнуть её роль как барьера, отделяющего содержимое ядра|ядра от цитоплазмы, ограничивающего свободный доступ в ядро крупных агрегатов биополимеров, барьера, активно регулирующего транспорт макромолекул между ядром и цитоплазмой.
Одной из основных функций ядерной оболочки следует считать также её участие в создании внутриядерного порядка, в фиксации хромосомного материала в трёхмерном пространстве ядра|ядра.
Ядерный матрикс
Этот комплекс не представляет собой какую-то чистую фракцию, сюда входят компоненты и ядерной оболочки, и ядрышка, и кариоплазмы. С ядерным матриксом оказались связаны как гетерогенная РНК, так и часть ДНК. Эти наблюдения дали основание считать, что матрикс ядра|ядра играет важную роль не только в поддержании общей структуры интерфазного ядра|ядра, но и может участвовать в регуляции синтеза нуклеиновых кислот.
Видео по теме : Строение ядерной оболочки



