Что такое пептидогликан в микробиологии
• Большинство бактерий содержат пептидогликан, жесткую внешнюю клеточную стенку, образованную полимерной сеткой гликановых нитей, связанных между собой короткими пептидами
• Пентапептиды дисахаридной природы, являющиеся предшественниками пептидогликана, образуются в цитоплазме, экспортируются из клетки, и собираются вне цитоплазматической мембраны
• Согласно одной из моделей, объясняющей синтез клеточной стенки, мульти белковый комплекс вставляет новый материал, следуя принципу: «прежде чем удалить — сделай»
• Перестройка, модификация и репарация клеточной стенки происходят с участием многих автолитических ферментов
• У некоторых бактерий пептидогликан клеточной стенки играет важную роль в поддержании формы клетки
• Гомолог бактериального актина, MreВ, образует в цитоплазме спиральные филаменты, которые определяют форму клетки, контролируя синтез пептидогликана
У большинства бактерий основной компонент клеточной оболочки называется пептидогликан. Он состоит из сетки гликановых нитей, связанных пептидами. Полимер покрывает всю поверхность микроорганизма, образуя жесткую защитную оболочку. Клеточная стенка, состоящая из пептидогликана, играет критическую роль в обеспечении выживаемости бактерий, поскольку она оказывает сопротивление силе, возникающей за счет высокой внутренней осмолярности клетки, которая действует на цитоплазматическую мембрану.
Поэтому разрыв стенки имеет для клетки катастрофические последствия. Пептидогликан также играет роль в поддержании формы большинства клеток бактерий.
Известные виды бактерий подразеляются на две группы, которые отличаются друг от друга по отношению к прокрашиванию. Различное отношение грамположительных и грамотрицательных бактерий к прокрашиванию по Граму отражает фундаментальные различия в организации их клеточной стенки. У грамотрицательных бактерий присутствует тонкий слой пептидогликана, вероятно, состоящий из одного слоя гликановых нитей.
Поэтому он обладает двухмерной структурой и связующие пептидные мостики располагаются в той же плоскости, что и гликановые цепи. Напротив, у грамположительных бактерий слой пептидогликана гораздо толще и включает несколько слоев гликановых нитей. В этом случае поперечные мостики, вероятно, лежат в разных плоскостях и соединяют нити, находящиеся в одной плоскости с нитями расположенного ниже слоя. Неизвестно, почему у грамположительных и грамотрицательных бактерий сборка пептидогликана происходит столь разными путями. Одно из возможных преимуществ более толстой клеточной стенки заключается в том, что она обладает большей прочностью и поэтому может обеспечивать большую степень защиты от физических воздействий, а также от высокого внутреннего осмотического давления, характерного для грамположительных клеток.
Как показано на рисунке ниже, гликановые нити обычно состоят из повторяющегося дисахарида, N-ацетилглюкозамин-N-ацетилмурамовая кислота (NAG-NAM). Средний размер нитей составляет порядка 30 дисахаридов, однако он варьирует в широких пределах. К NAM ковалентно присоедияется пептид, который синтезируется вне рибосом и включает несколько небычных аминокислот, D-глутаминовую кислоту, D-аланин и диаминопимелиновую кислоту (БАР). (Аминокислоты, используемые для синтеза белка, обязательно должны находиться в L-форме.) DAP служит источником аминогрупп, которые участвуют в образовании сшивок между гликановыми нитями.
Сшивки приводят к высвобождению последнего D-аланина из пептидной цепи. У некоторых бактерий в состав пептидных мостиков входят дополнительные аминокислоты. У грамположительных бактерий нити NAM-NAG. могут соединяться одним или несколькими остатками глицина в пептидных мостиках.
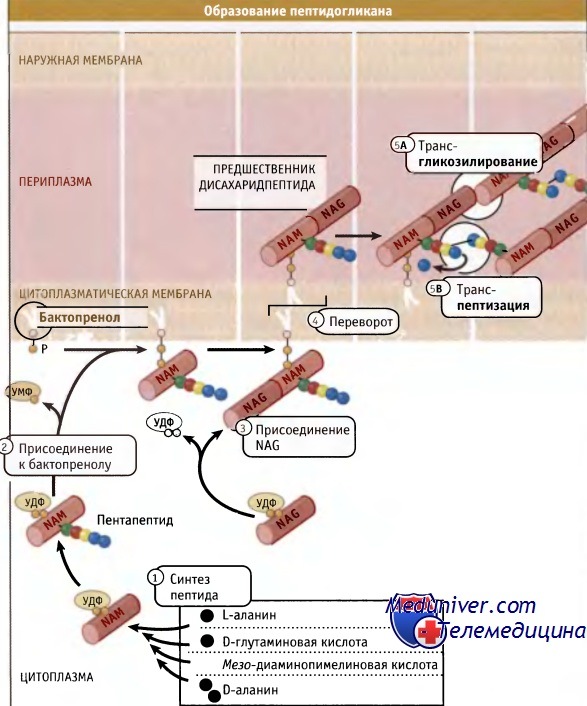 Синтез пептидогликана поисходит в несколько этапов, начинаясь в цитоплазме и заканчиваясь на наружной стороне цитоплазматической мембраны.
Синтез пептидогликана поисходит в несколько этапов, начинаясь в цитоплазме и заканчиваясь на наружной стороне цитоплазматической мембраны.
В ходе реакции трансгликозилирования из NAM высвобождается бактопренол (на рисунке не показан).
Как показано на рисунке ниже, дисахаридные предшественники образуются в цитозоле, начиная с уридин-5-дифосфата (УДФ)-NAM. Цепь аминокислот формируется последовательно, и в каждм шаге участвует один фермент. Образующийся пентапептид УДФ-NAM присоединяется к специфическому липиду клеточной мембраны (бактопренолу). Добавление NAG завершает построение дисахаридного предшественника, который затем ориентируется по направлению к внешней стороне мембраны для дальнейшей сборки существующего пептидогликана.
Для включения дисахаридного предшественника в полимер необходимо протекание двух ферментативных реакций: трансгликозилирования для формирования гликановых цепей и транспептидации для образования сшивок. Ферменты, ответственные за транспептидацию, были обнаружены при изучении связывания пенициллина, токсический эффект которого на клетки бактерий связан с его ингибированием транспептидаз. Поэтому они были названы пенициллин-связывающими белками (РВР) Высокомолекулярные белки, относящиеся к классу А РВР, представляют собой бифункциональные ферменты, которые характеризуются наличием отдельного дополнительного трансгликозилазного домена, а также домена, обладающего транспептидазной активностью. У палочковидных бактерий цилиндрическая часть клетки и полукруглые торцы (образовавшиеся при делении), вероятно, требуют специальных машин синтеза пептидогликана.
У Е. coli эти особые транспептидазы представлены белками РВР2 и РВР3 соответственно. Существуют данные, свидетельствующие о том, что эти транспептидазы используют несколько разные субстраты. Так, первая использует боковую цепь пентапептида, а вторая — тетра- или трипептид. Различий в структуре протеогликанов, синтезированных с участием двух этих ферментов, не обнаружено.
Для того чтобы клетка могла расти и делиться, ковалентные связи в пептидогликане должны быть разорваны с тем, чтобы обеспечить включение новых его участков. Существует привлекательная точка зрения, которая объясняет, каким образом обеспечивается безопасное включение новых единиц пептидогликана. Она основана на принципе, которвш можно сформулировать так: «прежде чем удалить — сделай». Иными словами, новые нити добавляются в места образования новых участков до разрыва существующих связей. Как показано на рисунке ниже, процесс в целом контролируется мультиферментным комплексом, который содержит РВР, необходимые для добавления новых участков, и ферменты автолиза, которые затем гидролизуют существующие.
Согласно модели «3 вместо 1», комплекс, содержащий две бифункциональные транспептидазы-трансгликозилазы и специальную гипотетическую трансгликозилазу, образует «тройную упаковку» новых сшитых между собой нитей. Две монофункциональные транспептидазы комплекса связывают новые внешние нити с существующими.
Связывание происходит с любой стороны «матричной» нити. Затем матричная нить гидролизуется ферментом автолиза, который также входит в состав комплекса. Хотя появляются указания на существование ассоциатов, содержащих перечисленные ферменты, остается убедиться в том. что все остальные детали предложенной схемы соответствуют действительной картине. Одна из трудностей, возникающая при попытке проверить эту модель, заключается в том, что для многих бактерий характерна высокая избыточность набора РВР, так что выключение соответствующего гена практически не отражается на фенотипе.
 Изолированная саккула клеточной стенки интактна, но принимает двухмерную структуру.
Изолированная саккула клеточной стенки интактна, но принимает двухмерную структуру.
Саккула была получена с использованием агаровой фильтрации.
Однако у большинства бактерий в клеточной стенке содержится множество ферментов автолиза. Эти ферменты действуют на разные связи в пептидогликане. По большей части функции автолизинов неизвестны, за исключением их очевидной роли в деградации клеточной стенки, позволяющей разойтись дочерним клеткам, образовавшимся в конце акта деления. Так же как и в случае РВР, нокаут множественных генов, кодирующих аутолизин, обычно практически не влияет на фенотип, за исключением некоторого утолщения клеточной стенки и задержки разделения клеток.
Существует несколько групп данных, в пользу точки зрения о том, что пептидогликан клеточной стенки играет ключевую роль в поддержании формы бактериальной клетки. Во-первых, как иллюстрирует рисунок ниже, изолированная саккула сохраняет форму клетки, из которой она была получена. Во-вторых, обработка клетки таким ферментом, как лизоцим, который гидролизует стенку, приводит к утрате клеткой своей формы. В-третьих, мутации, приводящие к изменению клеточной формы, часто затрагивают гены, принимающие участие в синтезе клеточной стенки.
На протяжении десятилетий считалось, что бактерии не обладают актиновым цитоскелетом, который определяет форму эукариотических клеток. Однако недавно оказалось, что белок бактерий, MreВ, характеризующийся очень низкой степенью структурной гомологии с актином, фактически является его функциональным гомологом. Действительно, при рентгеноструктурном анализе этого белка обнаружена трехмерная складка, которая совпадает с характерной для актина. Гены mreВ впервые были идентифицированы при исследовании мутантов Е. coli и В. subtilis, у которых отмечалось нарушение клеточных форм. На рисунке ниже показано влияние мутаций в двух отдельных гомологичных генах — mreВ и mbl (близкий к mreВ) на форму клеток В. subtilis.
Ген mreВ найден почти во всех бактериях несферической формы, однако отсутствует у кокков (обладающих круглой формой). Если в качестве исходной принять простую сферическую симметрию круглой формы, то белок MreВ может участвовать в функционировании клеточной системы, обеспечивающей образование более сложных форм: палочковидных (Е. coli и В. subtilis), изогнутых (вибрион) и спирализованных (Helicobacter). На рисунке ниже представлены клетки В. subtilis, экспрессирующие конструкцию, представляющую собой конъюгат кодирующей области белка Mbl и зеленого флуоресцирующего белка (GFP). Белки образуют спирализованные филаментарные структуры непосредственно под клеточной мембраной. Сами по себе эти структуры не обладают достаточной жесткостью, чтобы непосредственно определять форму клетки.
Если искусственно нарушить цилиндрическую форму клетки, например удалив клеточную стенку и приготовив протопласты, Mbl-филаменты утрачивают спиральную конфигурацию. Вероятно, они функционируют, регулируя синтез материала для новой клеточной стенки таким образом, что во время роста, в целом, поддерживаются форма клетки и ее размеры.
Пока окончательно не выяснено, как MreB-филаменты взаимодействуют с клеточной стенкой. Однако гены mreB почти всегда располагаются выше двух других консервативных генов бактерий, mreC и mreD. Оба этих гена также определяют форму клетки и кодируют трансмембранные белки, которые, в принципе, могут сопрягать информацию, получаемую от спирализованных MreB-филаментов внутри клетки, с синтезом клеточной стенки, происходящим вне ее.
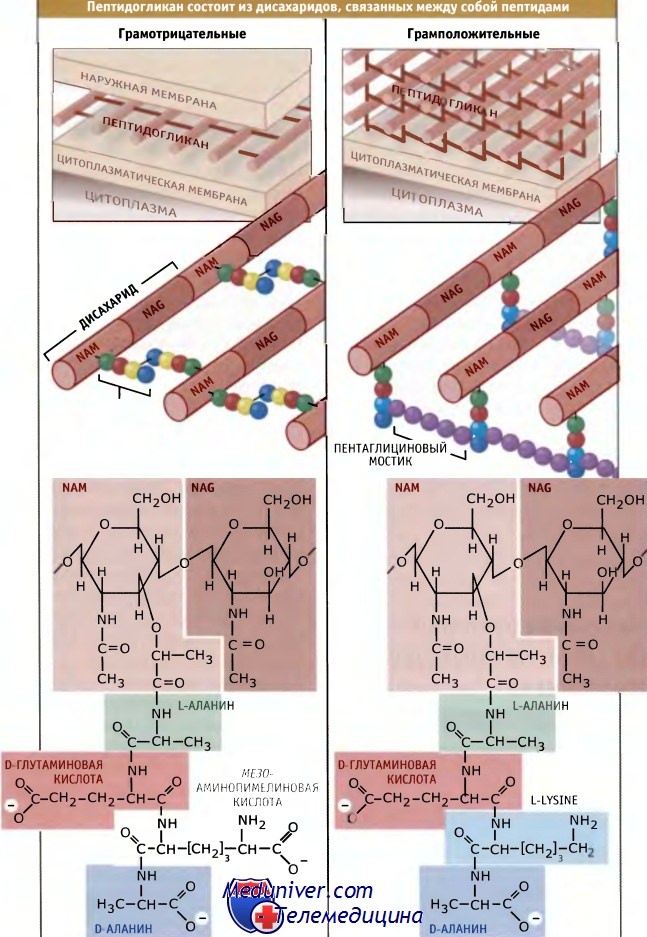 Слой пептидогликана у грамотрицательных и грамположительных бактерий содержит одни и те же повторяющиеся дисахариды, однако различается по пептидным связкам.
Слой пептидогликана у грамотрицательных и грамположительных бактерий содержит одни и те же повторяющиеся дисахариды, однако различается по пептидным связкам.
Некоторые грамположительные бактерии вместо мезо-диаминопимелиновой кислоты содержат L-лизин.
Дисахарид представляет собой М-ацетилглюкозамин-М-ацетилмурамовую кислоту (NAG-NAM). 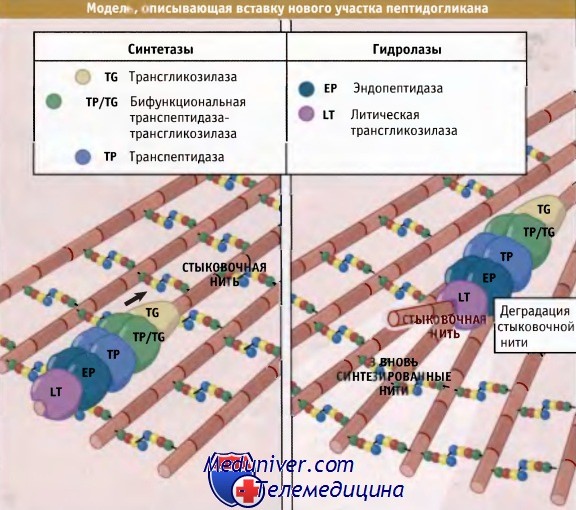 Модель «3 вместо 1» постулирует, что при вставке новообразованных участков пептидогликана реализуется принцип «прежде чем удалить — сделай».
Модель «3 вместо 1» постулирует, что при вставке новообразованных участков пептидогликана реализуется принцип «прежде чем удалить — сделай».
Согласно предложенной модели, мультиферментный комплекс, образующийся на предсуществующей нити (стыковочная нить),
синтезирует три новых сшитых между собой нити и вставляет их в уже имеющийся пептидогликановый слой до момента деградации стыковочной нити. 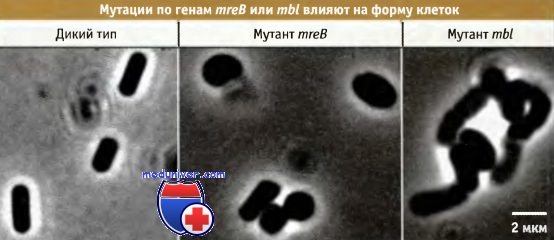 Влияние мутаций в генах mreB и mbl на форму клеток у В. subtilis.
Влияние мутаций в генах mreB и mbl на форму клеток у В. subtilis.
Клетки дикого типа имеют палочковидную форму, mreВ-мутанты, более округленную,
а мутантам по гену mbl свойственна изогнутая форма.  Белок Mbl бактерий, обладающий низкой степенью структурной гомологии с актином эукариот,
Белок Mbl бактерий, обладающий низкой степенью структурной гомологии с актином эукариот,
образует в клетках спирализованные филаментарные структуры.
На фотографии показаны три клетки Bacullus subtilis, экспрессирующие Mbl-GFP.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Что такое пептидогликан в микробиологии
Основные функции клеточной стенки следующие.
• Клеточная стенка защищает бактерии от внешних воздействий, придаёт им характерную форму, поддерживает постоянство внутренней среды и участвует в делении.
• Через клеточную стенку бактерий осуществляется транспорт питательных веществ и выделение метаболитов,
• На поверхности клеточной стенки располагаются рецепторы для бактериофагов, бактериоци-нов и различных химических веществ.
• Структура и состав элементов клеточной стенки определяет антигенную характеристику бактерий (по структуре О- и Vi-Аг).
• Клеточная стенка способна по-разному воспринимать красители; на этом основаны тинкториальные свойства бактерий.
• Нарушение синтеза компонентов клеточной стенки приводит к гибели бактерии или образованию 1-форм.
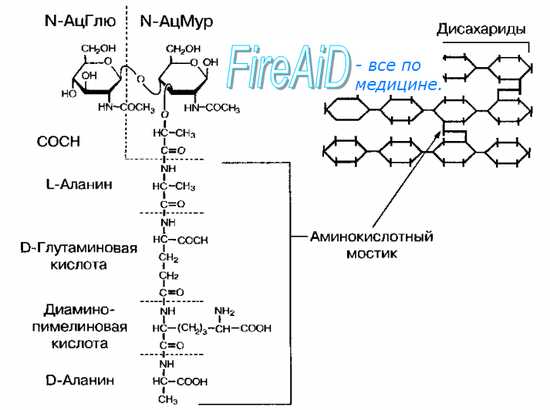 Рис. 4-3. Структура пептидогликана (муреина) Escherichia coll. Гетерополимерные цепочки, состоящие из чередующихся остатков N-ацитилглюкозамина (N-АцГлю) и N-ацетилмурамовой кислоты (N-АцМур), связаны между собой аминокислотными мостиками. Слева— детальное строение фрагмента пептидогликана. Справа — схематическое изображение структуры однослойного муреи-нового мешка.
Рис. 4-3. Структура пептидогликана (муреина) Escherichia coll. Гетерополимерные цепочки, состоящие из чередующихся остатков N-ацитилглюкозамина (N-АцГлю) и N-ацетилмурамовой кислоты (N-АцМур), связаны между собой аминокислотными мостиками. Слева— детальное строение фрагмента пептидогликана. Справа — схематическое изображение структуры однослойного муреи-нового мешка.
Пептидогликан. Муреиновый мешок. Структура пептидогликана (муреина)
Опорный каркас клеточной стенки бактерий — пептидогликан (муреин) — гетерополимер, состоящий из повторяющихся дисахаридных групп, соединённых поперечными и боковыми цепочками (рис. 4-3). Остов молекулы пептидогликана — дисахарид. Его образуют N-ацетилглюкозамин и N-ацетилмурамовая кислота, соединённые через р-гликозидные связи. К молекуле N-ацетилмурамовой кислоты присоединяются олигопептиды, образующие боковые цепочки.
Связывание фрагментов пептидогликана заключается в образовании пептидной связи между терминальным остатком аминокислотного мостика (D-аланином) с предпоследним остатком примыкающего аминокислотного мостика (L-лизином или диаминопимелиновой кислотой в зависимости от вида бактерии). Боковые мостики образуют четыре аминокислоты, поперечные (вертикально связывающие слои пептидогликана) — пять аминокислот. В состав боковых мостиков также входят уникальные аминокислоты, присутствующие только у прокариотов, например диаминопимелиновая кислота (у большинства грамотрицательных бактерий) и D-изомеры глутаминовой кислоты и аланина. Эти соединения — «ахиллесова пята» бактерий, так как препараты, ингибирующие синтез компонентов клеточной стенки, обычно безвредны для растений и животных.
Пептидогликан лабилен к действию различных агентов. В частности, лизоцим гидролизует пептидогликан, расщепляя гликозидные связи между N-ацетилглюкозамином и N-ацетилмурамовой кислотой, то есть лизоцим действует как N-ацетилмурамидаза. Пептидазы расщепляют межпептидные связи, амидазы — связи между N-ацетилмурамовой кислотой и боковым пептидом (L-аланином). Лизостафин разрушает поперечные мостики пептидогликанов стафилококков. В-Лактамные антибиотики нарушают связывание боковых мостиков.
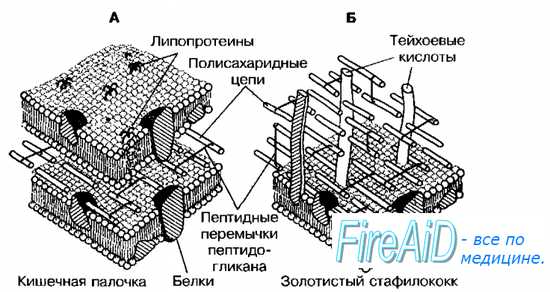 Рис. 4-4. Клеточная стенка грамотрицательных (А) и грамположительных (Б) бактерий.
Рис. 4-4. Клеточная стенка грамотрицательных (А) и грамположительных (Б) бактерий.
Гетерополимерные пептидогликановые цепочки, соединённые пептидными связями, образуют гигантскую мешковидную макромолекулу («муреиновый мешок»), покрывающую всё тело бактерии. На поверхности «муреинового мешка» и в его толще располагаются различные вещества, характер и содержание которых лежит в основе разделения бактерий на грамположительные и грамотрицателъные (рис. 4-4).
Пептидогликан
Пептидогликан (также известный как муреин) — гетерополимер N-ацетилглюкозамина и N-ацетилмурамовой кислоты, сшитый через лактатные остатки N-ацетилмурамовой кислоты короткими пептидными цепочками. Важнейший компонент клеточной стенки бактерий, выполняющий механические функции, осмотической защиты клетки, выполняет антигенные функции. Характерен только для бактерий (в клеточной стенке некоторых архей имеется аналог — псевдопептидогликан). Аминокислотный состав пептидных цепочек является систематическим признаком.
Содержание
Структура


Пептидогликан образует упорядоченную структуру ячеистого строения, построенную из N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенных β-1,4-гликозидными связями. Остатки N-ацетилмурамовой кислоты сшиты между собой при помощи коротких пептидов (сшивка производится ферментом транспептидазой). Типично пептидная цепочка содержит L-аланин, D-глутаминовую кислоту, мезо-диаминопимелиновую кислоту, L-лизин, D-аланин (следует отметить, что мезо-диаминопимелиновая кислота и D-аминокислоты в составе клеточных структур встречаются только в прокариотических клетках). Такая трёхмерная структура сообщает клеточной стенке бактерий прочность и защиту от осмотического лизиса.
Устойчивость пептидогликана
См. также
Ссылки
Примечания
 Углеводы Углеводы | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Общие: | Альдозы · Кетозы · Фуранозы · Пиранозы | ||||||||
| Геометрия | Аномеры · Мутаротация · Проекция Хоуорса | ||||||||
| Моносахариды |
Дезоксисахариды (Дезоксирибоза) | ||||||||
| Гексоза | Кетогексозы (Псикоза, Фруктоза, Сорбоза, Тагатоза) Дезоксисахариды (Фукоза, Фукулоза, Рамноза) | ||||||||
| Гептозы | Кетогептозы (Седогептулоза, Манногептулоза) | ||||||||
| >7 | Октозы · Нанозы (Нейраминовая кислота) | ||||||||
| Дисахариды | Сахароза · Лактоза · Мальтоза · Трегалоза · Тураноза · Целлобиоза · Мелибиоза · Генцибиоза · Вицианоза · Апиоза · Рутиноза |
| Трисахариды | Рафиноза · Мелицитоза · Мальтотриоза · Генцианоза · Солатриоза · Целлотриоза |
| Тетрасахариды | Акарбоза · Стахиоза |
| Олигосахариды | Фруктан · Галактан · Маннан · Изомальтан · Ксилан · Арабан |
| Полисахариды | Гликоген · Крахмал · Целлюлоза · Хитин · Амилоза · Амилопектин · Сахилоза · Инулин · Декстран · Пектин · Галактоманнан · Агароза · Лихенин · Пуллулан |
| Аминосахара · Фосфосахара · Ангидросахара · Гликозиды · N-Гликозиды · Гликали · Гликоны · Енозы · Гликозеены · Гликозаны · Гликаны · Глюканы | |
| Гликозаминогликаны | Гепарин · Гепаринсульфат · Хондроитин · Хондроитинсульфат · Гиалуроновая кислота · Гепаран · Дерматан · Дермантансульфат · Кератан · Кератансульфат · Пептидогликан · Хитозамин · Хондрозамин |
| Аминогликозиды | Канамицин · Стрептомицин · Тобрамицин · Неомицин · Паромомицин · Апрамицин · Гентамицин · Нетилмицин · Амикацин |
 |
 |
 |
Полезное
Смотреть что такое «Пептидогликан» в других словарях:
пептидогликан — пептидогликан … Орфографический словарь-справочник
пептидогликан — сущ., кол во синонимов: 2 • мукопептид (2) • муреин (2) Словарь синонимов ASIS. В.Н. Тришин. 2013 … Словарь синонимов
пептидогликан — мукопептид — [http://www.dunwoodypress.com/148/PDF/Biotech Eng Rus.pdf] Тематики биотехнологии Синонимы мукопептид EN peptidoglycan … Справочник технического переводчика
пептидогликан — glikopeptidas statusas T sritis chemija apibrėžtis Oligopeptido ir polisacharido junginys. atitikmenys: angl. glycopeptide; peptidoglycan; peptidoglycane rus. гликопептид; пептидогликан ryšiai: sinonimas – peptidoglikanas … Chemijos terminų aiškinamasis žodynas
Муреин — Пептидогликан (также известный как муреин) гетерополимер N ацетилглюкозамина и N ацетилмурамовой кислоты, сшитый через лактатные остатки N ацетилмурамовой кислоты короткими пептидными цепочками. Важнейший компонент клеточной стенки бактерий,… … Википедия
КЛЕТОЧНАЯ СТЕНКА — бактерий, специфическая по химич. составу оболочка, окружающая протопласт и тесно связанная структурно функциональными взаимоотношениями с цитоплазматич. мембраной. Толщ. 10 50 нм. Составляет 10 50% сухой массы клеток. У большинства бактерий в… … Биологический энциклопедический словарь
Деление прокариотических клеток — Деление прокариотических клеток процесс образования дочерних прокариотических клеток из материнской. Ключевыми событиями клеточного цикла как прокариот, так и эукариот являются репликация ДНК и деление клетки. Отличительной чертой деления… … Википедия
Иммуномакс — Латинское название Immunomax АТХ: ›› L03AX Иммуностимуляторы другие Фармакологические группы: Биологически активные добавки к пище (БАДы) ›› Иммуномодуляторы Нозологическая классификация (МКБ 10) ›› A49 Бактериальная инфекция неуточненной… … Словарь медицинских препаратов
Археи — Halobacteria, штамм NRC 1, каждая клетка около 5 мкм длиной … Википедия
Гетерополимеры — полимеры, молекулы которых состоят из нескольких разных типов мономеров. Этим они отличаются от гомополимеров, в состав которых входит единственный тип мономеров. К гетерополимерам относятся такие природные биополимеры, как белки, нуклеиновые… … Википедия



