РАНВЬЕ ПЕРЕХВАТ
Смотреть что такое «РАНВЬЕ ПЕРЕХВАТ» в других словарях:
ранвье перехват — (L. A. Ranvier) см. Перехват узла … Большой медицинский словарь
перехват узла — (isthmus nodi, LNH; син. Ранвье перехват) участок истончения оболочки миелинового нервного волокна, соответствующий границе между леммоцитами (шванновскими клетками) … Большой медицинский словарь
Ранвье́ перехва́т — (L.A. Ranvier) см. Перехват узла … Медицинская энциклопедия
Перехват Ранвье (Node Of Ranvier) — сужение миелинового нервного волокна, образующееся на границе между двумя соседними шванновскими клетками. В перехвате отсутствует миелиновая оболочка. Источник: Медицинский словарь … Медицинские термины
ПЕРЕХВАТ РАНВЬЕ — (node of Ranvier) сужение миелинового нервного волокна, образующееся на границе между двумя соседними шванновскими клетками. В перехвате отсутствует миелиновая оболочка … Толковый словарь по медицине
Перехва́т узла́ — (isthmus nodi, LNH; син. Ранвье перехват) участок истончения оболочки миелинового нервного волокна, соответствующий границе между леммоцитами (шванновскими клетками) … Медицинская энциклопедия
МИЕЛИНОВАЯ ОБОЛОЧКА — (от греч. myelos мозг), оболочка, окружающая отростки нервных клеток в мякотных волокнах. М. о. состоит из белого белково липидного комплекса миелина, в периферич. ЦНС образуется вследствие многократного обёртывания отростка шванновской клеткой… … Биологический энциклопедический словарь
Нейрон — (от греч. néuron нерв) неврон, нервная клетка, основная функциональная и структурная единица нервной системы (См. Нервная система); принимает сигналы, поступающие от рецепторов и др. Н., перерабатывает их и в форме нервных импульсов… … Большая советская энциклопедия
НЕЙРОН — (от греч. neuron жила, нерв), нервная клетка, нейроцит, осн. структурная и функциональная единица нервной системы, обладающая специфич. проявлениями возбудимости. Способен принимать сигналы, перерабатывать их в нервные импульсы и проводить к… … Биологический энциклопедический словарь
НЕРВНОЕ ОКОНЧАНИЕ — (terminatio nervi), специализированное образование в концевом разветвлении отростков нейрона, лишённых миелиновой оболочки; служит для приёма или передачи сигналов. Чувствительные, или сенсорные, Н. о,, осуществляющие приём сигналов (рецепцию),… … Биологический энциклопедический словарь
Обёртка для аксона
Аксон — дело тонкое. Ему постоянно нужна поддержка, осуществляемая шванновскими клетками и олигодендроцитами.
Автор
Редакторы
Очень часто при описании нервной системы используются «электрические» термины: например, нервы сравниваются с проводами. Это потому, что по нервному волокну действительно перемещается электрический сигнал. Каждому из нас известно, что оголенный провод опасен, ведь он бьет током, и по этой причине люди пользуются изоляционными материалами, не проводящими электричество. Природе тоже не чужда техника безопасности, и нервные «провода» она обматывает своим собственным изолирующим материалом — миелином.
Сложная обёртка
Миелин окружает отростки нервных клеток, изолируя их от внешнего воздействия. Это необходимо для более надежной и быстрой передачи сигнала по нервной системе. Благодаря изоляции нервного волокна электрический сигнал не рассеивается и добирается до места назначения без помех. Скорость прохождения сигнала по миелиновым и безмиелиновым волокнам может отличаться на три порядка: от 70 до 140 м/с и от 0,3 до 0,5 м/с соответственно.
По сути миелин — это клеточная мембрана глиальных клеток, многократно обмотанная вокруг аксона. Сама мембрана на 70–75% состоит из липидов и на 25–30% — из белков. В периферической нервной системе донором мембран становятся шванновские клетки, а в центральной — олигодендроциты. Эти клетки бережно обматывают своими мембранами ценные каналы связи, чтобы обеспечить надежное взаимодействие нервной системы и периферических органов. Миелин покрывает нервное волокно не целиком: существуют промежутки между наслоениями миелина, называемые перехватами Ранвье (рис. 1). Есть прямая зависимость между расстоянием от одного промежутка до другого и скоростью распространения нервного импульса по волокну: чем больше расстояние между перехватами Ранвье, тем выше скорость передачи сигнала в нерве [1].
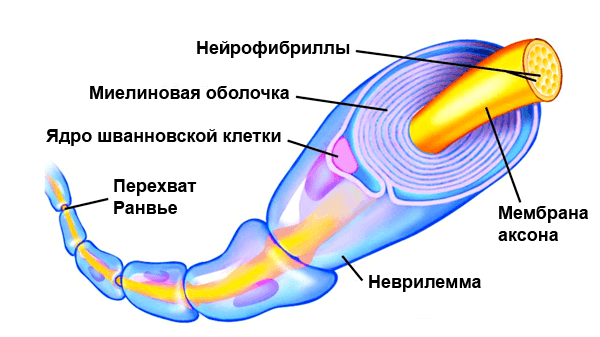
Рисунок 1. Нервное волокно, обернутое миелином. Видны ядра шванновских клеток (nucleus of Schwann cell) и перехваты Ранвье (nodes of Ranvier) — участки аксона, которые не покрыты миелиновой оболочкой.
Если говорить о белках, входящих в состав миелина, то надо уточнить, что это не только простые белки. В миелине встречаются гликопротеины — белки, к которым присоединены короткие углеводные последовательности. Важной составляющей миелина является главный структурный белок миелина (myelin basic protein, MBP), впервые выделенный около 50 лет назад. MBP — это трансмембранный белок, который может многократно «прошивать» липидный слой клетки. Его различные изоформы (рис. 2) кодируются геном под названием Golli (gene in the oligodendrocyte lineage). Структурной основой миелина служит изоформа массой 18,5 килодальтон [2].
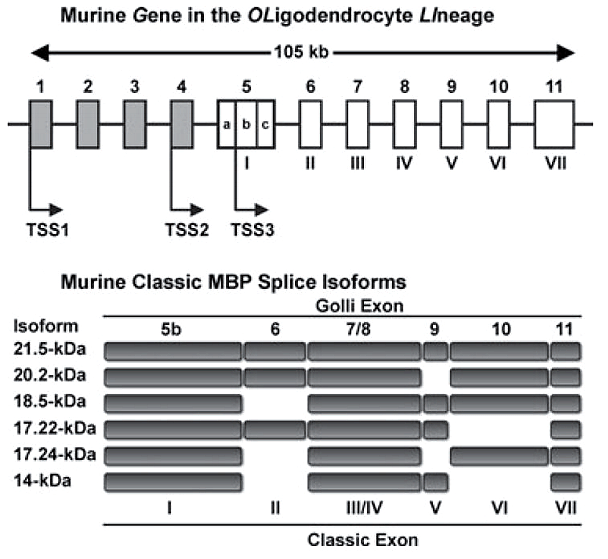
Рисунок 2. Различные изоформы основного белка миелина (MBP) создаются на основе одного и того же гена. Например, для синтеза изоформы массой 18,5 кДа используются все экзоны, кроме экзона II.
В состав миелина входят сложные липиды цереброзиды. Они представляют собой аминоспирт сфингозин, соединенный с жирной кислотой и остатком углевода. В синтезе липидов миелина принимают участие пероксисомы олигодендроцитов. Пероксисомы — это липидные пузырьки с различными ферментами (в общей сложности известно около 50 видов пероксисомных энзимов). Эти органеллы занимаются, в частности, β-окислением жирных кислот: жирных кислот с очень длинной цепью (very long chain fatty acids, VLCFA), некоторых эйкозаноидов и полиненасыщенных жирных кислот (ПНЖК, polyunsaturated fatty acids, PUFAs). Поскольку миелин может содержать до 70% липидов, пероксисомы крайне важны для нормального метаболизма этого вещества. Они используют N-ацетиласпартат, вырабатываемый нервной клеткой, для постоянного синтеза новых липидов миелина и поддержания его существования. Кроме этого, пероксисомы принимают участие в поддержании энергетического метаболизма аксонов [3].
Важная обёртка
Миелинизация (постепенная изоляция нервных волокон миелином) начинается у людей уже в эмбриональном периоде развития. Первыми этот путь проходят подкорковые структуры. В течение первого года жизни происходит миелинизация отделов периферической и центральной нервной системы, отвечающих за двигательную активность. Миелинизация участков головного мозга, регулирующих высшую нервную деятельность, заканчивается к 12–13 годам. Из этого видно, что миелинизация тесно связана со способностью отделов нервной системы осуществлять специфические для них функции. Вероятно, именно активная работа волокон до рождения запускает их миелинизацию.
Дифференцировка клеток — предшественниц олигодендроцитов зависит от ряда факторов, связанных с работой нейронов. В частности, работающие отростки нейронов могут выделять белок нейролигин 3, который способствует пролиферации и дифференциации клеток-предшественниц [4]. В дальнейшем созревание олигодендроцитов происходит за счет ряда других факторов. В статье с характерным названием «Насколько велик миелинизирующий оркестр?» описывается происхождение олигодендроцитов в разных частях мозга [5]. Во-первых, в различных частях мозга олигодендроциты начинают созревать в разное время. Во-вторых, за их созревание отвечают разные клеточные факторы, что тоже зависит от региона нервной системы (рис. 3). У нас может возникнуть вопрос: а сходны ли между собой олигодендроциты, появившиеся с таким расхождением в стартовых данных? И насколько схож у них миелин? В целом, авторы статьи считают, что между популяциями олигодендроцитов из разных участков головного мозга действительно существуют различия, и обусловлены они во многом именно местом закладки клеток, воздействием на них окружающих нейронов. И всё же типы миелина, синтезируемые разными пулами олигодендроцитов, не имеют настолько больших отличий, чтобы они не были взаимозаменяемыми.
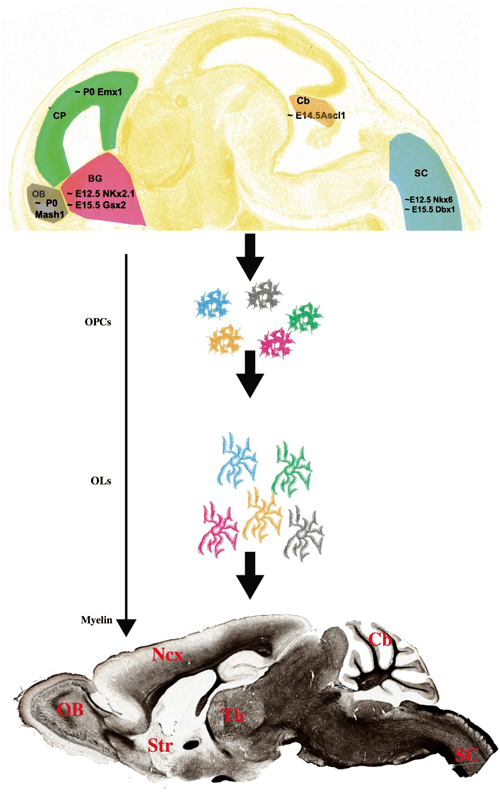
Рисунок 3. Различия во времени закладки олигодендроцитов в разных отделах головного мозга и в клеточных факторах, влияющих на их развитие.
Сам процесс миелинизации нервных волокон в центральной нервной системе происходит следующим образом (рис. 4). Олигодендроциты выпускают несколько отростков к аксонам разных нейронов. Входя с ними в контакт, отростки олигодендроцитов начинают оборачиваться вокруг них и расползаться по длине аксона. Количество оборотов постепенно увеличивается: в некоторых участках ЦНС их число доходит до 50. Мембраны олигодендроцитов становятся всё более тонкими, распространяясь по поверхности аксона и «выдавливая» из себя цитоплазму. Чем раньше слой миелина был обернут вокруг нервного окончания, тем более тонким он будет. Самый внутренний слой мембраны остается довольно толстым — для осуществления метаболической функции. Новые слои миелина наматываются поверх старых, перекрывая их так, как показано на рисунке 4 — не только сверху, но и увеличивая площадь аксона, покрытую миелином.

Рисунок 4. Миелинизация нервного волокна. Мембрана олигодендроцита наматывается на аксон, постепенно уплотняясь с каждым оборотом. Внутренний, прилегающий к аксону слой мембраны остается относительно толстым, что необходимо для выполнения метаболической функции. На разных частях рисунка (а-в) с разных ракурсов показано постепенное наматывание новых слоев миелина на аксон. Красным цветом выделен более толстый, метаболически активный слой, синим — новые уплотняющиеся слои. Внутренний слой миелина (inner tongue на части б) охватывается всё новыми и новыми слоями мембраны не только сверху, но и по бокам (в), вдоль аксона.
Миелинизация нервных волокон олигодендроцитами также значимо зависит от белка нейрегулина 1. Если он не воздействует на олигодендроциты, то в них запускается программа миелинизации, не учитывающая активность нервной клетки. Если же олигодендроциты получили сигнал от нейрегулина 1, то далее они начнут ориентироваться на работу аксона, и миелинизация будет зависеть от интенсивности выработки глутамата и активации им специфических NMDA-рецепторов на поверхности олигодендроцитов [6]. Нейрегулин 1 — ключевой фактор для запуска процессов миелинизации и в случае шванновских клеток [7].
Изменчивая обёртка
Миелин постоянно образуется и разрушается в человеческом организме. На синтез и распад миелина могут влиять факторы, связанные с особенностями внешней среды. Например, воспитание. С 1965 по 1989 год Румынией руководил Николае Чаушеску. Он установил жесткий контроль над репродуктивным здоровьем и институтом брака в своей стране: усложнил процедуру развода, запретил аборты и ввел ряд стимулов и льгот для женщин, имевших более пяти детей. Итогом этих мер стало ожидаемое повышение рождаемости. Вместе с рождаемостью увеличилось количество криминальных абортов, не добавивших здоровья румынкам, и возросло количество детей-отказников. Последние воспитывались в детских домах, где с ними не очень-то активно общался персонал. Румынские дети в полной мере ощутили на себе то, что называется социальной депривацией — лишение возможности полноценного общения с другими людьми. Если речь идет о маленьком ребенке, то следствиями социальной депривации станут нарушение формирования эмоциональных привязанностей и расстройство внимания. Когда режим Чаушеску пал, западным ученым предстояло в полной мере оценить результат социальной политики этого диктатора. Румынских детей, имеющих выраженные проблемы со вниманием и установкой социальных контактов, впоследствии стали называть детьми Чаушеску.
Кроме различий при выполнении нейропсихологических тестов, у детей Чаушеску по сравнению с детьми, не находившимися в таких условиях, отличалось даже строение головного мозга [8]. При оценке состояния белого вещества мозга ученые используют показатель фрактальной анизотропии. Он позволяет оценить плотность нервных волокон, диаметр аксонов и их миелинизацию. Чем больше фрактальная анизотропия, тем разнообразнее волокна, которые встречаются в этой области мозга. У детей Чаушеску отмечалось снижение фрактальной анизотропии в пучке белого вещества, соединяющего височную и лобную доли в левом полушарии, то есть связи в этом регионе были недостаточно сложными и разнообразными, с нарушениями миелинизации. Такое состояние связей мешает нормальному проведению сигналов между височной и лобной долями. В височной доле находятся центры эмоционального реагирования (миндалина, гиппокамп), а орбитофронтальная кора лобной доли также связана с эмоциями и принятием решений. Нарушение формирования связей между этими отделами мозга и проблемы в их работе в итоге приводили к тому, что выросшие в детдомах дети испытывали трудности в установлении нормальных отношений с другими людьми.
На миелинизацию также может влиять и состав еды, которую дают ребенку. При белково-энергетической недостаточности питания отмечается снижение образования миелина. Недостаток жирных кислот тоже отрицательно сказывается на синтезе этого ценного вещества, так как оно больше чем на 2/3 состоит из липидов. Дефицит железа, йода и витаминов группы В приводит к снижению образования миелина [9]. В основном эти данные были получены при изучении лабораторных животных, но история, к сожалению, дала людям возможность оценить влияние недостатка еды и на формирующийся мозг ребенка [10]. Голодная зима (голл. hongerwinter) 1944–1945 гг. в Нидерландах привела к тому, что родилось множество детей, чьи матери плохо питались. Оказалось, что в условиях голодания мозг этих детей формировался с нарушениями. В частности, наблюдалось большое количество нарушений именно в белом веществе, то есть возникали проблемы с формированием миелина. В итоге это приводило к разнообразным психическим расстройствам.
Поврежденная обёртка
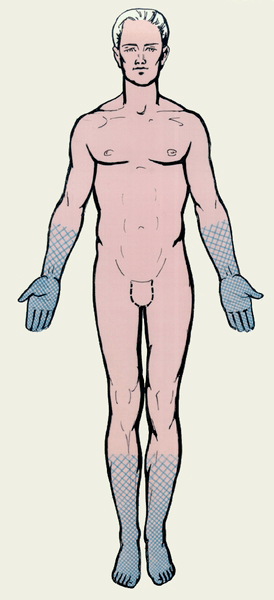
Рисунок 5. Нарушение чувствительности по полиневритическому типу. Название «носки — перчатки» связано с тем, что анатомические зоны, соответствующие поражению нервов, похожи на области, покрываемые этими предметами одежды.
Как мне кажется, для человеческого организма вполне подходит следующее правило: если есть орган, значит, к нему должна быть болезнь. В принципе, это правило можно расширить до молекулярных процессов: есть процесс — есть и болезни, связанные с нарушением этого процесса. В случае с миелином это демиелинизирующие заболевания. Их довольно много, но подробнее я расскажу о двух — синдроме Гийена-Барре и рассеянном склерозе. При этих расстройствах повреждение миелина приводит к нарушению адекватного проведения сигнала по нервам, что и обуславливает симптомы болезни.
Синдром Гийена-Барре (СГБ) — это заболевание периферической нервной системы, при котором происходит разрушение миелиновой оболочки, формируемой шванновскими клетками. СГБ является классическим аутоиммунным заболеванием. Как правило, ему предшествует инфекция (часто — вызванная микробом Campylobacter jejuni). Присутствие различных возбудителей в организме человека запускает аутоиммунное повреждение миелина нервных волокон T- и B-лимфоцитами. Клинически это проявляется мышечной слабостью, нарушением чувствительности по типу «носки — перчатки» (полиневритический тип) (рис. 5). В дальнейшем мышечная слабость может нарастать вплоть до полного паралича конечностей и поражения туловищной мускулатуры. Поражения чувствительной нервной системы также могут быть разнообразны: от снижения способности различать собственные движения (нарушение глубокой чувствительности) до выраженного болевого синдрома. При тяжелых формах СГБ главную опасность представляет потеря способности к самостоятельному дыханию, требующая подключения к аппарату искусственной вентиляции легких (ИВЛ). Для лечения СГБ в настоящее время используют плазмаферез (очистку плазмы от вредных антител) и внутривенные вливания препаратов человеческого иммуноглобулина для нормализации иммунного ответа. В большинстве случаев лечение приводит к стойкому выздоровлению.
Рассеянный склероз (РС) заметно отличается от СГБ. Во-первых, это демиелинизирующее заболевание приводит к поражению центральной нервной системы, то есть затрагивает миелин, синтезируемый олигодендроцитами. Во-вторых, с причинами РС до сих пор много неясного: слишком большое разнообразие генетических и средовых факторов задействовано в патогенезе заболевания. Принципиальный момент в запуске РС — нарушение непроницаемости гематоэнцефалического барьера (ГЭБ) для иммунных клеток. В норме ткань мозга отгорожена от всего остального организма этим надежным фильтром, который не пропускает к ней множество веществ и клеток, в том числе иммунных. ГЭБ появляется уже в эмбриональном периоде развития, изолируя ткань мозга от формирующейся иммунной системы. В это время иммунная система человека «знакомится» со всеми существующими тканями, чтобы в дальнейшем, при взрослой жизни, не нападать на них. Мозг и ряд других органов остаются «не представленными» иммунной системе. При нарушении целостности ГЭБ иммунные клетки получают возможность для атаки незнакомых ей тканей мозга. В-третьих, РС отличается более тяжелыми симптомами, которые требуют других терапевтических подходов. Симптоматика зависит от того, где локализуются повреждения нервной системы (рис. 6 и 7). Это может быть шаткость походки, нарушения чувствительности, различные когнитивные симптомы. Для лечения РС используются высокие дозы глюкокортикоидов и цитостатики, а также препараты интерферона и специфические антитела (натализумаб). По-видимому, в дальнейшем будут развиваться новые методы лечения РС, основанные непосредственно на восстановлении миелиновой оболочки в поврежденных участках мозга. Ученые указывают на возможность трансплантации клеток — предшественниц олигодендроцитов или усиления их роста за счет введения инсулиноподобного фактора роста или тиреоидных гормонов [11]. Однако это еще впереди, а пока неврологам недоступны более «молекулярные» методы лечения.
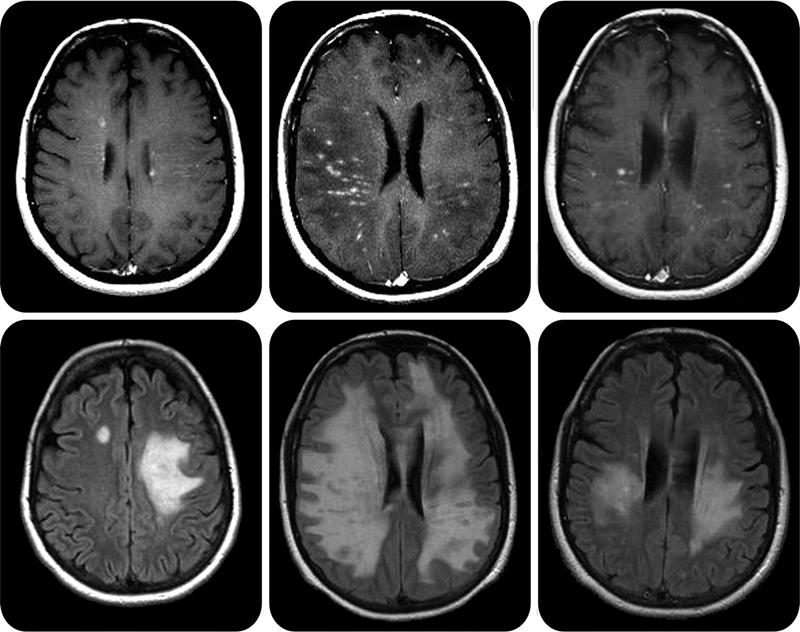
Рисунок 6. Очаги поражения центральной нервной системы при рассеянном склерозе на МРТ выглядят как белые бляшки.
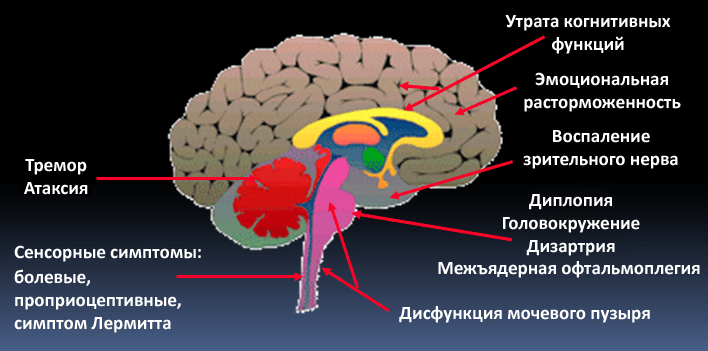
Рисунок 7. В зависимости от места поражения нервной системы при рассеянном склерозе может быть разная симптоматика: от тремора и атаксии при повреждении мозжечка до эмоциональных расстройств при локализации очагов в лобных долях.
Что такое перехват ранвье
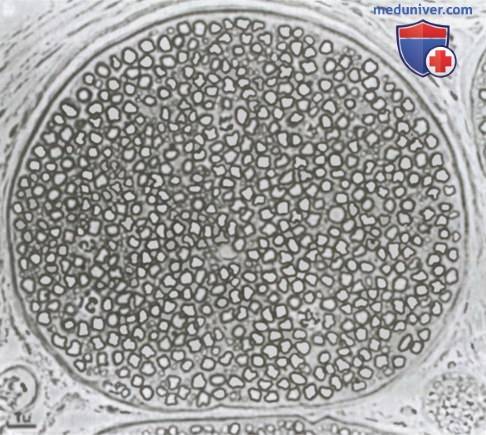 Поперечное сечение небольшого нервного ствола, содержащего миелинизированные и немиелинизированные волокна.
Поперечное сечение небольшого нервного ствола, содержащего миелинизированные и немиелинизированные волокна. 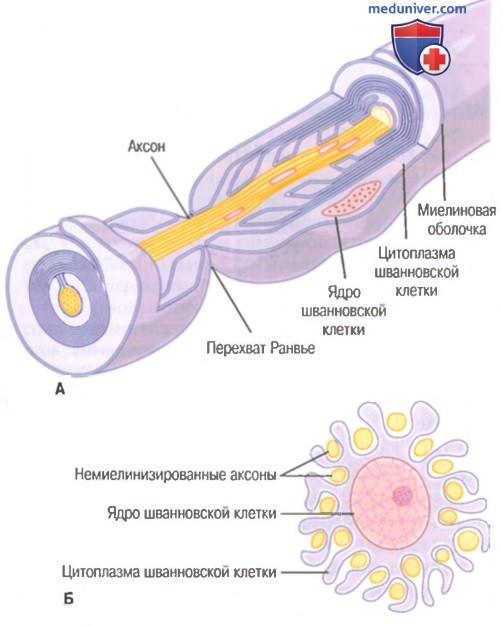 Значение шванновских клеток для изоляции нервных волокон.
Значение шванновских клеток для изоляции нервных волокон.
А. Накручивание мембраны шванновской клетки на крупный аксон формирует миелиновую оболочку миелинизированного нервного волокна.
Б. Частичная закрутка мембраны и цитоплазмы шванновской клетки вокруг множества немиелинизированных нервных волокон (показано в поперечном сечении).
На рисунке показано типичное миелинизированное волокно. Центральной его частью является аксон, по мембране которого проводится потенциал действия. Аксон заполнен аксоплазмой — вязкой внутриклеточной жидкостью. Аксон окружен миелиновой оболочкой, которая часто много толще, чем сам аксон. Примерно через каждые 1-3 мм вдоль миелиновой оболочки имеется перехват Ранвье.
Миелиновая оболочка формируется вокруг аксона шванновскими клетками. Мембрана шванновской клетки сначала охватывает аксон, затем шванновская клетка многократно вращается вокруг аксона, образуя многочисленные мембранные слои, содержащие липидное вещество сфингомиелин. Это вещество является отличным изолятором и снижает ионный ток через мембрану аксона примерно в 5000 раз. Между каждыми двумя последовательно расположенными шванновскими клетками по ходу аксона остается маленькая неизолированная область длиной всего 2-3 мкм, где ионы могут свободно переходить через мембрану аксона из внеклеточной жидкости во внутриклеточную и обратно. Эту область называют перехватом Ранвье.
Сальтаторное проведение в миелиновых волокнах от перехвата к перехвату. Ионы практически не могут проходить через толстую миелиновую оболочку мякотных волокон, однако они легко диффундируют через перехваты Ранвье. Следовательно, потенциалы действия возникают только в перехватах и проводятся от перехвата к перехвату; это называют салътаторным (скачкообразным) проведением. В этом случае электрический ток течет через внеклеточную жидкость снаружи от миелиновой оболочки, а также через аксоплазму внутри аксона от перехвата к перехвату, последовательно возбуждая один перехват за другим. Таким образом, нервный импульс как будто прыгает по волокну, на основании этого и появился термин «салътаторное проведение».
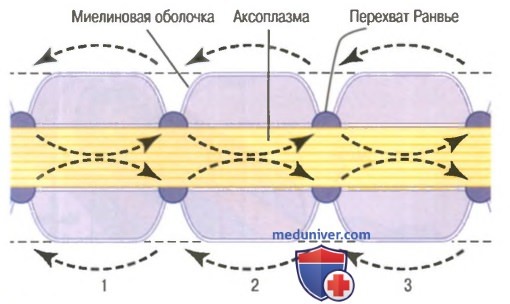 Сальтаторное проведение по миелинизированному аксону. Стрелки указывают направление движения электрического тока от перехвата к перехвату.
Сальтаторное проведение по миелинизированному аксону. Стрелки указывают направление движения электрического тока от перехвата к перехвату.
Сальтаторное проведение имеет два преимущества. Во-первых, заставляя процесс деполяризации «прыгать» через большие промежутки вдоль аксона, этот механизм повышает скорость проведения в миелинизированных волокнах в 5-50 раз. Во-вторых, сальтаторное проведение сохраняет энергию для аксона, поскольку деполяризуются только перехваты, что позволяет приблизительно в 100 раз снизить потерю ионов по сравнению с возможными потерями в других условиях. В связи с этим снижаются траты энергии, необходимые для восстановления трансмембранной разности концентраций ионов натрия и калия после серии нервных импульсов.
Существует другая особенность сальтаторного проведения в крупных миелинизированных волокнах: отличная изоляция, обеспечиваемая миелиновой оболочкой, и 50-кратное снижение мембранной емкости позволяют осуществлять реполяризацию путем перемещения очень незначительного числа ионов.
Скорость проведения в нервных волокнах. Скорость проведения в нервных волокнах колеблется от 0,25 м/сек в очень тонких немиелинизирован-ных волокнах до 100 м (длина футбольного поля) в 1 сек в очень толстых миелинизированных волокнах.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Молекулярная специфика в перехватах Ранвье
| Эволюционная необходимость быстрой и эффективной проводимости потенциалов действия в нейронах позвоночных привела к образованию миелиновой оболочки. Миелин – многослойная оболочка, сформированная в центральной нервной системе олигодендроцитами, а в периферической нервной системе Шванновскими клетками – обертывает сегменты аксона, разделенные узловыми перехватами Ранвье (РИС.1а). Миелиновая оболочка ослабляет прохождение current flow через мембрану аксона посредством снижения ее емкости (capacitance) и увеличения ее transverse resistance, давая, таким образом, возможность быстрому сальтаторному продвижению нервного импульса от перехвата к перехвату. В результате большое число аксонов с высокой скоростью проводимости может быть заключено в ограниченное пространство. И этот признак дает возможность для развития более сложной нервной системы. Кроме того, сальтаторная проводимость устраняет необходимость восстановления потенциала действия в каждой точке мембраны аксона. Следовательно, снижаются метаболические требования для активности нейронов. |
Organization and function of the nodal environs
Скоординированная дифференцировка аксона и его миелинизирующих клеток требует тесной взаимосвязи между нейронами и глией на самых ранних стадиях развития. Сигналы, передаваемые аксоном, регулируют пролиферацию, выживаемость и дифференцировку олигодендроцитов и Шванновских клеток и участвуют в детерминации толщины миелина. Реципрокные глиальные сигналы влияют на цитоскелет аксонов и аксонный транспорт и необходимы для выживаемости аксонов. В результате таких реципрокных связей миелинизированные волокна приобретают структурные признаки, позволяющие им максимизировать скорость проведения импульса. Одним из таких признаков является разделение мембраны аксона на отдельные молекулярные, структурные и функциональные домены. Такие домены включают перехваты Ранвье, paranodal junction (околоузловое соединение), juxtaparanodes (участок, расположенный за paranodal junction ) и internodal (межперехватный участок (РИС.1b и 2). Авторы сфокусировали внимание на молекулярных механизмах, лежащих в основе образования и поддержания этих уникальных доменов аксона, необходимых для нормального функционирования нервной системы.
The node of Ranvier
Узловые перехваты Ранвье представляют собой небольшие по длине, равномерно расположенные разрывы миелиновой оболочки. Интервал между перехватами Ранвье имеет длину примерно в 100 раз большую диаметра нервного волокна. Имеются некоторые различия в структурных характеристиках перехватов между центральной и периферической нервными системами. В периферических нервах весь миелинизированный участок аксона покрыт базальной оболочкой (базальным слоем) (basal lamina), а внешний слой (outer collar) шванновской клетки имеет микроворсинки, покрывающие узловые перехваты (РИС. 1b). Околоузловое пространство (т.е. пространство между аксолеммой и базальным слоем), содержащее микроворсинки, также заполнено волокнистым матриксом. В ЦНС нет basal lamina и контакт перехватов осуществляется через околоузловые астроциты (perinodal astrocyte=перинодальные астроциты), которые недавно были названы синантоцитами (synantocytes) [Butt, A. M., Kiff, J., Hubbard, P. & Berry, M. Synantocytes: new functions for novel NG2 expressing glia. J. Neurocytol. 31, 551-565. 2002].
Cytoskeletal proteins.Перехваты и начальный сегмент аксона обогащены ankyrin G – мембранно-цитоскелетным адаптером, связывающим интегральные мембранные белки с spectrin cytoskeleton. Ankyrin G взаимодействует с Na+ каналами – с их α- и β-субъединицами, а также с Nf 186, Nrcam и Kv3.1. β-субъединица «вербует» ankyrin G для плазменной мембраны и такое взаимодействие регулируется tyrosine phosphorylation. Связывание ankyrin G с α-субъединицей осуществляется через последовательность девяти аминокислот, присутствующих во всех известных voltage-gated Na+ каналах. Такой девяти-аминокислотный лейтмотив необходим для аккумуляции α-субъединицы в начальном сегменте аксона. Более того, такой ankyrin-binding сайт локализован в пределах короткой последовательности, которой достаточно, чтобы таргетировать белки для начального сегмента аксона. Остается определить, нужна ли эта короткая последовательность для таргетирования перехватов Ранвье. Связывание ankyrin G с двумя перехватными Ig-CAMs, Nf186 и Nrcam опосредуется группой из двенадцати аминокислот, найденной в их цитоплазматических доменах. Ankyrin G связывается с этими группами только тогда, когда он дефосфорилирован, указывая на то, что неидентифицируемые тирозин киназы и фосфатазы могут регулировать такое взаимодействие. Tyrosine-phosphorylated neurofascin локализован в глиальных параузлах (paranodes), а не в перехватах, свидетельствуя в пользу гипотезы, согласно которой перехватный neurofascin тесно связан с ankyrin G. Ankyrin G также связывает spectrin βIV – spectrin изоформу, присутствующую в изобилии в перехватах Ранвье и начальных сегментах аксонов – связывая в дальнейшем узловые Na + каналы и Ig-CAMs с цитоскелетом аксона.
The nodal gap, extracellular matrix and the glial membrane В ПНС узловой пробел (перехват) заполнен микроворсинками Шванновских клеток, происходящими из наружной оболочки клетки (РИС.1b). В проксимальной области микроворсинок мембраны двух соседних Шванновских клеток соединены tight junctions (плотным соединением), но эти соединения не изолируют узелковый пробел. Было обнаружено, что они проницаемы для пероксидазы хрена, прикладываемой снаружи нервного волокна. Три белка – erzin, radixin и moesin, так же как и erzin-связывающий белок EBP50 и Rho-AGTPasa локализованы в микроворсинках. Потенциально эти белки могут связывать актин-обогащенные микроворсинки с интегральными мембранными белками (integral membrane proteins). Кроме того, несколько белков внеклеточного матрикса (extracellular matrix (ECM) proteins) найдено в узловых пробелах под базальной оболочкой, включая hyaluronan-binding proteoglycan versican, tenascin-C и NG2 proteoglycan. Недавно было показано, что dystroglycan в избытке экспрессирующийся на abaxonal поверхности миелинизирующих Шванновских клеток также локализуется в перехватах. Специфическое удаление dystroglycan в Шванновских клетках приводит к дезорганизации микроворсинок, заметной редукции узловых Na+ каналов и к последующему нарушению проводимости нервного волокна.
В отличие от ПНС, в ЦНС отростки перинодальных астроцитов (perinodal astrocytes) (РИС.1b) контактируют с большинством узловых перехватов. Было показано, что узловой пробел в ЦНС включает несколько протеогликанов, в том числе tenascin и phosphacan. Узловые пробелы ЦНС также содержат versican-binding protein Bral1, продуцируемый нейронами. Функции этих белков пока неясны, однако предполагают, что благодаря высокому содержанию в них acidic disaccharides, они могут обеспечивать негативную среду, служащую в качестве внеклеточного резервуара Na+ в перинодальном пространстве. И tenascin-C, и tenascin-R связываются с Na+ каналами и изменяют их электрофизиологические свойства. Генетическое удаление tenascin-R ведет к снижению нервной проводимости, но не оказывает никакого эффекта на распределение Na+ каналов в узловых перехватах, указывая на то, что такое взаимодействие может стабилизировать узловые комплексы или регулировать активность каналов, но не требуется для первичной кластеризации этих каналов. Сообщалось также, что Na+ каналы связывают цитоплазматический конец (tail) и внеклеточный домен Rptpβ – рецепторную тирозин фосфатазу, о локализации которой в узловой аксолемме (nodal axolemma) пока не сообщалось. Более того, значимость этих взаимодействий для нормальной физиологии миелинизированных нервов пока неясна, т.к. распределение узловых Na+ каналов и скорость проводимости миелинизированных аксонов в ЦНС нормальны у Rptpβ-дефицитных мышей. Стоит отметить, что и tenascin-R, и Rptpβ также взаимодействуют с contactin и Nrcam, присутствующими в узловых перехватах ЦНС, указывая на возможное присутствие крупных макромолекулярных комплексов в перинодальном пространстве.
The axoglial paranodal junction
Morphology and molecular composition. C обеих сторон перехватов Ранвье компактная миелиновая оболочка раскрывается и формирует заполненные цитоплазмой глиальные петли (glial loops), которые спирально наматываются вокруг аксона (РИС.1b). Такие паранодальные петли соединены с аксолеммой серией перемычек (поперечных полосок – transverse bands), которые напоминают septate junctions (перегородчатые соединения) у беспозвоночных (BOX1). Аксоглиальные соединения появляются относительно поздно во время миелинизации – сначала они образуются ближе к перехватам наиболее отдаленной паранодальной петлей и постепенно распространяются по мере того, как дополнительные петли прикрепляются к аксону. В итоге, они состоят из множества колец, каждое из которых представляет собой виток миелиновой оболочки.
Мембрана аксона в месте аксоглиального соединения содержит комплекс из двух cell-recognition молекул — contactin-associated protein (Caspr, также известные как paranodin) и contactin (РИС.2b). Caspr является трансмембранным белком типа I, принадлежащим к отдельной подгруппе neurexins – полиморфного белкового семейства, участвующего в клеточной адгезии и межклеточных связях. У человека известно 5 генов семейства Caspr – CASPR1–CASPR5 (Spiegel, I., Salomon, D., Erne,et. al. Caspr3 and caspr4, two novel members of the caspr family are expressed in the nervous system and interact with PDZ domains. Mol. Cell. Neurosci. 20, 283-297. 2002). У дрозофилы известно 2 гена – nrxIV и axo и у Caenorhabditis elegans также 2 гена – itx-1 и nlr-1 [L. Haklai-Topper and E.P., unpublished observations]. Эти белки связывают некоторые CAMs и, следовательно, могут рассматриваться как CAM-ассоциированные белки. Их внеклеточная область состоит из нескольких доменов, участвующих в межбелковых взаимодействиях, включая discoidin и fibrinogen-like domain, epidermal growth factor (EGF) motifs, и несколько областей гомологичных G домену laminin A (BOX 1). Caspr, но не другие члены семейства Caspr, формирует комплекс с contactin только в cis. Взаимодействие между Caspr и contactin необходимо для эффективного экспорта Caspr из эндоплазматического ретикулума к плазменной мембране, оно регулирует glycosylation и транспорт contactin. Caspr и contactin связаны в эндоплазматическом ретикулуме и могут быть транспортированы через Гольджи-независимый путь (Golgi-independent pathway) к клеточной поверхности. Так же как и при исследовании in vitro, у contactin-дефицитных мышей Caspr сохраняется в теле нейрона и не достигает аксона (Boyle, M. E. et al.2001), но Caspr необходим для поддержания contactin в paranodes (Gollan, L. et al. Retention of a cell adhesion complex at the paranodal junction requires the cytoplasmic region of Caspr. J. Cell Biol. 157, 1247-1256 (2002). Shows that the stabilization of the Caspr/contactin complex at the cell membrane requires the intracellular region, which includes the protein 4.1B-binding domain).
И Caspr, и contactin важны для образования axoglial junction (аксоно-глиального соединения) и их отсутствие ведет к исчезновению септ (перемычек) и расширению пространства между аксоном и паранодальными петлями. Результаты указывают, что Caspr и contactin могут быть частью paranodal adhesion complex, который необходим для плотного соединения двух оболочек. Фенотип contactin-дефицитных мышей сходен с фенотипами двух других паранодальных мутантов – galactolipids-дефицитными мышами, у которых отсутствует UDP-galactose ceramide galactosyltransferase (Cgt) и не синтезируются galactocerebroside (GalC) и sulfatide и cerebroside sulfotransferase ( Cst)-null мышами, у которых отсутствует синтез только sulfatide. У всех этих мутантов Caspr и contactin отсутствуют в paranodes. Путь, при котором отсутствие GalC и sulfatide вызывает паранодальные аномалии, неясен, но можно предположить, что это результат прямого связывания sulfatide с Caspr/contactin комплексом. И, напротив, говоря о роли galactolipids в формировании LIPLD RAFTS (липидных плотиков) и организации миелина, можно предположить, что их отсутствие может привести к превращению junctional glial компонентов в не-компактный миелин. Последнее предположение подтверждается недавними находками, показавшими, что генетическое удаление миелинового и лимфоцитарного (myelin and lymphocyte – MAL) белка, – raft-ассоциированной молекулы, участвующей во внутриклеточном трафике, – ведет к паранодальным аномалиям (N. Schaeren-Wiemers and U. Suter, personal communication).
Внутриклеточные области Caspr и Caspr2 содержат juxtamembrane последовательность, которая связывает protein 4.1B, присутствующий в paranodes и juxtaparanodes. Как и другие 4.1 белки, 4.1В белок содержит консервативный actin–spectrin-binding домен и, следовательно, может связывать Caspr (и следовательно contactin) с цитоскелетом. В соответствии с этой гипотезой, protein 4.1B аномально распределяется вдоль периферических миелинизорованных аксонов у мышей с отсутствием либо contactin, либо galactolipids, у которых отсутствуют паранодальные Caspr. У этих мутантов местоположение белка 4.1В коррелирует с положением Caspr и Caspr2, указывая на то, что они могут определять его локализацию. Более того, цитоплазматический конец Caspr нужен для стабилизации Caspr/contactin комплекса в paranodes, т.к. у Caspr мутантов с отсутствием этого домена не сохраняется axoglial junction. Поэтому, вероятно, что Caspr является трансмембранным скаффолдом, который стабилизирует Caspr/contactin adhesion complex в septate-подобных соединениях связывая комплекс с аксонным цитоскелетом через белок 4.1В. Этот механизм напоминает функцию neurexin IV Drosophila, который захватывает Coracle (гомолог белка 4.1) для septate junctions (BOX 1). Кроме того, цитоплазматический Caspr также связывает FERM domain (four-point-one, ezrin–radixin–moesin)-содержащий белок Schwanomin/merlin. Однако значение такого взаимодействия не совсем ясно, поскольку Schwanomin не концентрируется в области паранодального соединения.
Распределение Caspr и contactin вдоль internodes (межперехватов) (см. далее в тексте), их накопление в paranodes (как число колец, представляющих витки миелиновой обертки во время развития), аномальное распределение Caspr при рассеянном склерозе и у некоторых «миелиновых» мутантов указывают на то, что миелиновая оболочка определяет локализацию Caspr и contactin в аксолемме. Более того, добавление растворимого Rptpβ, связывающего contactin с myelinating co-cultures, нарушает паранодальное накопление Caspr, указывая на то, что локализация Caspr/contactin complex в этом участке может быть опосредована его взаимодействием с глиальным лигандом. Наиболее подходящим кандидатом глиального лиганда для Caspr/contactin complex является Nf155 – глиальная изоформа CAM neurofascin, который локализован вместе с Caspr и contactin в месте аксоглиального соединения (axoglial junction) и не локализуется в этом участке в отсутствии Caspr. В подтверждение этой гипотезы недавно появилось сообщение, что soluble Nf155-Fc chimaera связывается с клетками, экспрессирующими Caspr и contactin, а преципитация этих белков из лизатов мозга крыс указывает на то, что Nf155 может служить в качестве рецептора для Caspr/contactin complex. Однако последние сообщения подвергли сомнению данную модель, так как было показано, что когда Nf155 связывается непосредственно с contactin, то Caspr подавляет такое взаимодействие. Это указывает на возможное существование других рецепторов для Caspr/contactin complex в миелинизирующей глии. Это наблюдение согласуется с предыдущими сообщениями, показавшими, что Nf155 в paranodes появляется намного позже, чем Caspr.
Function of the paranodal junction. Предполагают, что paranodal junction присоединяет миелиновую оболочку к аксону, чтобы отделить электрическую активность в перехватах Ранвье от межперехватного участка (internodal region), находящегося под компактной миелиновой оболочкой, и служит в качестве преграды (fence), которая ограничивает латеральную диффузию белков аксолеммы. Недавние исследования с использованием 4-х паранодальных мутантных мышей – мышей с отсутствием Caspr, contactin, Cgt и Cst, (все с отсутствием характерной перегородки (septa) в аксоглиальном соединении) – позволили прекратить проверку этих оригинальных идей. В ЦНС этих мутантов паранодальные петли (paranodal loops) дезорганизованы, имеется множество перекрывающихся и инвертированных петель, обращенных в сторону от аксона. В ПНС этих мутантов морфологические изменения менее выражены, возможно, благодаря присутствию базального слоя (basal lamina). Параузлы (paranodes) хорошо организованы, однако увеличено пространство между глиальной оболочкой и аксоном. Но даже в отсутствие перегородки (septa) паранональные петли (loops) все еще тесно прикреплены к аксону во многих участках ЦНС и ПНС, указывая на присутствие до сих пор не идентифицированных паранодальных компонентов, которые опосредуют аксо-глиальный контакт в этом участке. Вместе с электронно-микроскопическими данными, показавшими, что поперечные соединения (transverse bands) образуются позже во время миелинизации, эти исследования указывают на возможную роль перегородки (septa) в сохранении паранодальных петель (колец) для аксона в аксоглиальном соединении. В соответствии с этими наблюдениями, постепенное, зависимое от возраста открепление паранодальных колец от аксона наблюдали в ЦНС Caspr-null мышей (Rios, J. C. et al. Paranodal interactions regulate expression of sodium channel subtypes and provide a diffusion barrier for the node of Ranvier. J. Neurosci. 23, 7001-7011. 2003).
Отсутствие паранодальной перегородки (paranodal septa) у четырех паранодальных мутантов ведет к реорганизации мембраны аксона (РИС.3). У этих мутантов shaker-type К+ каналы, в норме присутствующие в juxtaparanodal области неправильно локализованы по отношению к паранодальной мембране аксона. Поэтому, возможно, что паранодальное перегородчатое соединение (paranodal septate junction) функционирует как барьер, который ограничивает передвижение К+ каналов из-под компактного миелина, отделяя их от Na+ каналов в узловых перехватах. В противоположность juxtaparanodal К+ каналам, нарушение паранодальной перегородки минимально влияет на распределение узловых Na+ каналов. Существует небольшое увеличение длины перехватов, сопровождаемое уменьшением мембранных частиц (particles) в узловой аксолемме, определяемое при FREEZE –FRACTURE электронной микроскопии. Это указывает на то, что paranodal septate junction не является необходимым для появления перехватов. Однако глиальное прикрепление в области paranodes в ЦНС необходимо для сохранения Na+ кластеризации в аксолемме.перехвата Ранвье
Juxtaparanode локализуется в короткой области сразу за paranodal junction (РИС.1b). При freeze-fracture электронной микроскопии эта область представляет собой нерегулярно расположенные частицы, которые в большей степени сконцентрированы вблизи paranodes и более диффузны по мере приближения к межперехватам (internodes). Эти частицы соответствуют скорее всего гетеромультимерам delayed rectifier K+ каналов семейства Shaker – Kv1.1, Kv1.2 и Kvβ2. В juxtaparanodal аксолемме эти каналы колокализуются и образуют комплекс с Caspr2, вторым членом Caspr семейства. Кроме того, в этом участке присутствует Kv1.6, преимущественно в небольших аксонах. Два других белка, обнаруженных в juxtaparanodes, представляют собой транзиторный аксональный glycoprotein-1 (Tag1) – GPI-связанный CAM, родственный contactin, и connexin 29 (Cx29), обнаруженый в глиальной мембране. Связь Caspr2 с К+ каналами опосредуется их карбокси-терминальной областью, вероятнее всего через неидентифицируемый PDZ DOMAIN-содержащий белок. И хотя один такой белок Psd95 локализован в juxtaparanodes и ассоциируется с К+ каналами, он, как оказалось, не участвует во взаимодействии этих каналов с Caspr2 или их накоплении в этой области. Два последних исследования показали, что Caspr2 и Tag1 формируют juxtaparanodal complex, состоящий из глиальной Tag1 молекулы и аксонального Caspr2/Tag1 гетеродимера (РИС.2с) [Poliak, S. et al. Juxtaparanodal clustering of Shaker-like K+ channels in myelinated axons depends on Caspr2 and TAG-1. J. Cell Biol. 162, 1149-1160 (2003)]. Этот комплекс необходим для аккумуляции К+ каналов в juxtaparanodes, поскольку прицельное нарушение Caspr2 или Tag1 приводит к значительному снижению juxtaparanodal аккумуляции этих каналов в аксонах как ЦНС, так и ПНС (РИС.3). Эти результаты указывают, что Caspr2 и Tag1 могут формировать скаффолды, способные позиционировать ионные каналы в специфических участках плазменной мембраны, и это напоминает механизм, работающий во время формирования синапсов.
Role of K+ channels under the myelin sheath. Предполагали, что juxtaparanodal К+ каналы работают как активные «заслонки» (damper) для возвратного возбуждения (re-entrant excitation) и способствуют сохранению межперехватного потенциала покоя (internodal resting potential). И хотя теоретически этих каналов, разбросанных в межперехватах, достаточно для поддержания потенциала покоя, для предупреждения re-entrant excitation требуется повышенная пространственная кластеризация К+ каналов около перехвата. Несмотря на значительное устранение в juxtaparanodal кластере Kv1.1/Kv1.2 у Caspr2- и Tag1-нокаутных мышей, никаких изменений в возбудимости миелинизированных нервов у них не выявлено. То, что общее количество этих каналов остается постоянным у обоих мутантов может указывать на то, что главная роль этих покрытых миелином К+ каналов заключается в поддержании межперехватного потенциала покоя. Кроме того, компьютерное моделирование, при котором учитывались вариации распределения К+ каналов и проводимости аксоглиального соединения (axoglial junctional conductance) показало, что кластеризация К+ каналов в juxtaparanode может обеспечивать защитную функцию в аксонах, которые могут подвергаться слабой демиелинизации (РИС.4). В соответствии с этой моделью Caspr2 иTag1 могут обеспечивать стабильность в аксонах, аксоглиальные соединения которых подвержены определенному риску.
Другой функцией juxtaparanodal К+ каналов может быть их посредническая роль в аксоглиальном процессе передачи информации. В ПНС эти каналы локализованы across from Cx29 гемиканалов (hemichannels), которые присутствуют в adaxonal мембране миелинизирующих Шванновских клеток и наиболее вероятно соответствуют розеточным частицам, видимым при freeze-fracture электронной микроскопии в этом участке. Такие гемиканалы могут обеспечивать прямой путь для ионов К+ из аксона в глию. Это, в свою очередь, могло бы генерировать зависимый от активности сигнал в Шванновскую клетку, что напоминает электрические синапсы сформированные gap junctions. В поддержку этой гипотезы предположили, что Ca2+ transients, записываемые в Шванновских клетках до электрической стимуляции аксона, генерировались К+ efflux (утечкой) из аксона, что деполяризует глиальную мембрану. Обмен информацией через такой «аксоглиальный синапс» в juxtaparanodes может обеспечивать дополнительный механизм для axon–gliacommunication. Интересно то, что паранодальное аксоглиальное соединение (paranodal axoglial junction) также может быть ремоделировано активностью нейронов. Этот эффект может быть отчасти опосредован контролем экспрессии contactin на поверхности аксона.
Хотя никакой специфики соединений не наблюдали между глией и аксоном в области межперехватов, при freeze-fracture электронной микроскопии было обнаружено, что межперехватная аксолемма в ПНС содержит продольные нити (longitudinal strands) внутримембранных частиц, напоминающих нити, найденные в paranodes и в juxtaparanodal областях. Как показано на РИС. 1с и 1d, Caspr и contactin локализуются по всей межперехватной области в полосе, фланкированной К+ каналами и Caspr2, которая соединяет внутренний мезаксон (inner mesaxon) миелиновой оболочки и формирует периферическое кольцо ниже внутренней стороны SCHMIDT-LANTERMAN INCISURES. Эта линия, названная juxtamesaxonal и juxta-incisural (Scherer, S. S. & Arroyo, E. J. Recent progress on the molecular organization of myelinated axons. J. Peripher. Nerv. Syst. 7, 1-12. 2002), является прямым продолжением paranodes/juxtaparanodes. Следовательно, Nf155, Cx29 и Tag1 ограничены в дополнительном распространении на adaxonal мембране миелинизирующих Шванновских клеток. Эти находки указывают, что локализация аксонных белков в межперехватах диктуется миелиновой оболочкой, вероятно, посредством механизма, сходного с тем, который работает в paranode/juxtaparanode. Однако недавний анализ Caspr2-null мышей показал, что разные механизмы могут контролировать локализацию К+ каналов в juxtaparanodes и в juxtamesaxon. Молекулярная организация межперехватной области не изучена в миелинизированных нервах ЦНС.
Formation of the nodal environ
The role of myelinating glia. Во время развития миелинизации нервов в ПНС разные узловые домены формируются постепенно. Na+ каналы сначала кластеризуются в перехватах, затем появляется паранодальное соединение и позже происходит группировка К+ каналов в области juxtaparanode. И в ЦНС, и в ПНС Na+ каналы кластеризуются сначала в участках, прилегающих к границам отростков олигодендроцитов и миелинизирующих Шванновских клеток. Дальнейший продольный рост этих отростков вызывает смещение кластеров до тех пор, пока два соседних кластера не сольются, формируя новый перехват Ранвье. Эти результаты свидетельствуют о том, что Na+ кластеры позиционируются посредством прямого глиального контакта. Следовательно, распространение Na+ каналов является диффузным в ретинальных ганглиозных клетках (retinal ganglion cells), но они правильно кластеризуются в перехватах, после того как аксоны пересекут lamina cribrosa и станут миелинизированными. Эти каналы не группируются после удаления олигодендроцитов или Шванновских клеток и рассеиваются во время демиелинизации. Более того, перехватные Na+ каналы связаны с краями миелинизирующих Шванновских клеток в нервах с более короткими межперехватами, что является результатом ремиелинизации (remyelination) или генетической мутации, как, например, у мутантных мышей CLAW PAW. Однако исследование с использованием ганглиозных клеток ретины показало, что Na+ кластеризация может быть индуцирована in vitro растворимыми факторами, секретируемыми культивируемыми олигодендроцитами. И хотя Шванновские клетки не проявляют такой кластеризирующей активности, некоторая кластеризация Na+ каналов определялась в отсутствии миелинизирующих Шванновских клеток у дистрофичных мышей. Недавний анализ dysmyelinating или paranodal мутантов, а также моделей демиелинизации показал, что присутствие интактных миелинизирующих олигодендроцитов необходимо для включения во время развития изоформы Na+ канала в перехватах. И, напротив, Nav1.6 обнаружена в перехватах у двух миелиновых мутаций, которые ассоциируются с гибелью олигодендроцитов и отсутствием нормального паранодального соединения. – у MYELIN DEFICIENT (MD) крыс и у JIMPY мутантных мышей. Эти наблюдения указывают, что «включение» может происходить в отсутствие нормального паранодального контакта или миелина. Следует отметить, что недавний анализ SHIVERER мутантов показал, что в то время как аксоглиальный контакт необходим для экспрессии Nav1.6 в перехватах, он не нужен для таргетирования этой подгруппы в начальном сегменте аксона, что свидетельствует о существовании множественных адресных механизмов при миелинизации аксонов.
Molecular assembly. Во время миелинизации нервов в ПНС, Nrcam и Nf186 первыми определяются в перехватах, после чего появляются ankyrin G и Na+ каналы. Однако в ЦНС ankyrin G определяется в перехватах до кластеризации Nf186 и Na+ каналов. Эти результаты указывают, что Nrcam, Nf186 или неидентифицируемый ankyrin G-binding белок связывают ankyrin G, который, в свою очередь, «рекрутирует» Na+ каналы. В поддержку этой модели свидетельствует тот факт, что добавление растворимого Nrcam в культуру myelinating dorsal root ganglia подавляет кластеризацию Na+ каналов. Более того, появление ankyrin G и Na+ каналов в перехватах отсрочено у Nrcam-null мышей, что свидетельствует об участии этой адгезивной молекулы в кластеризации. Формирование перехватов у этих животных можно объяснить присутствием Nf186, который содержит сходный ankyrin G-binding сайт и, следовательно, может компенсировать отсутствие Nrcam. Значимость взаимодействия между ankyrin G и такими компонентами перехвата была показана у мышей с отсутствием мозжечковой изоформы ankyrin G, у которых Na+ каналы, Ig-CAMs и spectrin βIV не группируются в начальном сегменте аксонов клеток Пуркинье. Похожую картину наблюдали у спонтанной мутации по spectrin βIV – QUIVERING мышей (или у мышей с прицельным нарушением этого гена), у которых найдены аномалии перехватов и измененное распределение каналов. Однако ankyrin G также присутствует в параузлах (paranodes) во время ранней миелинизации аксонов, что говорит не о его прямом участии в первичном таргетировании Na+ каналов, а скорее о его важности для их стабилизации. Кроме того, ankyrin G в норме локализован в перехватах у dystroglycan-null мышей, у которых показана значительная редукция кластеров перехватных Na+ каналов. После первичной кластеризации компонентов перехватов в волокнах ПНС, Nf155 и Caspr/Contactin комплекс аккумулируются в paranodal junction. После этого появляются Caspr2 и К+ каналы в juxtaparanodal области. Caspr2, К+ каналы и TAG-1 первыми определяются в параузлах (paranodes) и позже они перемещаются в juxtaparanodes как формы паранодального соединения (paranodal junction forms). В отсутствии такого соединения К+ каналы не перемещаются в juxtaparanodes и остаются рядом с перехватами (РИС.4; ТАБЛ.1 и 2). Дальнейшее поддержание К+ каналов в juxtaparanodal области требует Caspr2 и Tag1, поскольку в их отсутствии эти каналы перераспределяются по межперехватам.
Molecular sieves, pickets and fences. Сегрегация белков на отдельные домены в нейронах достигается через специфические сортировочные механизмы, за которыми следует закрепление и кластеризация этих белков в плазменной мембране. Формирование окружения перехвата Ранвье может включать несколько различных молекулярных механизмов (РИС.5). Удаление Na+ каналов с границ миелинизирующей глии во время развития может быть осуществлено с помощью селективной молекулярной фильтрации или «сита», обнаруженной в paranodes (РИС.5с). Предполагали, что такое «сито» не пропускает крупные белковые комплексы, включая Na+ каналы и Ig-CAMs, соединенные с ankyrin G, и пропускает мелкие мембранные частицы – соответствующие К+ каналам. Такой процесс требует аксоглиального контакта, но он не опосредуется Caspr/contactin комплексом, т.к. его отсутствие не препятствует кластеризации Na+ каналов в перехватах. Поэтому, появление зрелых, септ-содержащих (septa-containing) паранодальных соединений может быть необязательным для эффективной кластеризации Na+ каналов. Это мнение подтверждается данными freeze-fracture электронной микроскопии, при которой была показана ранняя дифференцировка перехватов еще до появления paranodal septa (паранодальной перегородки). Следовательно, накопление Caspr в параузлах и кластеризация Na+ каналов в перехватах происходит перед появлением septa (перегородки). Следует заметить, что постепенное открепление паранодальных петель (paranodal loops) в ЦНС у паранодальных мутантов сопровождается расширением пробела перехвата и рассеиванием Na+ каналов. Это указывает на то, что хотя septa и не является необходимым компонентом для первичной сборки Na+ каналов в перехватах, стабилизированные глиальные контакты (зависимые от septa) в paranodes могут быть необходимым компонентом для поддержания этих кластеров. Интересно, что кластеризация Na+ каналов в зрительном нерве Caspr-null мышей, у которых отсутствует paranodal septa, ассоциируется с кластерами соседних К+ каналов и это увеличивает вероятность того, что Caspr2 и Tag1 компенсируют в этих участках отсутствие Caspr и contactin.
В отличие от кластеризации Na+ каналов в перехватах, формирование septa- содержащих аксоглиальных соединений важно для изоляции К+ каналов в juxtaparanodes. Эти наблюдения указывают, что, будучи однажды сформированными, аксоглиальные septate соединения функционируют как барьер, ограничивающий движение этих каналов и других молекул из-под миелиновой оболочки к перехватам. Предполагается также, что молекулярное «сито», функционирующее в paranodes во время формирования перехватов, могло бы изменить эти свойства после того как paranodal loops уже обеспечат безопасность аксона посредством septate junction. Появление такого барьера может быть опосредовано прикреплением Caspr/contactin комплекса к цитоскелету аксона, связывающегося с глиальным лигандом, и сборкой специфических липидных микродоменов. И хотя вклад липидного состава мембраны в появление доменов аксона пока не изучен, интересно то, что contactin, Caspr и Tag1 ассоциируются с липидными плотиками (rafts).
Кроме paranodal junction возможно также существование мембранного барьера в перехватах. И хотя Caspr2/K+ канальный комплекс и Tag1 неправильно локализованы в паранодальной области в отсутствие paranodal junction, эти белки не проникают в перехваты, указывая на присутствие дополнительного барьера в этом участке. Такой барьер в области перехвата может быть сходен с диффузным барьером (или мембранным барьером), обнаруженным в начальном сегменте аксона, который можно рассматривать как первый перехват в большинстве миелинизированных аксонов. В начальном сегменте аксона этот барьер сформирован высоко концентрированными локальными трансмембранными белками, которые прикреплены к актиновому цитоскелету и которые служат в качестве «пикетов», способных блокировать диффузию мембранных белков и фосфолипидов. Интересно то, что интактный актиновый цитоскелет в аксонах ганглиев сетчатки также необходим для кластеризации Na+ каналов при воздействии растворимого фактора, секретируемого олигодендроцитами.
Рассматриваются два других молекулярных механизма, участвующих в формировании перехватов. В ПНС кластеризация перехватных Na+ каналов во время развития может быть опосредована контактирующими глиальными отростками, которые «тянут» Na+ каналы и Ig-CAMs по направлению к их окончательной позиции на аксолемме (РИС.5b). Это может быть осуществлено с помощью связывания Na+ каналов с микроворсинками Шванновских клеток, либо непосредственно через их β-субъединицы, либо косвенно через Nrcam и Nf186. Во время развития ERM-позитивные ворсинки Шванновских клеток осуществляют ранний контакт с перехватами во время их образования. Эти участки контактов (названные «caps») содержат phosphorylated adaptor EBP50 и face across axonal ankyrin G. Разрушение микроворсинок у мышей без dystroglycan в Шванновских клетках ведет к значительному снижению кластеризации Na+ каналов в узловых перехватах. Остается выяснить, связывает ли dystroglycan какие либо узелковые белки, опосредуя, таким образом, такое аксоглиальное взаимодействие.
Микроворсинки содержат также другие кандидатные белки, включая L1 и neurofascin, способные связывать Ig-CAMs, присутствующие в аксолемме. Наконец, возможно, что кластеризация Na+ каналов в перехватах Ранвье осуществляется с помощью downregulation этих каналов из нижних (from beneath) межперехватов и посредством селективной инсерции вновь синтезированных или рециклированных молекул в формирующийся узловой пробел (РИС.5d). Маловероятно, но во время развития может работать механизм «доставки». На это указывают такие наблюдения как перемещение изоформ Na+ каналов после формирования перехватов, присутствие высокой концентрации везикул в перехватах и тесная связь Na+ каналов с микротрубочками. Сходный механизм также может работать при формировании juxtaparanodes в ЦНС, где К+ каналы первыми определяются во время развития.



