Что такое пикообразные потенциалов действия покажите это схематически
Следовые потенциалы. Последний участок фазы реполяризации для некоторых видов потенциалов действия бывает замедленным; хорошим примером служит потенциал действия мышцы на рис. 2.4. Приблизительно через 1 мс после начала потенциала действия наблюдается отчетливый перегиб кривой реполяризации; следующее за ним медленное изменение потенциала называется деполяриза-ционным следовым потенциалом. В других тканях, например в нейронах спинного мозга, кривая деполяризации быстро пересекает уровень потенциала покоя, так что на некоторое время потенциал становится более отрицательным, чем потенциал покоя. Это явление получило название гиперполяризационный следовой потенциал (рис. 2.5).
Природа потенциала действия
Порог и возбудимость. Каким образом потенциал покоя, обычно поддерживаемый на постоянном уровне посредством только что обсуждавшихся механизмов, нарушается до такой степени, что возникает потенциал действия? Потенциалы действия генерируются при деполяризации мембраны от потенциала покоя до примерно —50 мВ. Механизмы развития этой начальной деполяризации будут рассмотрены позднее. Уровень потенциала, при котором деполяризация приводит к потенциалу действия, называется порогом (рис. 2.5). При таком пороговом потенциале заряд мембраны становится нестабильным; он нарушается посредством внутреннего механизма, который ведет к реверсии полярности-быстрому нарастанию потенциала действия до пика. Это состояние автоматического прогрессирующего нарушения мембранного заряда называется возбуждением. Обычно возбуждение продолжается менее 1 мс. Оно подобно взрыву-протекает мощно, но быстро завершается. После фазы деполяризации наступает процесс восстановления заряда мембраны, присущего состоянию покоя.
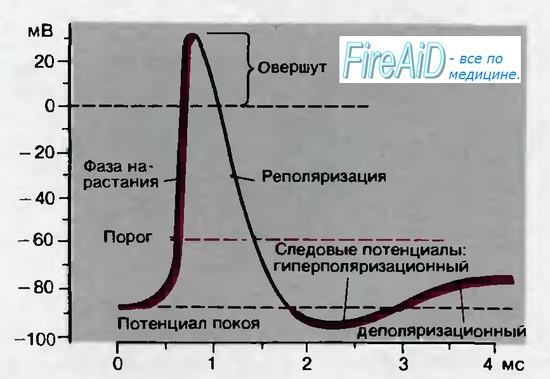 Рис. 2.5. Временной ход потенциала действия в нейроне; показаны последовательные фазы потенциала действия, описанные в тексте
Рис. 2.5. Временной ход потенциала действия в нейроне; показаны последовательные фазы потенциала действия, описанные в тексте
Клетки, в которых можно вызвать потенциалы действия, называются возбудимыми. Возбудимость является типичным свойством нервных и мышечных клеток. Клетки каждого типа характеризуются собственным постоянным временным ходом потенциала действия. Он практически не зависит от источника или частоты возбуждения клетки. Поскольку форма потенциала действия постоянна, говорят, что возбуждение протекает по закону «все или ничего».
Что такое пикообразные потенциалов действия покажите это схематически
В нервных волокнах сигналы передаются с помощью потенциалов действия, которые представляют собой быстрые изменения мембранного потенциала, быстро распространяющиеся вдоль мембраны нервного волокна. Каждый потенциал действия начинается со стремительного сдвига потенциала покоя от нормального отрицательного значения до положительной величины, затем он почти так же быстро возвращается к отрицательному потенциалу. При проведении нервного сигнала потенциал действия движется вдоль нервного волокна вплоть до его окончания.
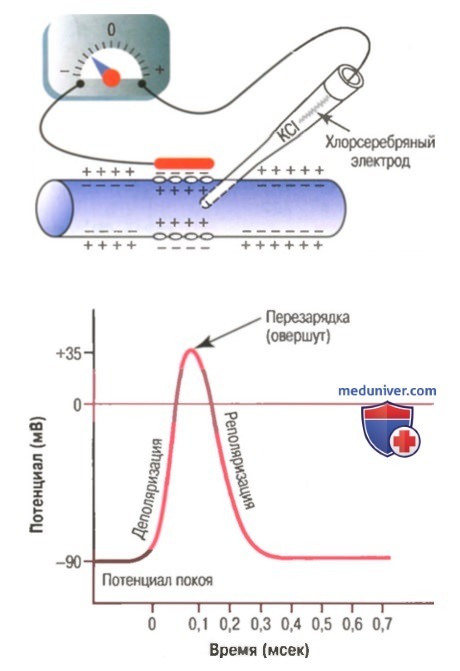 Типичный потенциал действия, зарегистрированный с помощью метода, представленного в верхней части рисунка.
Типичный потенциал действия, зарегистрированный с помощью метода, представленного в верхней части рисунка.
Фаза реполяризации. В течение нескольких долей миллисекунды после резкого повышения проницаемости мембраны для ионов натрия, натриевые каналы начинают закрываться, а калиевые — открываться. В результате быстрая диффузия ионов калия наружу восстанавливает нормальный отрицательный мембранный потенциал покоя. Этот процесс называют реполя-ризацией мембраны.
Для более полного понимания факторов, являющихся причиной деполяризации и реполяризации, необходимо изучить особенности двух других типов транспортных каналов в мембране нервного волокна: электроуправляемых натриевых и калиевых каналов.
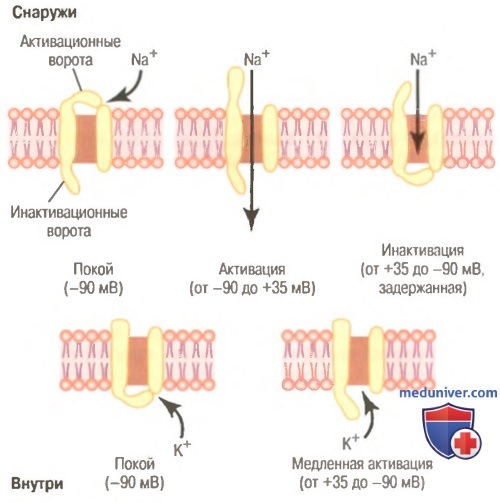 Типичные изменения состояния электроуправляемых натриевых (вверху) и калиевых (внизу) каналов при изменении мембранного потенциала от нормального отрицательного значения до положительных величин.
Типичные изменения состояния электроуправляемых натриевых (вверху) и калиевых (внизу) каналов при изменении мембранного потенциала от нормального отрицательного значения до положительных величин.
Показана последовательная активация и инактивация натриевых каналов и задержанная активация калиевых каналов.
Инактивация натриевого канала. В верхней правой части рисунке показано третье состояние натриевого канала. Увеличение потенциала, открывающее активационные ворота, закрывает инактивационные ворота. Однако инактивационные ворота закрываются в течение нескольких десятых долей миллисекунды после открытия активационных ворот. Это значит, что конформационное изменение, приводящее к закрытию инактивационных ворот, — процесс более медленный, чем конформационное изменение, открывающее активационные ворота. В результате через несколько десятых долей миллисекунды после открытия натриевого канала инактивационные ворота закрываются, и ионы натрия не могут более проникать внутрь волокна. С этого момента мембранный потенциал начинает возвращаться к уровню покоя, т.е. начинается процесс реполяризации.
Существует другая важная характеристикая процесса инактивации натриевого канала: инактивационные ворота не открываются повторно до тех пор, пока мембранный потенциал не вернется к значению, равному или близкому к уровню исходного потенциала покоя. В связи с этим повторное открытие натриевых каналов обычно невозможно без предварительной реполяризации нервного волокна.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в оглавление раздела «Физиология человека.»
Потенциал действия нервной клетки
Содержание материала
 Потенциал действия (ПД) — быстрое колебание МП — самораспространяющийся процесс, связанный с изменениями ионной проводимости мембраны, вызванными функционированием ионных каналов. ПД распространяется без затухания, то есть практически без уменьшения амплитуды.
Потенциал действия (ПД) — быстрое колебание МП — самораспространяющийся процесс, связанный с изменениями ионной проводимости мембраны, вызванными функционированием ионных каналов. ПД распространяется без затухания, то есть практически без уменьшения амплитуды.
Временной ход потенциала действия
Продолжительность потенциала действия нервной клетки измеряется единицами миллисекунд (мс).
Потенциалы действия, зарегистрированные двумя электродами, один из которых находится внутри клетки, а другой — в окружающем растворе, представлены на рис. 5-3 и 5-7.
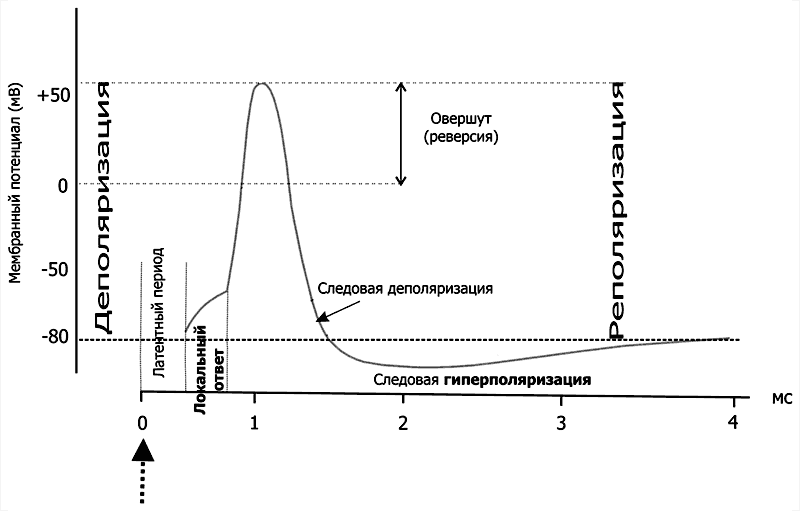
Рис. 5–3. Изменения мембранного потенциала и потенциал действия. Вертикальная стрелка в нижней части рисунка — момент появления раздражающего стимула, на отметке –80 мВ — исходный уровень МП.
Между моментом нанесения раздражения и первым проявлением ПД имеется задержка — латентный период. Латентный период соответствует времени, когда ПД движется по мембране нервной клетки от места раздражения до отводящего электрода. Под действием раздражающего стимула происходит нарастающая деполяризация мембраны — локальный ответ. При достижении критического уровня деполяризации, который в среднем составляет —55 мВ, начинается фаза деполяризации. В эту фазу уровень МП падает до нуля и даже приобретает положительное значение (овершут), а затем возвращается к исходному уровню (фазареполяризации). Фазы деполяризации, овершута и реполяризации образуют спайк (пик) ПД. Длительность спайка составляет 1—2 мс. После спайка наблюдается замедление скорости спада потенциала — (раза следовой деполяризации. После достижения исходного уровня покоя нередко наблюдается фаза следовой гиперполяризации. Эти следовые потенциалы могут длиться десятки и сотни миллисекунд.
Ионные механизмы потенциала действия
В основе изменений мембранного потенциала (МП), происходящих в течение потенциала действия (ПД), лежат ионные механизмы. На рис. 5—7 представлены суммарные ионные токи, протекающие через мембрану нервной клетки в ходе потенциала действия.

Рис. 5-7. Потенциал действия и ионные токи нервной клетки [5|. По оси ординат отложены значения МП (мВ), по оси абсцисс — время (мс)
Локальный потенциал. Раздражение клетки приводит к открытию части Na+-каналов и появлению локального (нераспространяющегося) потенциала.
Фаза деполяризации. При достижении критического уровня деполяризации мембраны (соответствует порогу активации потен-циалозависимых Na+-каналов) начинается лавинообразный процесс открытия большого количества Na+-каналов. В фазу деполяризации происходит массивный вход в клетку ионов Na+ по концентрационному и электрохимическому градиентам.
Овершут. Деполяризация мембраны приводит к реверсии МП (МП становится положительным). В фазу овершута Na+-ток начинает стремительно спадать, что связано с инактивацией потенциало-зависимых Na+-каналов (время открытого состояния –– доли миллисекунды) и исчезновением электрохимического градиента Na+.
Фаза реполяризации. Помимо инактивации Na+-каналов, развитию реполяризации способствует открытие потенциалозависимых K+-каналов. Этот процесс происходит медленнее, чем открытие Na+-каналов, но K+-каналы остаются открытыми более продолжительное время. Выход К+ наружу способствует полному завершению фазы реполяризации.
Следовые потенциалы связаны с длительными изменениями кинетических свойств К+-каналов. Восстановление исходного уровня МП приводит Na+- и K+-каналы в состояние покоя.
Изменения возбудимости во время потенциала действия
В ходе развития ПД происходят изменения возбудимости мембраны нервной клетки.
Абсолютно рефрактерный период. Во время фазы деполяризации и большей части фазы реполяризации ПД клетка находится в абсолютно рефрактерном периоде, в течение которого даже сверхпороговое раздражение не способно вызвать ПД. Этот феномен связан с инактивацией большинства Na+-каналов.
Относительно рефрактерный период. В конце фазы реполяризации, а также во время следовой гиперполяризации клетка способна генерировать ПД только в ответ на сверхпороговые раздражители. Это связано со значительным реполяризующим действием выходящих калиевых токов.
Наличие рефрактерности ограничивает частоту генерации ПД. Физиологическое значение рефрактерности заключается в создании условий для своевременного и полного осуществления восстановительных процессов в нервной клетке. Феномен рефрактерности лежит в основе понятия о функциональной подвижности, или лабильности (Н.Е. Введенский [1] ).
Лабильность — максимально возможная частота генерации ПД для данного типа возбудимой клетки. Лабильность большинства нейронов составляет приблизительно 400 ПД/с, а у интернейронов спинного мозга доходит до 1000 ПД/с.
В нормальных условиях рефрактерность предохраняет нервные клетки от излишне частой генерации ПД. Мутации, гипоксия, механические травмы и другие патологические воздействия приводят к значительным изменениям возбудимости нейронов. Такие нейроны или группы нейронов являются потенциальными источниками возникновения пароксизмальных состояний ЦНС, эпилептических припадков и других неврологических расстройств.
Проведение возбуждения по нервным волокнам
Нервные волокна — аксоны нервных клеток, окружённые оболочкой из олигодендроглиоцитов в ЦНС и шванновских [2] клеток в периферических нервах. Нервные волокна подразделяют на 2 типа — безмиелиновые и миелиновые. Основная функция нервных волокон — проведение ПД. Скорость проведения в миелиновых и безмиелиновых волокнах различна (рис. 5–8) и существенно зависит от диаметра нервных волокон.

Рис. 5–8. Скорость проведения возбуждения в миелиновых и безмиелиновых нервных волокнах разного диаметра [4]. Скорость проведения пропорциональна диаметру нервного волокна и в миелиновых волокнах выше, чем в безмиелиновых.
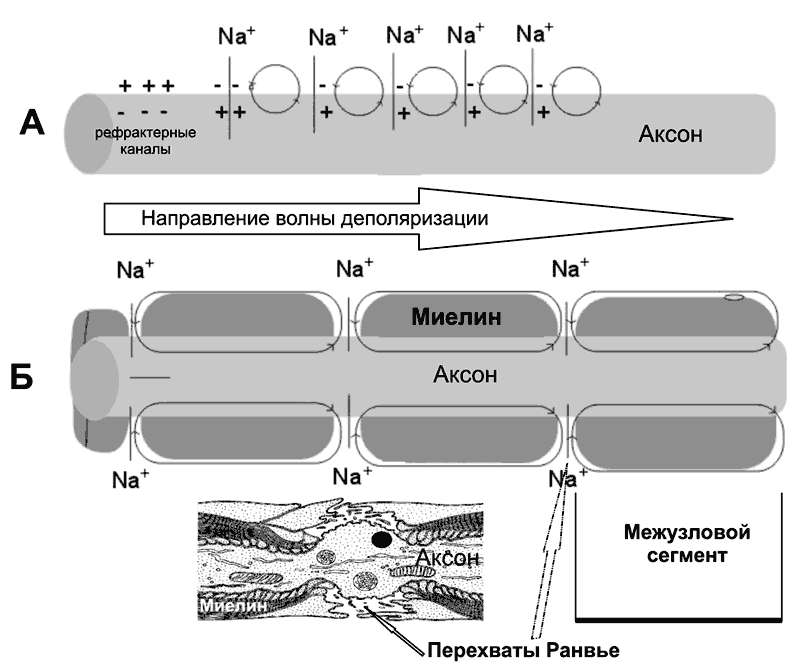
Безмиелиновые нервные волокна (рис. 5–9А). В покое мембрана аксона (осевого цилиндра) поляризована — положительно заряжена снаружи и отрицательно внутри. При ПД полярность изменяется, и наружная поверхность мембраны приобретает отрицательный заряд. Из-за разности потенциалов между возбуждённым и невозбуждёнными сегментами возникают локальные токи, деполяризующие соседний участок мембраны. Теперь этот участок становится возбуждённым и деполяризует следующий участок мембраны.
Рис. 5–9. Проведение возбуждения в нервных волокнах [7]. А — безмиелиновое волокно (электротоническое проведение), Б — миелиновое волокно (скачкообразное проведение). Миелин, полностью окружая аксон в межузловых промежутках, выступает в роли электрического изолятора, а межклеточная жидкость в перехватах Ранвье [3] — проводник.
Появление так называемых рефрактерных каналов (рефрактерное состояние мембраны после прохождения ПД) предупреждает распространение возбуждения в обратном направлении.
Скорость проведения возбуждения по безмиелиновому нервному волокну в основном составляет 0,5–2 м/с и зависит от диаметра волокна: чем больше диаметр, тем выше скорость проведения ПД (см. рис. 5–8).
Миелиновое нервное волокно (рис. 5–9Б) состоит из осевого цилиндра (аксона), вокруг которого шванновские клетки образуют миелин за счёт концентрического наслаивания собственной плазматической мембраны. Миелин прерывается через регулярные промежутки (от 0,2 до 2 мм) концентрической щелью шириной около 1 мкм, это узлы, или перехваты Ранвье. Таким образом, межузловые сегменты аксона, расположенные между соседними перехватами Ранвье, содержат миелин — электрический изолятор, не позволяющий проходить через него локальным токам, поэтому ПД возникают только в перехватах Ранвье. Другими словами, ПД перемещается вдоль нервного волокна скачками, от одного перехвата Ранвье к другому перехвату (скачкообразное проведение).
Плотность потенциалозависимых Na+?каналов аксолеммы в перехватах Ранвье — до 2000 на 1 мкм2 (в перикарионе — 50–70, в начальном сегменте аксона — 2000, в межузловых сегментах Na+?каналы практически отсутствуют). В силу высокой плотности Na+?каналов перехваты Ранвье характеризуются высокой возбудимостью, а локальные токи достаточно велики для возбуждения соседнего перехвата.
Локальные токи текут от перехвата к перехвату (через внеклеточную жидкость кнаружи от миелина и через аксоплазму внутри аксона) с минимальными потерями.
Скорость проведения ПД в миелиновых волокнах в десятки раз выше, чем в наиболее «быстрых» безмиелиновых аксонах.
Энергозатраты нервного волокна на проведение ПД относительно невелики, поскольку возбуждаются только перехваты Ранвье, площадь которых составляет менее 1% общей поверхности мембраны аксона. Поэтому даже после длительных ритмических пачек ПД трансмембранный градиент концентраций ионов практически не изменяется.
В физиологических условиях ПД движутся в одном направлении от места раздражения (ортодромное проведение). ПД, проходящий по нервному волокну, возбуждает следующий, но не предыдущий участок мембраны. Это связано с рефрактерностью предыдущего участка после возбуждения. Проведение в противоположном направлении (антидромное проведение) возможно при травматическом поражении нервных волокон и в редких случаях (аксон–рефлекс).
Нарушение миелинизации нервных волокон приводит к нарушениям проводимости (демиелинизирующие заболевания). При разрушении миелиновой оболочки происходит резкое снижение скорости и надёжности проведения возбуждения по нервам. Наиболее распространённым среди демиелинизирующих заболеваний является множественный склероз, проявляющийся различными параличами и потерей чувствительности.
Законы проведения возбуждения
Бездекрементное проведение возбуждения. Амплитуда ПД в различных участках нерва одинакова, то есть проведение возбуждения по нервному волокну осуществляется без затухания (бездекрементно). Таким образом, кодирование информации осуществляется не за счёт изменения амплитуды ПД, а путём изменения их частоты и распределения во времени.
Изолированное проведение возбуждения. Нервные стволы обычно образованы большим количеством нервных волокон, однако ПД, идущие по каждому из них, не передаются на соседние. Эта особенность нервных волокон обусловлена:
Физиологическая и анатомическая целостность. Необходимым условием проведения возбуждения является не только его анатомическая целостность, но и нормальное функционирование мембраны нервного волокна (физиологическая целостность). В клинике широко применяют различные ЛС, нарушающие физиологическую целостность нервных волокон. Так, эффекты местных анестетиков (новокаин, лидокаин, и др.) основаны на блокаде потенциалозависимых Na+?каналов. Нарушение физиологической целостности чувствительных нервных волокон вызывает анестезию (потерю чувствительности).
Типы нервных волокон и их функции

[1] Введенский Николай Евгеньевич (1852-1922), отечественный физиолог, ученик И.М. Сеченова; впервые при отведении токов действия нерва с помощью телефона прослушал ритм возбуждения в нерве. Сравнивая ритмы возбуждения в нерве и мышце, он открыл явления оптимума и пессимума, на основе которого установил закон относительной функциональной подвижности — лабильности. Ему принадлежит учение о парабиозе, развитое в монографии «Возбуждение, торможение и наркоз» (1901).
[2] Шванн Теодор (Schwann T.), немецкий гистолог и физиолог (1810–1882); вместе с М. Шульце создал клеточную теорию (1839 г.); в 1836 г. открыл пепсин, в 1838 г. опубликовал первую работу по строению миелиновой оболочки.
[3] Ранвье Луи (Ranvier L.), французский патолог (1835–1922). Занимался изучением нервной ткани с применением азотнокислого серебра и хлорного золота. Его именем названы безмиелиновые участки (узловые перехваты) миелинового нервного волокна.
Потенциал действия. Ионный механизм возникновения потенциала действия
Потенциалом действия (ПД)называют быстрое колебание мембранного потенциала, возникающее при возбуждении нервных, мышечных и некоторых других клеток. В его основе лежат изменения ионной проницаемости мембраны. Амплитуда ПД мало зависит от силы вызывающего его раздражителя, важно лишь, чтобы эта сила была не меньше некоторой критической величины, которая называется порогом раздражения.Возникнув в месте раздражения, ПД распространяется вдоль нервного или мышечного волокна, не изменяя своей амплитуды.
В естественных условиях ПД генерируются в нервных волокнах при раздражении рецепторов или возбуждении нервных клеток. Распространение ПД по нервным волокнам обеспечивает передачу информации в нервной системе.Достигнув нервных окончаний, ПД вызывают секрецию химических веществ (медиаторов), обеспечивающих передачу сигнала на мышечные или нервные клетки. В мышечных клетках ПД инициируют цепь процессов, вызывающих сократительный акт. Ионы, проникающие в цитоплазму во время генерации ПД, оказывают регулирующее влияние на метаболизм клетки и, в частности, на процессы синтеза белков, составляющих ионные каналы и ионные насосы.
| В ПД выделяют следующие фазы (рис. 3): ■ деполяризации; ■ реполяризации. |
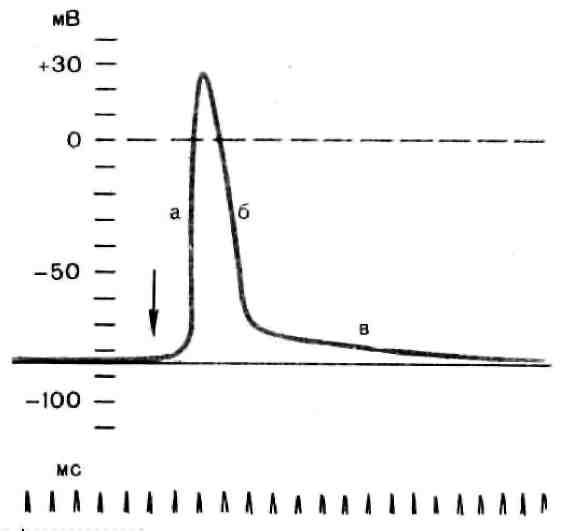
Рис. 3. Потенциал действия скелетного мышечного волокна,зарегистрированный с помощью внутриклеточного микроэлектрода: а – фаза деполяризации, б – фаза реполяризации, в – фаза следовой деполяризации (отрицательный следовой потенциал). Момент нанесения раздражения показан стрелкой.
Установлено, что во время восходящей фазы (фазы деполяризации) происходит не просто исчезновение потенциала покоя (как это первоначально предполагали), а возникает разность потенциалов обратного знака: внутреннее содержимое клетки становится заряженным положительно по отношению к наружной среде, иными словами, происходит реверсия мембранного потенциала.Во время нисходящей фазы (фазы реполяризации) мембранный потенциал возвращается к своему исходному значению. Если рассмотреть пример записи ПД в скелетном мышечном волокне лягушки (см. рис. 3), то видно, что в момент достижения пика мембранный потенциал составляет +30 – +40 мВ. Длительность пика ПД у различных нервных и мышечных волокон варьирует от 0,5 до 3 мс, причем фаза реполяризации продолжительнее фазы деполяризации.
Изменения мембранного потенциала, следующие за пиком потенциала действия, называют следовыми потенциалами.Различают два вида следовых потенциалов – следовую деполяризацию и следовую гиперполяризацию.
Повышение проницаемости мембраны для Na + продолжается лишь очень короткое время. Вслед за этим проницаемость мембраны для Na + вновь понижается, а для K + возрастает. Процесс, ведущий к понижению ранее увеличенной натриевой проницаемости мембраны, назван натриевой инактивацией.В результате инактивации поток Na + внутрь цитоплазмы резко ослабляется. Увеличение же калиевой проницаемости вызывает усиление потока K + из цитоплазмы во внешний раствор. В итоге этих двух процессов и происходит реполяризация мембраны: внутреннее содержимое клетки вновь приобретает отрицательный заряд по отношению к наружной стороне мембраны. Этому изменению потенциала соответствует нисходящая фаза ПД (фаза реполяризации). Опыты на гигантских нервных волокнах кальмара позволили получить подтверждение правильности натриевой теории возникновения ПД.
В основе рассмотренных изменений ионной проницаемости мембраны при генерации ПД лежат процессы открывания и закрывания специализированных ионных каналов в мембране, обладающих двумя важнейшими свойствами:
■ избирательностью (селективностью) по отношению к определенным ионам;
■ электровозбудимостью, т.е. способностью открываться и закрываться в ответ на изменения мембранного потенциала.
Так же как ионные насосы, ионные каналы образованы макромолекулами белков, пронизывающими липидный бислой мембраны.
Активный и пассивный ионный транспорт.В процессе восстановления после ПД работа калий-натриевого насоса обеспечивает «откачку» излишних ионов натрия наружу и «накачивание» потерянных ионов калия внутрь, благодаря чему нарушенное при возбуждении неравенство концентраций Na + и K + по обе стороны мембраны восстанавливается. На работу этого механизма тратится около 70 % необходимой клетке энергии.
Таким образом, в живой клетке существует две системы движения ионов через мембрану.
Один из них осуществляется по градиенту концентрации ионов и не требует затраты энергии (пассивный ионный транспорт). Он ответствен за возникновение потенциала покоя и ПД и ведет в конечном итоге к выравниванию концентрации ионов по обе стороны клеточной мембраны.
Проведение возбуждения
Нервный импульс (потенциал действия) обладает способностью распространяться вдоль по нервным и мышечным волокнам.
В нервном волокне потенциал действия является очень сильным раздражителем для соседних участков волокна. Амплитуда потенциала действия обычно в 5 – 6 раз превышает пороговую величину деполяризации. Это обеспечивает высокую скорость и надежность проведения.
Между зоной возбуждения (имеющей на поверхности волокна отрицательный заряд и на внутренней стороне мембраны – положительный) и соседним невозбужденным участком мембраны нервного волокна (с обратным соотношением зарядов) возникают электрические токи – так называемые местные токи.В результате развивается деполяризация соседнего участка, увеличение его ионной проницаемости и появление потенциала действия. В исходной же зоне возбуждения восстанавливается потенциал покоя. Затем возбуждением охватывается следующий участок мембраны и т.д. Таким образом, с помощью местных токов происходит распространение возбуждения на соседние участки нервного волокна, т.е. проведение нервного импульса.По мере проведения амплитуда потенциала действия не уменьшается,т.е. возбуждение не затухает даже при большой длине нерва.
Во время развития потенциала действия мембрана полностью теряет возбудимость. Это состояние называют полной невозбудимостью, или абсолютной рефрактерностью.За ним следует относительная рефрактерность, когда потенциал действия может возникать лишь при очень сильном раздражении. Постепенно возбудимость восстанавливается до исходного уровня.
Законы проведения возбуждения в нервах:
1. Проведение импульсов возможно лишь при условии анатомической и физиологической целостности волокна.
2. Двустороннее проведение: при раздражении нервного волокна возбуждение распространяется по нему и в центробежном, и в центростремительном направлениях.
3. Изолированное проведение: в периферическом нерве импульсы распространяются по каждому волокну изолированно, т.е. не переходя с одного волокна на другое и оказывая действие только на те клетки, с которыми контактируют окончания данного нервного волокна.



