ПЛАЦЕНТАРНЫЕ
Смотреть что такое «ПЛАЦЕНТАРНЫЕ» в других словарях:
Плацентарные — Научная классификация … Википедия
ПЛАЦЕНТАРНЫЕ — группа живородящих млекопитающих. Зародыши развиваются в матке с образованием плаценты. К плацентарным относятся все млекопитающие (исключая первозверей и сумчатых) … Большой Энциклопедический словарь
плацентарные — ых; Зоол. Группа живородящих млекопитающих. * * * плацентарные группа живородящих млекопитающих. Зародыши развиваются в матке с образованием плаценты. К плацентарным относятся все млекопитающие (исключая первозверей и сумчатых). * * *… … Энциклопедический словарь
плацентарные — (син. одноутробные) животные, у которых во время беременности развивается плацента; к П. относятся все млекопитающие, кроме представителей отр. однопроходных (Monotremata) и сумчатых (Marsupialia) … Большой медицинский словарь
Плацентарные — (Eutheria) инфракласс живородящих млекопитающих (См. Млекопитающие), характеризующийся наиболее высокой организацией и эколого морфологическим разнообразием. Характерные особенности: головной мозг имеет сильно развитые большие полушария,… … Большая советская энциклопедия
Плацентарные — мн. Высшая группа млекопитающих, у которых зародыш связан с телом матери плацентой. Толковый словарь Ефремовой. Т. Ф. Ефремова. 2000 … Современный толковый словарь русского языка Ефремовой
ПЛАЦЕНТАРНЫЕ — группа живородящих млекопитающих. Зародыши развиваются в матке с образованием плаценты. К П. относятся все млекопитающие (исключая первозверей и сумчатых) … Естествознание. Энциклопедический словарь
плацентарные — мн., Р. плацента/рных … Орфографический словарь русского языка
плацентарные — ых; зоол. Группа живородящих млекопитающих … Словарь многих выражений
Основные отряды Плацентарных млекопитающих

Отряд Насекомоядные
(крот, землеройка, ёж, выхухоль) объединяет наиболее примитивных плацентарных зверей. Мозг их относительно мал, кора гладкая, без извилин. У насекомоядных большое количество слабо дифференцированных зубов. Морда вытянута в длинный подвижный хоботок. Размеры тела средние и мелкие. Питаются насекомыми и их личинками.
Отряд Грызуны
(мыши, полёвки, крысы, белки, суслики, сурки, бобры, ондатры, нутрии, хомяки, сони, тушканчики) — самый многочисленный среди млекопитающих (около 2 тыс. видов). Для них характерно отсутствие клыков, сильно развитые резцы и плоская жевательная поверхность коренных зубов. Резцы, а у многих и коренные зубы, у них не имеют корней и растут всю жизнь. Между резцами и коренными зубами широкое пространство, лишённое зубов.
Некоторые грызуны имеют промысловое значение, например, белка, ондатра, бобр, нутрия и др. Многие виды грызунов (мыши, полёвки, крысы) являются вредителями сельского хозяйства и переносчиками ряда опасных заболеваний человека и домашних животных (чумы, туляремии, клещевого возвратного тифа, энцефалита и др.).
Отряд Хищные
в основном плотоядные, реже всеядные животные. У Хищных резцы малы, клыки всегда хорошо развиты, коренные бугорчатые зубы с острыми режущими вершинами. Главные семейства Хищных:
Многие виды служат объектами пушного промысла или разводятся на звероводческих фермах (американская норка, соболь, голубой песец, серебристо-чёрная лисица). Численность наиболее опасных хищников (волки) регулируется человеком.
Отряд Парнокопытные
включает 170 видов. К ним относятся копытные млекопитающие с одинаково сильно развитыми третьим и четвёртым пальцами. Первый палец отсутствует, второй и пятый развиты слабо или полностью отсутствуют.
Различают нежвачных и жвачных парнокопытных.
У нежвачных (свиньи, бегемоты) желудок простой, и пищу для повторного пережёвывания они не отрыгивают.
Жвачные парнокопытные (коровы, овцы, козы, олени, верблюды, лоси, антилопы, жирафы и др.) имеют сложный желудок, состоящий из четырёх отделов: рубца, сетки, книжки и сычуга. В рубец попадает масса грубых, не измельчённых зубами растительных кормов, где они подвергаются брожению под влиянием деятельности бактерий и инфузорий. Из рубца пища переходит в сетку, откуда путём отрыгивания попадает в рот для повторного пережёвывания. Смешанная со слюной полужидкая масса проглатывается и попадает в книжку, а оттуда в сычуг (настоящий желудок), где обрабатывается кислым желудочным соком, который переваривает белковую часть корма.
К этому отряду принадлежат все (кроме яка и буйвола) разводимые породы крупного рогатого скота. Они выведены от нескольких видов диких быков. Одним из них был тур, широко распространённый в Европе и Азии и исчезнувший 350 лет назад. Разведение и селекция домашнего скота шли в направлении создания рабочих, молочных, мясных и мясомолочных пород.
Отряд Рукокрылые
многочисленный отряд летающих млекопитающих, распространённых везде, кроме Арктики и Антарктиды. Летают рукокрылые благодаря наличию кожистых перепонок, натянутых между длинными пальцами передних конечностей, боками тела, задними конечностями и хвостом. Как и птицы, на грудине они имеют киль, к которому прикрепляются мощные грудные мышцы, приводящие крылья в движение.
Ведут сумеречный или ночной образ жизни, ориентируясь в воздушном пространстве с помощью звуковой локации. В большинстве случаев приносят пользу, поедая вредных насекомых (летучие мыши), некоторые питаются кровью животных. Строение зубов рукокрылых сходно со строением зубов у насекомоядных (имеются все три рода зубов — резцы, клыки и коренные, но имеется большое разнообразие в их числе и форме).
Отряд Зайцеобразные
(зайцы, кролики). Основное сходство зайцеобразных и грызунов состоит в том, что у них отсутствуют клыки, а резцы и коренные зубы отделены широким, лишённым зубов пространством. От грызунов зайцеобразные отличаются тем, что в верхней челюсти у них не одна, а две пары резцов. Вторая пара резцов у зайцеобразных развита слабее и располагается позади основной пары.
Отряд Ластоногие
включает 30 видов (тюлени, морские котики, моржи). Большую часть жизни они проводят в воде, а на сушу или лёд выходят для размножения и линьки. Благодаря обтекаемой форме тела, укороченным и видоизменённым в ласты конечностям, а также большим подкожным жировым отложениям ластоногие хорошо приспособлены к жизни в водной среде. Питаются в основном рыбой. Являются ценными объектами промысла и дают жир, кожу, мясо, пушнину.
Отряд Китообразные
включает 80 видов. Это исключительно водные млекопитающие, имеющие рыбообразную форму тела с горизонтально расположенным хвостовым плавником. Передние конечности превращены в ласты, задние отсутствуют. Не имеют шерстного покрова и ушных раковин. Подкожный слой жира мощный, достигает 50 см. Удельный вес крупных китообразных близок к удельному весу воды. Зубатые киты (дельфины, кашалоты) питаются рыбой и имеют большое число зубов одинакового строения. У беззубых усатых китов (синий кит) на месте зубов развит цедильный аппарат в виде роговых пластин (китовый ус), сидящих по бокам нёба и свешивающихся в ротовую полость, с помощью которого они отцеживают планктон (ежесуточно синий кит (масса 150 т, длина 33 м) поедает 4–5 т планктона).
Отряд Непарнокопытные
включает 16 видов (лошади, носороги, ослы, зебры). На ногах сильно развит один (третий) палец.
Отряд Обезьяны, или Приматы
включает 190 видов. Головной мозг у Приматов сравнительно больших размеров. Полушария переднего мозга очень большие, имеют многочисленные извилины. Глазницы направлены вперёд. Пальцы имеют ногти. Большой палец конечностей противопоставлен остальным. Живут в тропических и субтропических лесах, ведут как древесный, так и наземный образ жизни. Питаются растительной и животной пищей. Семейство человекообразных обезьян (орангутан, шимпанзе, горилла) обитает в лесах экваториальной и тропической Африки.
Что такое плацентарные млекопитающие
К подклассу первозверей относятся яйцекладущие утконос и ехидна.
Для настоящих зверей характерно внутриутробное вынашивание и живорождение.
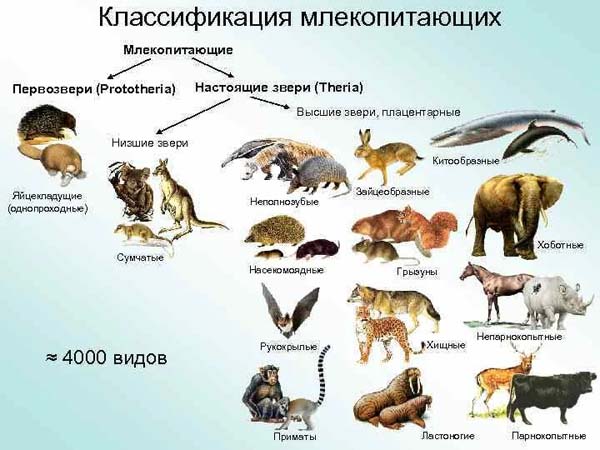
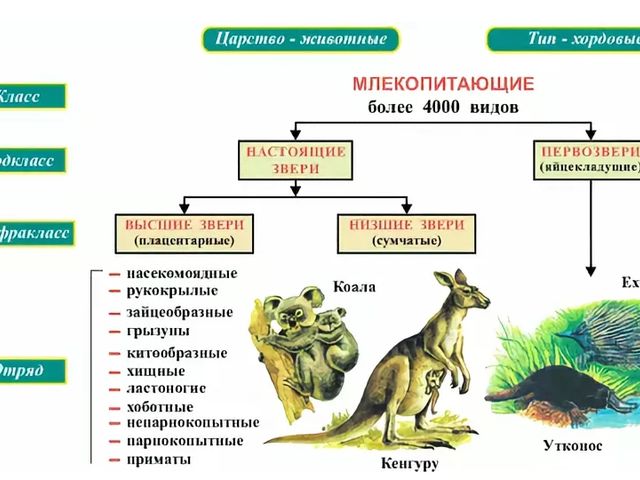
Подкласс настоящих зверей делится на 2 инфракласса:
К низшим зверям относят отряд сумчатые. Они отличаются короткой беременностью и недоразвитостью рождаемого детёныша.
Таким образом, плацентарные, – это основная группа млекопитающих, к которой относится 18 отрядов.
Кора полушарий переднего мозга состоит из нескольких слоёв клеток и содержит центры высшей нервной деятельности. У некоторых видов кора образует борозды, что означает увеличение её поверхности.
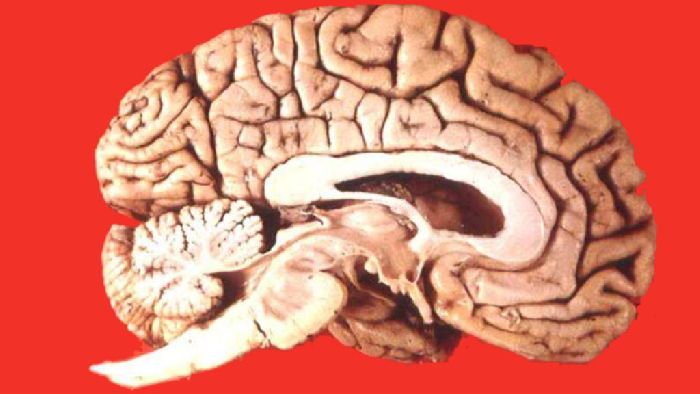
Рис. 1. Мозг высших млекопитающих.
Температура тела плацентарных +38°, в то время как у первозверей +25 – 30°, а у сумчатых +36°.
Плацента
Плацента является временным органом, существующим только в период беременности. Образуется она из клеток материнского и детского организма и является как бы переходным участком между ними.
Слово плацента имеет греческое происхождение и переводится как «лепёшка», т. к. внешне похожа на диск.
После родов плацента выходит вместе с детёнышем во внешнюю среду.
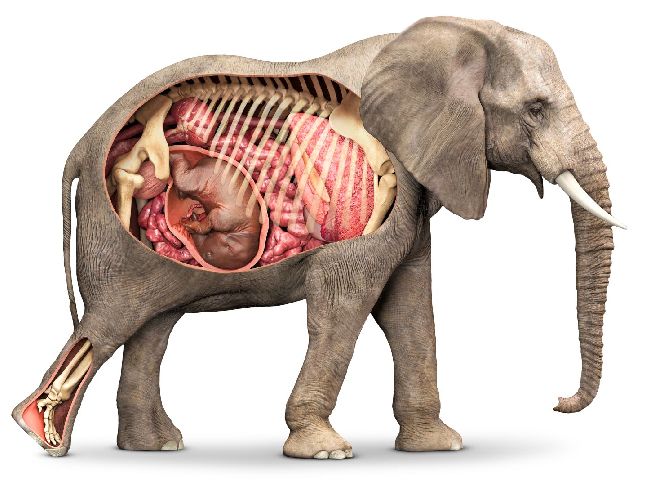
Рис. 2. Детёныш млекопитающего в матке.
Большинство самок млекопитающих после родов съедают свою плаценту, т. к. в ней содержатся вещества, необходимые для восстановления организма после периода беременности.
У сумчатых животных также есть плацента, но она зачаточная и быстро рассасывается. В результате детёныши сумчатых рождаются недоразвитыми. Следовательно, плацента способствует развитию плода.
Среди плацентарных детёныши рождаются с разной степенью развитости. Например, кролики и зайцы относятся к одному отряду, но зайцы рождаются зрячими, а кролики слепыми и голыми.

Рис. 3. Новорожденные крольчонок и зайчонок.
Основные отряды Плацентарных млекопитающих:
К плацентарным млекопитающим относятся все звери, кроме сумчатых и яйцекладущих. Особенностями размножения плацентарных млекопитающих являются: вынашивание детёнышей в матке, образование из плодных оболочек и тканей матки плаценты, обеспечивающей оптимальное питание детёнышей, живорождение.
Плацентарные млекопитающие
Удивительные животные
Ехидны и утконосы — самые необычные представители класса млекопитающих. Однопроходными их называют, потому что и кишечник, и мочевой пузырь этих животных открываются в одну специальную полость — клоаку. Туда же выходят и два яйцевода у самок однопроходных. У большинства млекопитающих нет клоаки; эта полость характерна для рептилий. Желудок у яйцекладущих тоже удивительный — он, как птичий зоб, не переваривает пищу, а лишь хранит ее. Пищеварение происходит в кишечнике. У этих странных млекопитающих даже температура тела ниже, чем у других: не поднимаясь выше 36°С, она может опускаться до 25°С в зависимости от окружающей среды, как у рептилий. Ехидны и утконосы безголосые — у них нет голосовых связок, и беззубые — быстроразрушающиеся зубы есть только у молодых утконосов.
Живут ехидны до 30 лет, утконосы — до 10. Обитают они в лесах, степях, поросших кустарником, и даже в горах на высоте до 2500 м.
Систематика
В классе млекопитающих выделяют 2 подкласса:
К подклассу первозверей относятся яйцекладущие утконос и ехидна.
Для настоящих зверей характерно внутриутробное вынашивание и живорождение.
Подкласс настоящих зверей делится на 2 инфракласса:
К низшим зверям относят отряд сумчатые. Они отличаются короткой беременностью и недоразвитостью рождаемого детёныша.







Таким образом, плацентарные, – это основная группа млекопитающих, к которой относится 18 отрядов.
Кора полушарий переднего мозга состоит из нескольких слоёв клеток и содержит центры высшей нервной деятельности. У некоторых видов кора образует борозды, что означает увеличение её поверхности.
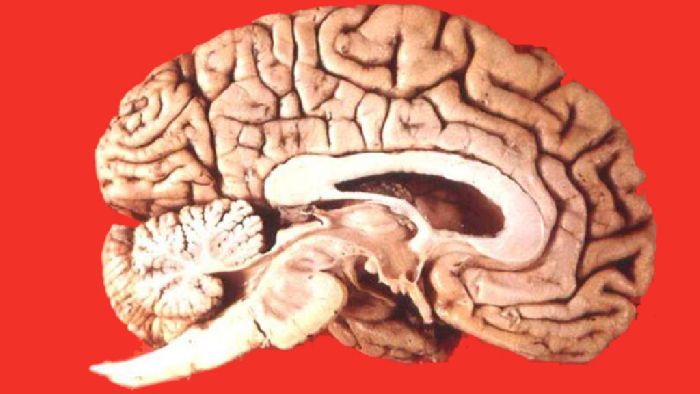
Рис. 1. Мозг высших млекопитающих.
Температура тела плацентарных +38°, в то время как у первозверей +25 – 30°, а у сумчатых +36°.
Утконос
 Собирая пищу со дна, утконос закрывает глаза, ноздри и уши
Собирая пищу со дна, утконос закрывает глаза, ноздри и уши
У утконоса перепончатые лапы, уплощенный, как у бобра, хвост, а морда оканчивается большим клювом, похожим на утиный. Когда первые шкурки утконоса в начале XIX в. попали в Европу, многие ученые посчитали их искусной подделкой: невероятным казалось, что животное, покрытое шерстью, как млекопитающее, имеет птичий клюв. Только спустя 20 лет выяснилось, что у утконосов есть железы, выделяющие молоко, а в конце XIX в. было доказано, что эти животные откладывают яйца. Клюв утконоса вовсе не клюв, а уплощенная морда, покрытая кожей с большим количеством нервных окончаний.








Это главный осязательный орган зверька, с помощью которого он копается в иле на дне водоемов и разыскивает моллюсков, насекомых, червей и ракообразных. Когда наступает пора размножения, самка утконоса роет на берегу нору, а в глубине ее устраивает гнездо из растительного материала, который приносит к норе с помощью цепкого хвоста. В гнезде самка откладывает от 1 до 4 маленьких яиц и около 10 дней согревает их теплом собственного тела. Вылупившиеся детеныши слизывают молоко с ее брюха и быстро растут. В норе они остаются около 4 месяцев, пока не покроются шерстью, а потом начинают сами добывать себе пищу.
Плацента
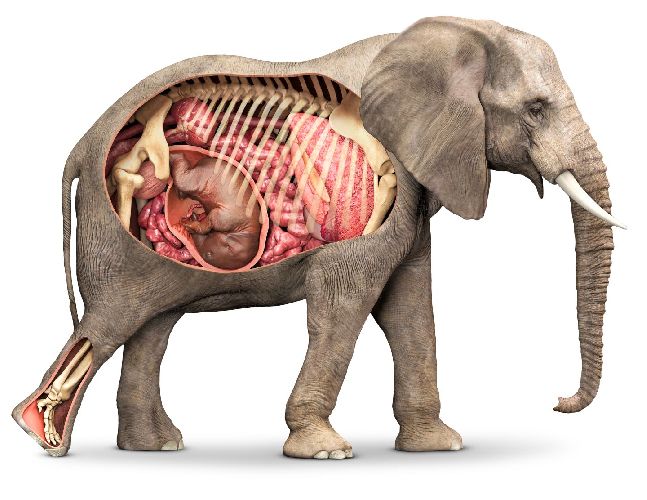
Плацента является временным органом, существующим только в период беременности. Образуется она из клеток материнского и детского организма и является как бы переходным участком между ними.
Слово плацента имеет греческое происхождение и переводится как «лепёшка», т. к. внешне похожа на диск.
После родов плацента выходит вместе с детёнышем во внешнюю среду.
Рис. 2. Детёныш млекопитающего в матке.









Большинство самок млекопитающих после родов съедают свою плаценту, т. к. в ней содержатся вещества, необходимые для восстановления организма после периода беременности.
У сумчатых животных также есть плацента, но она зачаточная и быстро рассасывается. В результате детёныши сумчатых рождаются недоразвитыми. Следовательно, плацента способствует развитию плода.
Среди плацентарных детёныши рождаются с разной степенью развитости. Например, кролики и зайцы относятся к одному отряду, но зайцы рождаются зрячими, а кролики слепыми и голыми.

Рис. 3. Новорожденные крольчонок и зайчонок.
Что мы узнали?
К плацентарным млекопитающим относятся все звери, кроме сумчатых и яйцекладущих. Особенностями размножения плацентарных млекопитающих являются: вынашивание детёнышей в матке, образование из плодных оболочек и тканей матки плаценты, обеспечивающей оптимальное питание детёнышей, живорождение.









Тест по теме
К плацентарным относятся:
Начать тест(новая вкладка)
Особенности яйцекладущих
Самки откладывают 1-3 яйца, покрытых кожистой оболочкой, но детенышей выкармливают молоком, как и другие млекопитающие. Настоящих сосков у них нет — млечные железы открываются наружу порами, скопления которых образуют железистые поля на брюхе самки. Кишечник и мочеточники не имеют отдельных выводящих отверстий, а открываются, как у птиц и пресмыкающихся, в клоаку. Хотя яйцекладущие теплокровные животные, температура тела у них очень низкая и колеблется в широких пределах — от 25 до 36 °С. Зубы у взрослых животных отсутствуют, но у детенышей утконосов имеются зачатки 10 молочных зубов.
Древнейшие плацентарные: начало истории успеха
Алексей Лопатин, Александр Аверьянов
«Природа» №4, 2018

Реконструкции внешнего вида прокенналеста Трофимова (слева) и ховурлеста. Рисунок А. А. Атучина по эскизу А. В. Лопатина
Об авторах

Алексей Владимирович Лопатин — академик РАН, доктор биологических наук, директор Палеонтологического института им. А. А. Борисяка РАН, профессор кафедры палеонтологии геологического факультета Московского государственного университета им. М. В. Ломоносова. Область научных интересов — морфология, филогения и эволюция позвоночных животных, биостратиграфия, палеобиогеография, палеоэкология, антропогенез.

Александр Олегович Аверьянов — профессор РАН, доктор биологических наук, заведующий лабораторией териологии Зоологического института РАН, профессор Института наук о Земле Санкт-Петербургского государственного университета. Научные интересы связаны с морфологией и эволюцией позвоночных животных.


Основные группы плацентарных: а — общий предок Theria (Metatheria + Eutheria), б — общий предок Placentalia, в — стволовые эутерии, г — кроновые плацентарные, д — общая (тотальная) группа Pan-Placentalia, или Eutheria. Серым цветом отмечены вымершие таксоны
Теперь, когда мы договорились о том, кого считать «своими», отметим следующие неожиданные факты. Если вымершие млекопитающие считаются эутериями в систематическом смысле, то это вовсе не означает, что они обязательно имели плаценту. А если метатериями, то не обязательно имели сумку. Первично все млекопитающие были яйцекладущими, как современные утконос и ехидна, и в основании эволюционных ветвей как плацентарных, так и сумчатых тоже находились яйцекладущие звери.
По данным молекулярной биологии предполагается, что дивергенция сумчатых и плацентарных млекопитающих могла произойти 147 млн лет назад [1]. Примерно тем же временем, т.е. раннемеловой эпохой, датируется появление в геологической летописи первых стволовых эутериевых млекопитающих. Тогда сушей безраздельно владели рептилии — многочисленные динозавры и их родственники. Для представителей древних групп млекопитающих (триконодонтов, симметродонтов, мультитуберкулят и др.) оставалась возможность развиваться лишь в мелком размерном классе, что они и делали уже многие миллионы лет. Древние плацентарные довольно быстро заняли важное место в сообществах. Но почему и как это произошло? Чтобы ответить на эти вопросы, нужно проанализировать некоторые аспекты строения скелета, репродуктивной биологии и образа жизни этих вымерших существ.
Пересчитать все зубы
Прежде всего отметим, что большинство находок мезозойских млекопитающих составляют фрагменты челюстей или даже изолированные зубы. Находки целых черепов и скелетов исключительно редки. Поэтому выводы об образе жизни этих древних форм носят преимущественно гипотетический характер. Удивительным образом способ размножения сумчатых млекопитающих нашел отражение в их зубной системе. Дело в том, что новорожденные детеныши сумчатых проводят один-два месяца в сумке матери, прикрепившись к ее соску. Из-за этого у них нарушается нормальная для стволовых териев смена зубов, в результате чего трансформируется зубная формула: у сумчатых в каждой половине челюсти всего три премоляра (предкоренных зуба) и четыре моляра (коренных), первый из которых, на самом-то деле, — несменяемый предкоренной зуб. Соответственно, если мезозойское млекопитающее обладало такой зубной формулой, его определенно следует относить к сумчатым. Также у древних сумчатых было больше резцов (пять верхних и четыре нижних), чем у ранних плацентарных (обычно четыре верхних и три нижних, хотя есть исключения).
Способ размножения плацентарных млекопитающих никак не отразился на их зубной системе. Они унаследовали от стволовых териев зубную формулу с пятью премолярами (сократившимися до четырех лишь у более поздних форм) и тремя молярами. Это создает определенные трудности при идентификации древнейших стволовых плацентарных.
Яркий пример таких трудностей — история с юрамайей (Juramaia sinensis), чей почти полный скелет был обнаружен в китайской провинции Ляонин в верхнеюрских (оксфордских) отложениях возрастом около 160 млн лет [2]. Это был мелкий зверек с длиной тела 7–10 см (без учета длины хвоста) и массой около 15 г. По характеру локомоции он напоминал современных крыс [3], а судя по строению зубов, питался насекомыми. При первоописании в 2011 г. этот род был отнесен к стволовым плацентарным млекопитающим, что вызвало сенсацию. Действительно, остатки эутериев еще никогда не находили в юрских отложениях.
Однако строение зубов юрамайи соответствует не эутериям, а примитивным стволовым териям — т.е. группе зверей, существовавших до расхождения ветвей сумчатых и плацентарных [4]. Несколько форм млекопитающих из раннего мела Северной Америки, обычно относимых к Eutheria [5, 6], — монтаналестес (Montanalestes keeblerorum), голоклеменсия (Holoclemensia texana) и паппотерий (Pappotherium pattersoni) — также должны рассматриваться среди стволовых териев [4].
Есть еще два кандидата на статус древнейших эутериев. Это дурлстотерий (Durlstotherium newmani) и дурлстодон (Durlstodon ensomi), описанные в 2017 г. из нижнемеловых (берриасских) отложений возрастом около 145 млн лет с побережья залива Дурлстон в Южной Англии [7]. Они плохо охарактеризованы: каждый вид представлен всего одним зубом, причем в обоих случаях это последний верхний моляр, диагностическое значение которого нельзя переоценивать. Однако и они демонстрируют по меньшей мере один очень продвинутый признак, неожиданный для столь древних эутериев, — дополнительные бугорки (конули) между внутренним бугорком (протоконом) и наружными (параконом и метаконом) у них приближены к последним. Этой особенностью оба английских вида напоминают так называемых желестид (Zhelestidae) из верхнего мела Центральной Азии. Безусловно, нужны новые находки дурлстотерия и дурлстодона, чтобы точнее понять их положение на филогенетическом древе Eutheria.
Находки несомненных древних эутериев отстоят от начала мелового периода на 15–35 млн лет. Наиболее полные происходят из китайской провинции Ляонин (там, напомним, была найдена юрамайя). Эомайя (Eomaia scansoria) из барремских отложений (возраст 125,5–130 млн лет) известна по целому скелету с отпечатком контуров тела и шерсти [8]. Судя по строению костей конечностей, эомайя вела древесный образ жизни и умела хорошо прыгать. Длина ее тела была около 10 см, а вес составлял 20–25 г. В отличие от более поздних плацентарных, животное имело по пять верхних резцов в каждой половине челюсти и по четыре нижних. Иногда считается, что строение костей таза эомайи (а именно наличие сумчатых костей) определенно свидетельствует, что эти животные не могли рожать крупных детенышей, а значит, физиологически не были плацентарными. Однако сумчатые кости связаны с кожной мускулатурой и поэтому никак не характеризуют размер детородного канала. Судя по зубной формуле (пять премоляров и три моляра в каждой половине челюсти), эомайя не была сумчатым. Это может означать, что эти млекопитающие либо имели плаценту, либо откладывали яйца, подобно однопроходным.
У плацентарных все три гена вителлогенина (белка-предшественника, служащего для построения желтка в ооците) — это псевдогены, утратившие кодирующую способность; у однопроходных работает один из этих генов. По генетическим оценкам [9], инактивация последнего из трех генов у сумчатых произошла не ранее 70 млн лет назад (т.е. в конце мелового периода), а у плацентарных — более 100 млн лет назад (т.е. в его середине). Такая инактивация должна соответствовать рубежу, когда предки этих групп перестали производить желток и откладывать яйца.
Акристатерий (Acristatherium yanensis) найден в отложениях нижнего апта (122,5–125,5 млн лет назад). Известен всего один череп довольно хорошей сохранности [10]. У акристатерия было по четыре верхних резца в каждой половине челюсти и по три нижних. На нижних коренных зубах акристатерия отмечен маленький дополнительный бугорок, служивший для усиления межзубного сцепления в нижнем зубном ряду. Почему-то этот бугорок исчез у других эутериев, исключая ховурлеста, о котором рассказывается ниже.
В России, в Бурятии, был обнаружен муртойлест (Murtoilestes abramovi), чьи остатки представлены тремя изолированными коренными зубами из пограничных отложений баррема и апта (около 122,5 млн лет назад) [11].
Cасаямамил (Sasayamamylos kawaii) был найден в 2007 г. в Японии, в префектуре Хёго, в серии Сасаяма раннеальбского возраста (около 112 млн лет назад) [12]. Описано несколько довольно полных нижнечелюстных фрагментов, по величине которых реконструируется длина тела в 10–15 см. Передняя часть нижней челюсти сасаямамила необычным образом вздернута вверх, из-за чего резцы поставлены почти вертикально, а клык и вовсе загибается назад. На рентгенографии видно, что задний наклон клыков всех экземпляров существенно усилен из-за фрагментации корневой части, но и его вертикальная постановка (в меньшей степени выраженная также у эомайи) свидетельствует о какой-то особой пищевой специализации — возможно, о питании сравнительно крупными насекомыми, например жуками.
И числом, и умением

Полевой лагерь Совместной Советско-Монгольской палеонтологической экспедиции на р. Аргуин-гол. Промывка костеносной породы из местонахождения Ховур. 1971 г. Фото Е. Л. Дмитриевой
Уникальный источник сведений о раннемеловых млекопитающих представляет собой местонахождение Ховур, расположенное на севере пустыни Гоби в Монголии. Возраст местонахождения определяется в пределах аптского — альбского веков раннего мела (110–120 млн лет назад). Здесь найдены многие сотни ископаемых остатков, принадлежащих представителям нескольких групп млекопитающих, в том числе относящихся к древнейшим стволовым плацентарным. Численно доминируют остатки плацентарных, принадлежащие двум видам рода прокенналестов — маленьким зверькам, питавшимся в основном насекомыми и другими наземными беспозвоночными животными. Прокенналест малый (Prokennalestes minor) не превышал величиной современных землероек среднего размера. Прокенналест Трофимова (Prokennalestes trofimovi) был приблизительно на четверть больше. В коллекциях Палеонтологического института имени А. А. Борисяка РАН в Москве и Геологического института Академии наук Монголии в Улан-Баторе хранятся более 500 экземпляров их челюстей и зубов, примерно поровну от каждого вида. Раскопочные работы в урочище Ховур велись Совместной Советско-Монгольской палеонтологической экспедицией в 1969–1971 гг. и возобновились Совместной Российско-Монгольской палеонтологической экспедицией в 2012 г. в новой точке — Зун-Ховур [13].
Детальное исследование всех образцов прокенналестов из Ховура позволило нам получить первые статистически достоверные результаты по изменчивости признаков столь древних млекопитающих на массовом материале (как мы уже отмечали, обычно находки раннемеловых эутериев единичны, если не уникальны) [14]. Также нами описан еще один вид плацентарных, названный ховурлестом (Hovurlestes noyon). Он отличается от прокенналестов более крупными клыками и наличием на коренных зубах маленького бугорка, усиливающего межзубное сцепление [15].

Фрагменты нижних челюстей прокенналеста малого (а) и прокенналеста Трофимова (б). Фото А. О. Аверьянова
На внутренней стороне нижнечелюстных костей прокенналестов (и других раннемеловых эутериев) есть характерный для древних млекопитающих признак — отчетливая меккелева борозда. Это тянущийся вдоль нижнего края кости длинный, узкий и неглубокий желобок, служивший для прикрепления передней части окостенелого меккелева хряща, к заднему концу которого были подвешены слуховые косточки. У взрослых современных млекопитающих слуховые косточки находятся в полости среднего уха и полностью обособлены от нижней челюсти, но в эмбриогенезе рекапитулируется описанное состояние, свойственное прокенналестам и многим другим мезозойским млекопитающим [16]. Резорбция меккелева хряща у современных млекопитающих обеспечивается специальными клетками — хондрокластами. Если по генетическим или фармакологическим причинам хондрокласты утрачиваются, эмбриональный меккелев хрящ сохраняется и окостеневает на взрослой стадии, формируя на нижней челюсти отчетливую борозду [17]. Эволюционный переход к «современному» состоянию косточек среднего уха происходил у плацентарных и сумчатых независимо. Эти преобразования способствовали усилению передаваемого звукового сигнала, увеличивая остроту слуха. Вероятно, они были сопряжены с увеличением и усложнением головного мозга.
Вряд ли факт подавляющего численного превосходства остатков прокенналестов в Ховуре связан с условиями захоронения или особенностями распространения тех или иных групп млекопитающих. По геологическим данным, все эти зверьки вместе с многочисленными ящерицами и мелкими динозаврами жили по берегам озер и проток, существовавших в раннемеловую эпоху на месте нынешней пустыни Гоби, и одновременно гибли во время сезонных наводнений. Тонкие кости их челюстей со слабо закрепленными в альвеолах зубами не вынесли бы переноса водными потоками на большое расстояние. Видимо, массовость остатков прокенналестов действительно отражает их обилие в древнем биоценозе.
Можно предположить, что представители архаичных групп млекопитающих росли несколько медленнее и жили дольше современных зверей такого же размера. Такую «жизненную стратегию» они унаследовали от своих предков — цинодонтов, имевших замедленный по сравнению с современными млекопитающими ювенильный рост и более продолжительный рост у взрослых особей [18]. Анализ возрастной индивидуальной изменчивости у представителей архаичного рода Morganucodon по высоте зубной кости нижней челюсти показал, что переход к свойственной млекопитающим модели произошел уже на эволюционном уровне морганукодонтов в позднетриасовую эпоху, т.е. сразу же при появлении млекопитающих [18]. Однако некоторые ранние млекопитающие сочетали быстрый ювенильный рост и некоторое его замедление у взрослых особей. Например, хищные триконодонты рода гобиконодонов (Gobiconodon), достигавшие размеров енота, продолжали активно расти и во взрослом состоянии. При этом у них несколько раз за жизнь сменялись коренные зубы, обеспечивая возможность нормально питаться, несмотря на снашивание зубных коронок [19].

Фрагмент нижней челюсти ховурлеста. Фото А. О. Аверьянова
Для млекопитающих в целом характерно сочетание быстрого ювенильного роста с детерминированной остановкой роста при достижении взрослого состояния. Еще предки плацентарных, стволовые териевые млекопитающие, приобрели совершенную зубную систему, называемую трибосфенической [20]. При таком строении зубов внутренний бугорок на верхних молярах взаимодействует с углубленным задним выступом на нижних молярах по принципу пестика в ступке, что позволяет эффективно обрабатывать пищу путем ее перетирания и в итоге дает значимые энергетические преимущества. Вероятно, древние плацентарные производили быстро созревавшее потомство, достигавшее взрослой стадии скорее, чем в других группах. Можно предположить, что с этим связана массовость находок плацентарных в таких местонахождениях, как Ховур и Зун-Ховур.

А. В. Лопатин на местонахождении Зун-Ховур, открытом в ходе работ Совместной Российско-Монгольской палеонтологической экспедиции. 2013 г. Фото А. А. Карху
Большинство найденных остатков обоих видов прокенналестов принадлежат взрослым особям, у которых молочные зубы уже сменились на постоянные; при этом средний из пяти премоляров оставался молочным во взрослом состоянии либо выпадал без соответствующей замены [13]. У более поздних эутериев (начиная с сасаямамила) этот зуб вообще исчезает, и в каждой половине челюсти остается только по четыре премоляра. Не исключено, что сокращение числа предкоренных зубов эволюционно связано с ускорением онтогенеза. У современных землероек, живущих всего лишь 1,5 года и становящихся самостоятельными в четырехнедельном возрасте, смены зубов не происходит, так как закладки молочных зубов резорбируются еще на эмбриональной стадии.
Отсутствие остатков молоди прокенналестов в Ховуре может свидетельствовать о низкой ювенильной смертности и указывать на высокий уровень заботы о потомстве, характерный для так называемой K-стратегии размножения, при которой относительно низкая численность приплода компенсируется высоким уровнем его доживания до взрослого состояния. Противоположна ей r-стратегия, суть которой состоит в производстве многочисленного потомства, как правило быстро достигающего самостоятельности и широко расселяющегося, но подверженного высокой смертности в ювенильном возрасте.
Следует также отметить, что с репродуктивно обусловленным усилением метаболизма первично могло быть связано развитие совершенной теплокровности у плацентарных [21].
В итоге можно предположить, что начало эволюционного успеха предковых плацентарных было связано не только с физиологическими преимуществами, обусловленными совершенствованием зубной системы, органов чувств и теплокровности, но и с новой стратегией размножения, включающей относительно длительное эмбриональное созревание, быстрый ювенильный рост, ускоренное взросление и развитую заботу о потомстве.
Работа выполнена при финансовой поддержке Российского фонда фундаментальных исследований (проект 16-04-00294).
Литература
1. Bininda-Emonds O. R. P., Cardillo M., Jones K. E. et al. The delayed rise of present-day mammals // Nature. 2007; 446(7219): 507–512.
2. Luo Z.-X., Yuan C.-X., Meng Q.-J., Ji Q. A Jurassic eutherian mammal and divergence of marsupials and placentals // Nature. 2011; 76(7361): 442–445.
3. Bonnan M. F., Shulman J., Varadharajan R. et al. Forelimb kinematics of rats using XROMM, with implications for small eutherians and their fossil relatives // PLoS One. 2016; 11(3): e0149377.
4. Averianov A. O. Taxonomic revision of tribosphenic mammals from the Lower Cretaceous Antlers Formation of Texas and Oklahoma, USA // Proceedings of the Zoological Institute RAS. 2015; 319: 141–181.
5. Davis B. M., Cifelli R. L. Reappraisal of the tribosphenidan mammals from the Trinity Group (Aptian — Albian) of Texas and Oklahoma // Acta Palaeontologica Polonica. 2011; 56(3): 441–462.
6. Cifelli R. L., Davis B. M. Tribosphenic mammals from the Lower Cretaceous Cloverly Formation of Montana and Wyoming // Journal of Vertebrate Paleontology. 2015; 35(3): e920848.
7. Sweetman S. C., Smith G., Martill D. M. Highly derived eutherian mammals from the earliest Cretaceous of southern Britain // Acta Palaeontologica Polonica. 2017; 62(4): 657–665.
8. Ji Q., Luo Z.-X., Yuan C.-X., et al. The earliest known eutherian mammal // Nature. 2002; 416(6883): 816–822.
9. Brawand D., Wahli W., Kaessmann H. Loss of egg yolk genes in mammals and the origin of lactation and placentation // PLoS Biol. 2008; 6(3): e63.
10. Hu Y., Meng J., Li C., Wang Y. New basal eutherian mammal from the Early Cretaceous Jehol biota, Liaoning, China // Proceedings of the Royal Society. Series B. 2010; 277: 229–236.
11. Averianov A. O., Skutschas P. P. A new genus of eutherian mammal from the Early Cretaceous of Transbaikalia, Russia // Acta Palaeontologica Polonica. 2001; 46(3): 431–436.
12. Kusuhashi N., Tsutsumi Y., Saegusa H. et al. A new Early Cretaceous eutherian mammal from the Sasayama Group, Hyogo, Japan // Proceedings of the Royal Society. Series B. 2013; 280(1759): e 20130142.
13. Лопатин А. В. Новые находки раннемеловых млекопитающих в Монголии // Доклады Академии наук. 2013; 449(4): 491–493.
14. Lopatin A. V., Averianov A. O. The stem placental mammal Prokennalestes from the Early Cretaceous of Mongolia // Paleontological Journal. 2017; 51(12): 1293–1374.
15. Лопатин А. В., Аверьянов А. О. Новое стволовое плацентарное млекопитающее из раннего мела Монголии // Доклады Академии наук. 2018; 478(1): 117–120.
16. Лопатин А. В. Происхождение и ранние этапы эволюции млекопитающих: современное состояние проблемы // Эволюционная и функциональная морфология позвоночных. М., 2017: 182–189.
17. Anthwal N., Urban D. J., Luo Z.-X et al. Meckel’s cartilage breakdown offers clues to mammalian middle ear evolution // Nature Ecology & Evolution. 2017; 1(Art. № 0093): 1–6.
18. O’Meara R. N., Asher R. J. The evolution of growth patterns in mammalian versus nonmammalian cynodonts // Paleobiology. 2016; 42(3): 439–464.
19. Lopatin A. V., Averianov A. O. Gobiconodon (Mammalia) from the Early Cretaceous of Mongolia // Journal of Mammalian Evolution. 2015: 22(1): 17–43.
20. Lopatin A. V., Averianov A. O. An aegialodontid upper molar and the evolution of mammal dentition // Science. 2006: 313(5790): 1092.
21. Levesque D. L., Lovegrove B. G. Increased homeothermy during reproduction in a basal placental mammal // Journal of Experimental Biology. 2014; 217: 1535–1542.



