Плодолистик
Гинецей (лат. Gynaeceum ) — женский репродуктивный орган цветковых растений.
В полных цветах, например лилий, левкоя, пиона и т. д., он занимает центральную часть цветка. Состоит из одной или многих частей, называемых плодниками или плодолистиками (carpellum) (в литературе также используется не вполне точный термин пестик, pistillum), из которых впоследствии образуются плоды.
Если в гинецее один пестик, гинецей называется одночленным, если много — многочленным.
Содержание
Пестик
В каждом пестике различаются три части, а именно:
В завязи заключена одна или несколько семяпочек (gemmulae), называвшихся прежде яичками (ovula). Это очень мелкие, иногда едва заметные тела, подвергающиеся оплодотворению и превращающиеся после того в семена.
Столбик, который у многих растений вовсе не развит или развит весьма слабо, содержит внутри себя канал, выстланный нежной и рыхлой тканью, часто совершенно его выполняющей. Через него происходит оплодотворение.
Рыльце выстлано, подобно каналу столбика, такой же рыхлой тканью, высачивающую из себя густую сахаристую влагу, и принимающую плодотворную пыль.
У двучленных и многочленных гинецеев пестики то между собой свободны, то срастаются. В первом случае многочленность гинецея вполне ясна, во втором — срастание бывает различно. Иногда срастаются одни только завязи и тогда столбиков бывает столько же сколько пестиков в гинецее, а иногда срастание касается и завязей и столбиков, тогда гинецей представится цельным, состоящим как бы из одного пестика, но число их изобличается, в основном, числом рыльцев или, по крайней мере, числом лопастей рыльцев.
Завязь
Многочленная сростная завязь имеет обыкновенно снаружи несколько продольных рёбер, число которых соответствует числу сросшихся частей. Внутри такой сростной завязи имеется весьма часто столько гнёзд, сколько срослось частей, хотя это не может считаться правилом без исключений. По своему положению относительно других частей цветка, завязь может быть верхней или нижней (germen inferum и germen superum). В первом случае все части цветка, а именно чашечка, венчик, тычинки и столбики или рыльца сидят на верхушке самой завязи, она как бы срослась с чашечкой; во втором — она находится в середине цветка, а все остальные её части располагаются ниже её, или на одной с ней плоскости; если же они и прикрепляются выше, то отнюдь не на ней самой, а на краю более или менее вогнутого цветочного ложа; верхнюю завязь называют по этому также свободной, несростной (germen liberum). Такое различие зависит от развития гинецея (см. Цветок и его развитие).
Виды гинецеев
Гинецей у разных растений чрезвычайно разнообразен не только по своему составу, числу частей и указанным выше обстоятельствам, но ещё по форме и относительным размерам своих частей. У голосеменных растений он состоит из двух или нескольких семяпочек, незамкнутых в завязь; вместо завязи у них часто чешуевидный листок, при основании которого они и сидят (сосны, пихты, ели и пр.). У остальных цветковых всегда имеется более или менее замкнутая завязь, вследствие чего их и называют покрытосеменными. Кроме того, имеются такие растения, у которых весь цветок состоит из одного только гинецея даже без всякого при нём покрова (ивы).
В настоящее время разделяют три вида гинецеев:
Значение
В жизни растений, именно в половой, гинецей имеет первенствующее значение, ибо он, после оплодотворения, разрастается в плод с семенами. Сообразно этому его значению и разные части его имеют различную степень важности для растений. Важнейшая его часть есть семяпочка, превращающаяся в семя, Поэтому гинецей без семяпочек лишается своего физиологического значения и справедливо считается недоразвитым.
Нормальных растений без семяпочек нет, имеются однако же растения, гинецей которых представлен только семяпочкой. Затем по важности следует рыльце, то есть орган, принимающий плодотворную пыль. Настоящего рыльца нет только у голосемянных, где оно заменено однако же рыхлой тканью, развивающейся при отверстии семяпочки (микропилярном). Менее важен столбик, которого вовсе нет у многих растений (многие лютиковые, барбарис и пр.). Настоящей завязи не имеется только у голосемянных.
ПЛОДОЛИСТИК
Смотреть что такое «ПЛОДОЛИСТИК» в других словарях:
плодолистик — плодолистик … Орфографический словарь-справочник
ПЛОДОЛИСТИК — ПЛОДОЛИСТИК, часть женского репродуктивного органа у ЦВЕТКА. Плодолистик состоит из РЫЛЬЦА (воспринимающего пыльцу при опылении), развивающегося на верхушке стилодия, или столбика, и семяпочки (где содержатся семязачатки). Группа плодолистиков… … Научно-технический энциклопедический словарь
плодолистик — лист, карпелла Словарь русских синонимов. плодолистик сущ., кол во синонимов: 2 • карпелла (1) • лист … Словарь синонимов
ПЛОДОЛИСТИК — спороносный лист (один или несколько) в цветке, несущий семязачатки и при срастании краями или с другими плодолистиками образующий завязь пестика … Большой Энциклопедический словарь
плодолистик — спороносный лист (один или несколько) в цветке, несущий семязачатки и при срастании краями или с другими плодолистиками образующий завязь пестика. * * * ПЛОДОЛИСТИК ПЛОДОЛИСТИК, спороносный лист (один или несколько) в цветке, несущий семязачатки… … Энциклопедический словарь
плодолистик — vaislapėlis statusas T sritis augalininkystė apibrėžtis Pakitęs augalų žiedo lapelis. Vienas ar keli suaugę vaislapėliai sudaro piestelę. atitikmenys: angl. carpel rus. плодолистик … Žemės ūkio augalų selekcijos ir sėklininkystės terminų žodynas
Плодолистик — (carpellum) орган в цветке покрытосеменных растений, на котором развиваются семезачатки (семяпочки). Из 1 или нескольких П. образуется пестик; совокупность П. называется гинецеем (См. Гинецеи). П. считают органом листового происхождения,… … Большая советская энциклопедия
Плодолистик — (folium carpellare) листовое образование, дающее вследствие срастания пестик. Пестик может состоять из одного или нескольких плодолистиков. См. Гинецей … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
Плодолистик — м. Видоизмененный лист в цветках покрытосеменных растений, на котором развиваются семяпочки. Толковый словарь Ефремовой. Т. Ф. Ефремова. 2000 … Современный толковый словарь русского языка Ефремовой
плодолистик — плодолистик, плодолистики, плодолистика, плодолистиков, плодолистику, плодолистикам, плодолистик, плодолистики, плодолистиком, плодолистиками, плодолистике, плодолистиках (Источник: «Полная акцентуированная парадигма по А. А. Зализняку») … Формы слов
Плодолистики или карпеллы
Полезное
Смотреть что такое «Плодолистики или карпеллы» в других словарях:
Цветок — укороченный и ограниченный в росте репродуктивный побег, выполняющий функции микро и мегаспорогенеза, опыления (осуществляемого при содействии агентов переноса пыльцы), оплодотворения, развития зародыша и образования плода. Ц. заканчивает … Большая советская энциклопедия
Цветок — Запрос «Цветы» перенаправляется сюда; см. также другие значения. Запрос «Цветки» перенаправляется сюда; см. также другие значения. Цветок (множ. цветки, лат. … Википедия
Венчик (ботаника) — Цветок Запрос «Цветы» перенаправляется сюда. Cм. также другие значения. Цветок (лат. flos, греч. ανθοζ) орган семенного размножения цветковых (покрытосеменных) растений. Цветок представляет собой видоизменённый, укороченный и ограниченный в росте … Википедия
Венчик зигоморфный — Цветок Запрос «Цветы» перенаправляется сюда. Cм. также другие значения. Цветок (лат. flos, греч. ανθοζ) орган семенного размножения цветковых (покрытосеменных) растений. Цветок представляет собой видоизменённый, укороченный и ограниченный в росте … Википедия
Двойной околоцветник — Цветок Запрос «Цветы» перенаправляется сюда. Cм. также другие значения. Цветок (лат. flos, греч. ανθοζ) орган семенного размножения цветковых (покрытосеменных) растений. Цветок представляет собой видоизменённый, укороченный и ограниченный в росте … Википедия
Зигоморфный цветок — Цветок Запрос «Цветы» перенаправляется сюда. Cм. также другие значения. Цветок (лат. flos, греч. ανθοζ) орган семенного размножения цветковых (покрытосеменных) растений. Цветок представляет собой видоизменённый, укороченный и ограниченный в росте … Википедия
Листочек (часть околоцветника) — Цветок Запрос «Цветы» перенаправляется сюда. Cм. также другие значения. Цветок (лат. flos, греч. ανθοζ) орган семенного размножения цветковых (покрытосеменных) растений. Цветок представляет собой видоизменённый, укороченный и ограниченный в росте … Википедия
Околоцветник — Цветок Запрос «Цветы» перенаправляется сюда. Cм. также другие значения. Цветок (лат. flos, греч. ανθοζ) орган семенного размножения цветковых (покрытосеменных) растений. Цветок представляет собой видоизменённый, укороченный и ограниченный в росте … Википедия
Околоцветник двойной — Цветок Запрос «Цветы» перенаправляется сюда. Cм. также другие значения. Цветок (лат. flos, греч. ανθοζ) орган семенного размножения цветковых (покрытосеменных) растений. Цветок представляет собой видоизменённый, укороченный и ограниченный в росте … Википедия
Околоцветник простой — Цветок Запрос «Цветы» перенаправляется сюда. Cм. также другие значения. Цветок (лат. flos, греч. ανθοζ) орган семенного размножения цветковых (покрытосеменных) растений. Цветок представляет собой видоизменённый, укороченный и ограниченный в росте … Википедия
плодолистики или карпеллы
Внутреннюю часть цветка, т. е. верхушечную область цветоложа (которая у вдавленного цветоложа может быть расположена ниже остальной его части), занимают плодолистики, или карпеллы. Термин «плодолистик», представляющий собой русский перевод немецкого термина Fruchtblatt, явно неудачен, так как уже в самом названии органа дана определенная (в данном случае листовая) интерпретация. Еще в 1817 г. был введен удачный термин саrpellum (латинизированное уменьшительное от греч. karpos — плод), первоначально для обозначения элементарной части плода, а затем он широко вошел в латинскую ботаническую терминологию и в английский и романские языки для обозначения плодолистика. В прошлом он переводился на русский язык словом «плодник», которое, однако, впоследствии вышло из употребления. Нередко плодолистик называют пестиком (простым пестиком), однако этого термина желательно избегать, так как он применяется в довольно разных смыслах и вносит только путаницу.
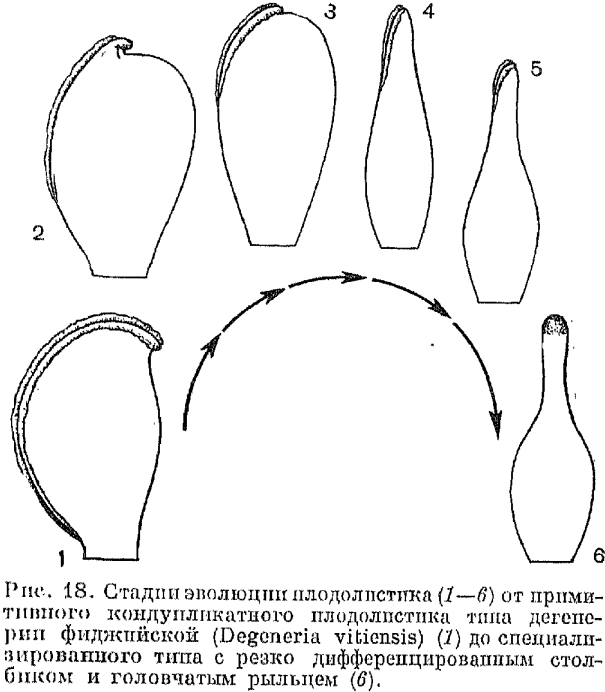
Функционально и морфологически плодолистики (карпеллы) соответствуют мегаспорофиллам. Относительно наиболее примитивный тип плодолистика сохранился у упомянутого нами рода дегенерия (рис. 18), а также у рода тасманния из семейства винтеровых. У них плодолистик состоит из короткой ножки, или гиноподия (gynopodium, от греч. gyne —- женщина и pus (podos) — нога), и относительно тонкой, сложенной вдоль средней жилки (кондупликатной) пластинки.
Если мы развернем плодолистик дегенерии, то перед нами будет более или менее выемчатая на верхушке листовидная пластинка, вдоль которой проходят три самостоятельные жилки (проводящих пучка), средняя (дорсальная) из которых сильно ветвится. Между средней жилкой и двумя боковыми расположены многочисленные семязачатки. Примитивность таких плодолистиков подчеркивается также тем, что некоторые из них не достигли еще полной замкнутости. Наиболее замечателен в этом отношении опять-таки плодолистик дегенерии, края которого, за исключением нижней их части, не только не срастаются, но фактически едва соприкасаются.
Защита семязачатков («покрытосемянность») обеспечивается только тем, что во время цветения боковые участки плодолистика, расположенные между его краями и семязачатками, тесно сближены и, кромо того, вход в плодолистик закрывают многочисленные короткие ягелезистые волоски. Срастание этих сближенных поверхностей происходит лишь после цветения, в процессе развития плода. У представителей семейства винтеровых плодолистики более замкнутые, причем в пределах семейства наблюдаются самые различные стадии срастания. У видов примитивного рода тасманния все еще наблюдается отсутствие срастания краев плодолистиков, но краевые зоны здесь сближены более тесно, чем у дегенерии. У более подвинутого рода дримис (Drimys) сближенные участки плодолистика частично или даже полностью срастаются во время цветения. Интересно, что неполное срастание краев плодолистика наблюдается также у некоторых других, более подвинутых растений, например у платана, где плодолистики в верхней части несколько приоткрыты.
С возникновением плодолистика, даже такого примитивного, как плодолистик дегенерии, прямой доступ пыльцевых зерен к заключенным в нем семязачаткам затрудняется. Хотя края плодолистика еще даже не соприкасаются, но они покрыты железистыми полосками, которые надежно защищают вход в его полость непрошеных посетителей. Железистые волоски играют и другую роль: они воспринимают пыльцу и способствуют своими выделениями ее прорастанию. Пыльцевые зерна, доставленные на цветок насекомыми-опылителями, прилипают к железистым волоскам и, прорастая на них, выпускают пыльцевую трубку, врастающую внутрь плодолистика и направляющуюся в сторону семязачатков. Таким образом, здесь уже имеется специальная рыльцовая область, которая у дегенерии протягивается широкой полоской по всей длине плодолистика. При этом рыльцовая область но ограничена только отогнутыми наружу свободными краями плодолистика (что хорошо видно на рис. 18), но образует внутри его полости довольно широкие полоски, доходящие почти до семязачатков.
Рыльцевая область плодолистика дегенерии представляет собой очень примитивное образование, которое еще далеко от типичного рыльца. У видов тасманнии рыльцовая поверхность также протягивается вдоль всего плодолистика, но у видов дримиса она уже более ограничена и локализована в его верхушечной области. Довольно примитивные рыльцевые структуры мы встречаем и у некоторых других представителей винтеровых, а также у лимонника, багрянника, эвптелеи, платана и некоторых других. Но гораздо чаще, в том числе даже в большинстве примитивных групп цветковых растений, имеется типичное рыльце, более или менее локализованное в верхушечной части плодолистика.
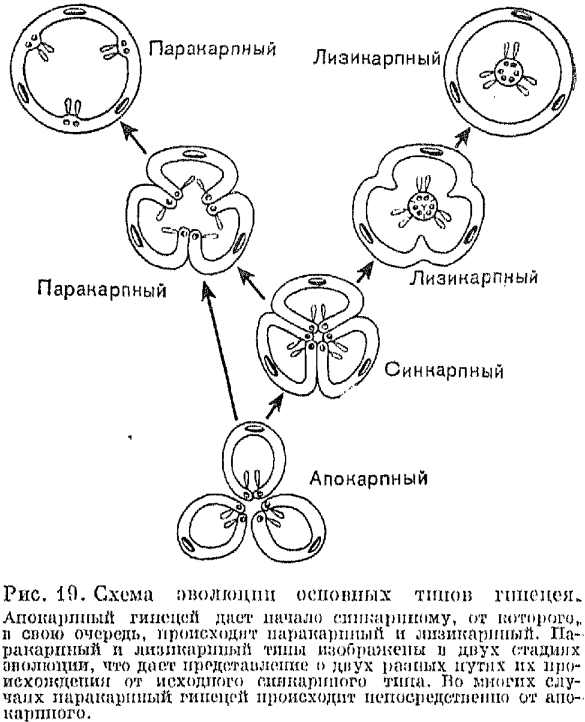
В процессе эволюции гинецея плодолистики постепенно срастаются меяеду собой и из апокарпного гинецея возникает ценокарпный (от греч. kainos — общий; рис. 19). В ценокарпном гинецее отдельные столбики (стилодии) могут оставаться свободными или же срастаются, образуя один общий столбик (сложный столбик). Ценокарпный гинецей бывает трех типов: синкарпный, паракарпный и лизикарпный.
Синкарпным (от греч. syn — вместе) называется гинецей из различного числа замкнутых плодолистиков, сросшихся между собой боковыми частями (рис. 19). Он дву-, многогнездный и характеризуется тем, что семязачатки расположены вдоль швов замкнутых плодолистиков, т. е. по углам, образованным брюшными их частями (так называемая угловая плацентация). Синкарпный гинецей обычно происходит из апокарпного гинецея с циклическим (круговым) расположением плодолистиков, но в некоторых случаях он явился производным и от спирального гинецея. Хорошим примером синкарпного гинецея могут служить лилия и тюльпан. На первых порах эволюции синкарпного гинецея срастаются только завязи плодолистиков, а столбики (стилодии) остаются свободными. Но постепенно процесс срастания захватывает также столбики, которые в конце концов срастаются в один сложный столбик, заканчивающийся рыльцевой головкой, что можно видеть у вересковых или у большинства однодольных, в том числе у лилии.
Во многих других случаях паракарпный гинецей произошел скорее всего непосредственно из апокарпного. Таково, вероятное всего, происхождение паракарпного гинецея некоторых африканских родов семейства ашюновых, большинство представителей которого характеризуется апокарпным гинецеем, или же происхождение паракарпного гиницея семейства капелловых. Таково же, по всем данным, происхождение паракарпного гинецея представителей семейств савруровых и кактусовых, всего порядка фиалковых и целого ряда происшедших от него порядков, а также семейства водокрасовых среди однодольных.
В паракарпном гинецее обычно происходит разрастание плацент, которые часто даже ветвятся. В очень многих случаях эти разросшиеся плаценты сталкиваются в центре завязи, срастаются между собой и полость одногнездного гинецея разделяется на камеры, представляющие собой ложные гнезда. Разросшиеся плаценты хорошо выражены у некоторых тыквенных, например у огурца и арбуза.
В некоторых линиях развития двудольных, например у портулаковых и первоцветных, из синкарпного гинецея возник лизикарпный (от греч. lysis — развязывание, освобождение, растворение). Это одногнездный гинецей, у которого плацентация не париетальная, как у гинецея паракарпного типа, а так называемая свободная центральная, или колончатая. Одиогнездность лизикарпного гинецея возникает в процессе эволюции (или в онтогенезе) в результате исчезновения перегородок (боковых стенок) синкарпной завязи. При этом сросшиеся краевые части не разрушаются и не расходятся, и на них по-прежнему продолжают сидеть семязачатки. Таким образом, эти сохраняющиеся краевые части плодолистиков вместе с плацентами превращаются в центральную колонку, возвышающуюся в полости завязи.
Кроме этих трех основных типов ценокарпного гинецея, различают еще так называемый псевдомономерный гинецей. Это очень редуцированный вариант синкарпного или паракарпного гинецея, в котором фертилен и полностью развит лишь один плодолистик. Стерильные плодолистики в псевдомономерном гинецее часто настолько редуцированы, что их присутствие удается обнаружть только в результате специального изучения их анатомического строения и развития. Типичный псевдомономерный гинецей можно наблюдать у ильма, шелковицы, крапивы, конопли и родственных им растений.
У многих цветковых растений, в том числе у примитивных групп, гинецей еще свободный, не сросшийся с окружающими его частями цветка. Но во многих линиях эволюции цветковых растений совершенно независимо и параллельно произошло большее или меньшее срастание гинецея (как правило, ценокарпного) с окружающими частями цветка и в результате возникла так называемая нижняя завязь. Этот процесс происходил постепенно, и имеются все промежуточные формы между верхней и нижней завязью. Поэтому в морфологии цветка, кроме терминов «верхняя» и «нижняя» завязь, существует также термин «нолунижняя» завязь. Происхождение нижней завязи является одним из трудных вопросов эволюционной морфологии растений, и ему посвящена большая литература. Для решения этого вопроса большое значение имело сравнительно-анатомическое изучение цветка, особенно его проводящей системы. Значение проводящей системы в решении различного рода морфологических вопросов основано на ее сравнительно большем постоянстве (консерватизме). Данные по анатомии проводящей системы цветков с нижней завязью приводят к выводу, что в подавляющем большинстве случаев нижняя завязь произошла в результате срастания с завязью оснований чашелистиков, лепестков и тычиночных нитей. Прирастание к завязи тесно облегающей ее цветочной трубки вполне естественно и биологически оправдано. Разные стадии этого прирастания цветочной трубки к завязи,можно проследить, например, в семействе вересковых. Однако, как показали те же сравнительно-анатомические исследования, происхождение нижней завязи может быть и иное. У некоторых семейств, таких, как аизооповые, кактусовые, санталовые и родственные им семейства, а также у некоторых других завязь обрастает не цветочная трубка, а цветоложе. Ход проводящих пучков ясно показывает, что завязь как бы погрузилась в чашеобразное углубление цветоложа и срослась с ним. Однако этот второй, путь образовании нижней завязи наблюдается лишь у очень небольшого числа семейств.
Что такое плодолистики у растений




Плодолистики, или карпеллы (А. Л. Тахтаджян)
Если мы развернем плодолистик дегенерии, то перед нами будет более или менее выемчатая на верхушке листовидная пластинка, вдоль которой проходят три самостоятельные жилки (проводящих пучка), средняя (дорсальная) из которых сильно ветвится. Между средней жилкой и двумя боковыми расположены многочисленные семязачатки. Примитивность таких плодолистиков подчеркивается также тем, что некоторые из них не достигли еще полной замкнутости. Наиболее замечателен в этом отношении опять-таки плодолистик дегенерии, края которого, за исключением нижней их части, не только не срастаются, но фактически едва соприкасаются.
Защита семязачатков («покрытосемянность») обеспечивается только тем, что во время цветения боковые участки плодолистика, расположенные между его краями и семязачатками, тесно сближены и, кроме того, вход в плодолистик закрывают многочисленные короткие железистые волоски. Срастание этих сближенных поверхностей происходит лишь после цветения, в процессе развития плода. У представителей семейства винтеровых плодолистики более замкнутые, причем в пределах семейства наблюдаются самые различные стадии срастания. У видов примитивного рода тасманния все еще наблюдается отсутствие срастания краев плодолистиков, но краевые зоны здесь сближены более тесно, чем у дегенерии. У более подвинутого рода дримис (Drimys) сближенные участки плодолистика частично или даже полностью срастаются во время цветения. Интересно, что неполное срастание краев плодолистика наблюдается также у некоторых других, более подвинутых растений, например у платана, где плодолистики в верхней части несколько приоткрыты.
С возникновением плодолистика, даже такого примитивного, как плодолистик дегенерии, прямой доступ пыльцевых зерен к заключенным в нем семязачаткам затрудняется. Хотя края плодолистика еще даже не соприкасаются, но они покрыты железистыми волосками, которые надежно защищают вход в его полость непрошеных посетителей. Железистые волоски играют и другую роль: они воспринимают пыльцу и способствуют своими выделениями ее прорастанию. Пыльцевые зерна, доставленные на цветок насекомыми-опылителями, прилипают к железистым волоскам и, прорастая на них, выпускают пыльцевую трубку, врастающую внутрь плодолистика и направляющуюся в сторону семязачатков. Таким образом, здесь уже имеется специальная рыльцевая область, которая у дегенерии протягивается широкой полоской по всей длине плодолистика. При этом рыльцевая область не ограничена только отогнутыми наружу свободными краями плодолистика (что хорошо видно на рис. 18), но образует внутри его полости довольно широкие полоски, доходящие почти до семязачатков.
Рыльцевая область плодолистика дегенерии представляет собой очень примитивное образование, которое еще далеко от типичного рыльца. У видов тасманнии рыльцевая поверхность также протягивается вдоль всего плодолистика, но у видов дримиса она уже более ограничена и локализована в его верхушечной области. Довольно примитивные рыльцевые структуры мы встречаем и у некоторых других представителей винтеровых, а также у лимонника, багрянника, эвптелеи, платана и некоторых других. Но гораздо чаще, в том числе даже в большинстве примитивных групп цветковых растений, имеется типичное рыльце, более или менее локализованное в верхушечной части плодолистика.
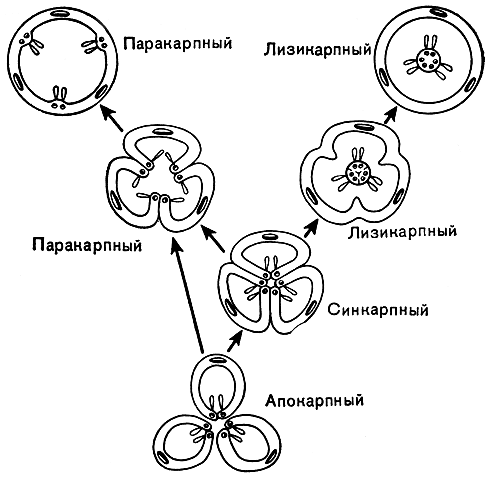
Рис. 19. Схема эволюции основных типов гинецея. Апокарпный гинецей дает начало синкарпному, от которого, в свою очередь, происходят паракарпный и лизикарпный. Паракарпный и лизикарпный типы изображены в двух стадиях эволюции, что дает представление о двух разных путях их происхождения от исходного синкарпного типа. Во многих случаях паракарпный гинецей происходит непосредственно от апокарпного
По мере того как рыльцевая поверхность локализуется в верхней части плодолистика, эта последняя обычно вытягивается в стерильное столбиковидное образование, приподнимающее рыльце над фертильной частью (завязью) плодолистика и служащее для прохождения пыльцевой трубки. Эта специализированная более тонкая верхняя часть плодолистика называется столбиком (соответствует латинскому термину stylus). По предложению немецкого ботаника М. Ханфа (1935) иногда столбик отдельного плодолистика называют стилодием (stylodium), оставляя термин «столбик» только для сросшихся стилодиев. Таким образом, примитивный плодолистик типа дегенерии постепенно дифференцируется на завязь и столбик (стилодий). На ранних стадиях эволюции столбик еще кондупликатный, с явственной брюшной бороздой и примитивным низбегающим рыльцем, состоящим из двух так называемых рыльцевых гребней, соответствующих отогнутым краям верхней части плодолистика (рис. 18). В процессе дальнейшей эволюции низбегающие рыльцевые гребни постепенно укорачиваются, и в конце концов рыльце локализуется на верхушке столбика. Так возникает характерное для многих семейств цветковых растений головчатое рыльце (рис. 18). Но даже и головчатое рыльце часто имеет более или менее двулопастный характер. Эти две лопасти напоминают нам о двух рыльцевых гребнях предковых форм.
Во многих других случаях паракарпный гинецей произошел скорее всего непосредственно из апокарпного. Таково, вероятнее всего, происхождение паракарпного гинецея некоторых африканских родов семейства анноновых, большинство представителей которого характеризуется апокарпным гинецеем, или же происхождение паракарпного гинецея семейства канелловых. Таково же, по всем данным, происхождение паракарпного гинецея представителей семейств савруровых и кактусовых, всего порядка фиалковых и целого ряда происшедших от него порядков, а также семейства водокрасовых среди однодольных.
В паракарпном гинецее обычно происходит разрастание плацент, которые часто даже ветвятся. В очень многих случаях эти разросшиеся плаценты сталкиваются в центре завязи, срастаются между собой и полость одногнездного гинецея разделяется на камеры, представляющие собой ложные гнезда. Разросшиеся плаценты хорошо выражены у некоторых тыквенных, например у огурца и арбуза.
Кроме этих трех основных типов ценокарпного гинецея, различают еще так называемый псевдо мономерный гинецей. Это очень редуцированный вариант синкарпного или паракарпного гинецея, в котором фертилен и полностью развит лишь один плодолистик. Стерильные плодолистики в псевдомономерном гинецее часто настолько редуцированы, что их присутствие удается обнаружить только в результате специального изучения их анатомического строения и развития. Типичный псевдомономерный гинецей можно наблюдать у ильма, шелковицы, крапивы, конопли и родственных им растений.
У многих цветковых растений, в том числе у примитивных групп, гинецей еще свободный, не сросшийся с окружающими его частями цветка. Но во многих линиях эволюции цветковых растений совершенно независимо и параллельно произошло большее или меньшее срастание гинецея (как правило, ценокарпного) с окружающими частями цветка и в результате возникла так называемая нижняя завязь. Этот процесс происходил постепенно, и имеются все промежуточные формы между верхней и нижней завязью. Поэтому в морфологии цветка, кроме терминов «верхняя» и «нижняя» завязь, существует также термин «полунижняя» завязь. Происхождение нижней завязи является одним из трудных вопросов эволюционной морфологии растений, и ему посвящена большая литература. Для решения этого вопроса большое значение имело сравнительно-анатомическое изучение цветка, особенно его проводящей системы. Значение проводящей системы в решении различного рода морфологических вопросов основано на ее сравнительно большем постоянстве (консерватизме). Данные по анатомии проводящей системы цветков с нижней завязью приводят к выводу, что в подавляющем большинстве случаев нижняя завязь произошла в результате срастания с завязью оснований чашелистиков, лепестков и тычиночных нитей. Прирастание к завязи тесно облегающей ее цветочной трубки вполне естественно и биологически оправдано. Разные стадии этого прирастания цветочной трубки к завязи можно проследить, например, в семействе вересковых. Однако, как показали те же сравнительно-анатомические исследования, происхождение нижней завязи может быть и иное. У некоторых семейств, таких, как аизооновые, кактусовые, санталовые и родственные им семейства, а также у некоторых других завязь обрастает не цветочная трубка, а цветоложе. Ход проводящих пучков ясно показывает, что завязь как бы погрузилась в чашеобразное углубление цветоложа и срослась с ним. Однако этот второй путь образования нижней завязи наблюдается лишь у очень небольшого числа семейств.



