полиаденилирование
Смотреть что такое «полиаденилирование» в других словарях:
полиаденилирование — Введение в матричную РНК эукариот полиадениловой кислоты на 3’конце [http://www.dunwoodypress.com/148/PDF/Biotech Eng Rus.pdf] Тематики биотехнологии EN polyadenylation … Справочник технического переводчика
Полиаденилирование — * поліадэніляванне * polyadenylation ферментативное посттранскрипционное добавление поли(А) хвостов (см.) размером в 200 адениновых остатков к 31 окончанию гетерогенной ядерной РНК и матричной (информационной) РНК у эукариот во время ее… … Генетика. Энциклопедический словарь
Процессинг (биология) — Схема синтеза белка рибосомой Биосинтез белка сложный многостадийный процесс синтеза полипептидной цепи из аминокислотных остатков, происходящий на рибосомах клеток живых организмов с участием молекул иРНК и тРНК. Содержание 1 Введение … Википедия
Процессинг РНК — (посттранскрипционные модификации РНК) совокупность процессов в клетках эукариот, которые приводят к превращению первичного транскрипта РНК в зрелую РНК. Наиболее известен процессинг матричных РНК, которые во время своего синтеза… … Википедия
МАТРИЧНЫЕ РИБОНУКЛЕИНОВЫЕ КИСЛОТЫ — (информационные РНК, мРНК, иРНК), молекулы РНК, представляющие собой комплементарные (см. Комплементарность) копии участков значащих цепей генов (ДНК), в к рых закодирована информация об аминокислотных последовательностях полипептидных цепей… … Химическая энциклопедия
Клеточное ядро — Клетки HeLa, ДНК которых окрашена голубым красителем Хёхста 33258. Центральная и правая клетки находятся в интерфазе, по … Википедия
МРНК — Основные этапы жизненного цикла мРНК эукариот Матричная рибонуклеиновая кислота (мРНК, синоним информационная РНК, иРНК) РНК, содержащая информацию о первичной структуре (аминокислотной последовательности) белков[1]. мРНК синт … Википедия
Центральная догма молекулярной биологии — Центральная догма молекулярной биологии обобщающее наблюдаемое в природе правило реализации генетической информации: информация передаётся от нуклеиновых кислот к белку, но не в обратном направлении. Правило было сформулировано Френсисом… … Википедия
ИРНК — Основные этапы жизненного цикла мРНК эукариот Матричная рибонуклеиновая кислота (мРНК, синоним информационная РНК, иРНК) РНК, отвечающая за перенос информации о первичной структуре белков от ДНК к местам синтеза белков. мРНК синтезируется на… … Википедия
Информационная РНК — Основные этапы жизненного цикла мРНК эукариот Матричная рибонуклеиновая кислота (мРНК, синоним информационная РНК, иРНК) РНК, отвечающая за перенос информации о первичной структуре белков от ДНК к местам синтеза белков. мРНК синтезируется на… … Википедия
Полиаденилирование
Полиаденилирование начинается, когда завершается транскрипция гена, то есть образование первичного транскрипта. Перед началом полиаденилирования особый мультисубъединичный белковый комплекс отщепляет 3′-концевой участок первичного транскрипта. Место расщепления определяется положением универсальных сигнальных последовательностей в первичном транскрипте; в некоторых случаях расщепление может происходить в нескольких альтернативных сайтах. Таким образом, полиаденилирование даёт возможность для образования различных мРНК одного гена (альтернативное полиаденилирование), подобно тому, как это происходит при альтернативном сплайсинге. После формирования нового 3′-конца транскрипта компонент белкового комплекса поли(А)-полимераза осуществляет синтез поли(А)-хвоста, используя 3′-концевой нуклеотид как затравку.
Поли(А)-хвост играет важную роль в транспорте мРНК из ядра, её трансляции и стабильности. Со временем поли(А)-хвост укорачивается, и, когда его длина станет достаточно малой, мРНК разрушается под действием специальных ферментов. Однако в клетках некоторых типов мРНК с короткими поли(А)-хвостами запасаются в цитозоле для дальнейшей активации путём реполиаденилирования. У бактерий, напротив, полиаденилирование запускает разрушение транскрипта. Подобный эффект полиаденилирования отмечен и для некоторых эукариотических некодирующих РНК.
Связанные понятия
ДНК-метилтрансфера́зы (ДНК-метилазы, англ. DNA methyltransferase, DNA MTase, DNMT) — группа ферментов, катализирующих метилирование нуклеотидных остатков в составе ДНК. Активность метилтрансфераз, заключающаяся в переносе метильных (CH3—) групп на азотистое основание цитозин в составе ДНК, ведет к изменению свойств ДНК, при этом изменяется активность, функции соответствующих генов, а также пространственная структура нуклеиновой кислоты (конформация).
Нуклеазы — большая группа ферментов, гидролизующих фосфодиэфирную связь между субъединицами нуклеиновых кислот. Различают несколько типов нуклеаз в зависимости от их специфичности: экзонуклеазы и эндонуклеазы, рибонуклеазы и дезоксирибонуклеазы, рестриктазы и некоторые другие. Рестриктазы занимают важное положение в прикладной молекулярной биологии.

Полиаденилирование это добавление поли (А) хвост к транскрипту РНК, обычно информационная РНК (мРНК). Поли (А) хвост состоит из нескольких аденозинмонофосфаты; другими словами, это участок РНК, который имеет только аденин базы. В эукариоты, полиаденилирование является частью процесса образования зрелой мРНК для перевод. Во многих бактерииполи (А) хвост способствует деградации мРНК. Следовательно, это часть более широкого процесса экспрессия гена.
Процесс полиаденилирования начинается с транскрипция из ген прекращается. В 3′-самый сегмент новообразованной пре-мРНК сначала отщепляется набор белков; эти белки затем синтезируют поли (А) хвост на 3′-конце РНК. В некоторых генах эти белки добавляют поли (А) хвост в одном из нескольких возможных сайтов. Следовательно, полиаденилирование может производить более одного транскрипта из одного гена (альтернативное полиаденилирование), похожий на альтернативное сращивание. [1]
Поли (A) хвост важен для ядерного экспорта, трансляции и стабильности мРНК. Хвост со временем укорачивается, и, когда он достаточно короткий, мРНК подвергается ферментативной деградации. [2] Однако в некоторых типах клеток мРНК с короткими поли (A) хвостами сохраняются для последующей активации путем повторного полиаденилирования в цитозоле. [3] Напротив, когда полиаденилирование происходит у бактерий, оно способствует деградации РНК. [4] Иногда то же самое и с эукариотическими некодирующие РНК. [5] [6]
Молекулы мРНК как у прокариот, так и у эукариот имеют полиаденилированные 3′-концы, причем поли (А) хвосты прокариот обычно короче, а молекулы мРНК полиаденилированы. [7]
Содержание
Справочная информация о РНК
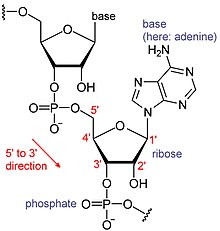
Ядерное полиаденилирование
Функция
При ядерном полиаденилировании поли (А) хвост добавляется к РНК в конце транскрипции. На мРНК поли (А) хвост защищает молекулу мРНК от ферментативной деградации в цитоплазма и помогает в терминации транскрипции, экспорте мРНК из ядра и трансляции. [2] Почти все мРНК эукариот полиаденилированы, [12] за исключением репликационно-зависимых гистон мРНК. [13] Это единственные мРНК у эукариот, у которых отсутствует поли (А) хвост, вместо этого заканчивающийся стебель-петля структура, за которой следует богатая пуринами последовательность, называемая нижележащим элементом гистона, который указывает место разрезания РНК, так что образуется 3′-конец мРНК гистона. [14]
Механизм
CPSF: фактор специфичности расщепления / полиаденилирования
CstF: фактор стимуляции расщепления
PAP: полиаденилатполимераза
ПАБИИ: полиаденилат-связывающий белок 2
CFI: фактор расщепления I
CFII: фактор расщепления II
РНК обычно расщепляется до терминации транскрипции, поскольку CstF также связывается с РНК-полимеразой II. [28] Благодаря плохо изученному механизму (по состоянию на 2002 г.) он дает сигнал РНК-полимеразе II, чтобы ускользнуть из транскрипта. [29] В расщеплении также участвует белок CFII, но неизвестно, каким образом. [30] Сайт расщепления, связанный с сигналом полиаденилирования, может варьироваться примерно до 50 нуклеотидов. [31]
Когда РНК расщепляется, начинается полиаденилирование, катализируемое полиаденилатполимеразой. Полиаденилат-полимераза строит поли (A) хвост, добавляя аденозинмонофосфат единиц от аденозинтрифосфат к РНК, отщепляя пирофосфат. [32] Другой белок, PAB2, связывается с новым коротким поли (A) хвостом и увеличивает сродство полиаденилатполимеразы к РНК. Когда поли (А) хвост составляет примерно 250 нуклеотиды долго фермент больше не может связываться с CPSF, и полиаденилирование прекращается, что определяет длину поли (A) хвоста. [33] [34] CPSF находится в контакте с РНК-полимеразой II, что позволяет ей сигнализировать полимеразе о прекращении транскрипции. [35] [36] Когда РНК-полимераза II достигает «терминирующей последовательности» (‘TTTATT³ на матрице ДНК и ⁵’AAUAAA³ на первичном транскрипте), сигнализируется окончание транскрипции. [37] Аппарат полиаденилирования также физически связан с сплайсосома, комплекс, удаляющий интроны из РНК. [26]
Последующие эффекты
Деаденилирование
В эукариотических соматические клеткиполи (А) хвосты большинства мРНК в цитоплазме постепенно становятся короче, а мРНК с более коротким поли (А) хвостом транслируются меньше и быстрее разрушаются. [44] Однако может пройти много часов, прежде чем мРНК распадется. [45] Этот процесс деаденилирования и деградации может быть ускорен микроРНК, комплементарными 3 ‘нетранслируемая область мРНК. [46] В незрелые яйцеклетки, мРНК с укороченными поли (A) хвостами не деградируют, а вместо этого хранятся и трансляционно неактивны. Эти короткохвостые мРНК активируются цитоплазматическим полиаденилированием после оплодотворения, во время активация яйца. [47]
Цитоплазматическое полиаденилирование
В цитозоле некоторых типов животных клеток, а именно в цитозоле, происходит полиаденилирование. линия зародыша, в начале эмбриогенез и в пост-синаптический сайты нервные клетки. Это удлиняет поли (А) хвост мРНК с укороченным поли (А) хвостом, так что мРНК будет переведено. [44] [49] Эти укороченные поли (A) хвосты часто состоят из менее 20 нуклеотидов и удлинены примерно до 80–150 нуклеотидов. [3]
В раннем эмбрионе мыши цитоплазматическое полиаденилирование материнских РНК из яйцеклетки позволяет клетке выживать и расти, даже если транскрипция не начинается до середины 2-клеточной стадии (4-клеточная стадия у человека). [50] [51] В мозге цитоплазматический полиаденилирование активно во время обучения и может играть роль в долгосрочное потенцирование, который является усилением передачи сигнала от нервной клетки к другой в ответ на нервные импульсы и важен для обучения и формирования памяти. [3] [52]
Для цитоплазматического полиаденилирования необходимы РНК-связывающие белки. CPSF и CPEB, и может включать другие РНК-связывающие белки, такие как Пумилио. [53] В зависимости от типа клетки, полимераза может быть полиаденилат-полимеразой (PAP) того же типа, что и в ядерном процессе, или цитоплазматической полимеразой. GLD-2. [54]
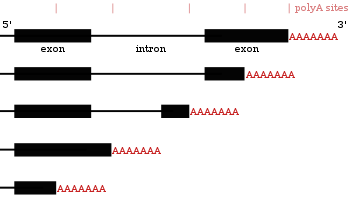
Альтернативное полиаденилирование
Многие гены, кодирующие белок, имеют более одного сайта полиаденилирования, поэтому ген может кодировать несколько мРНК, которые различаются по своему 3 ′ конец. [27] [55] [56] 3’-область транскрипта содержит множество сигналов полиаденилирования (PAS). Когда используются более проксимальные (ближе к 5’-концу) сайты PAS, это укорачивает длину 3 ’нетранслируемой области (3’ UTR) транскрипта. [57] Исследования как на людях, так и на мухах показали тканеспецифический APA. Нейронные ткани предпочитают использование дистальных PAS, что приводит к более длинным 3 ’UTR, а ткани семенников предпочитают проксимальные PAS, что приводит к более коротким 3’ UTR. [58] [59] Исследования показали, что существует корреляция между уровнем консервативности гена и его тенденцией к альтернативному полиаденилированию, при этом высококонсервативные гены демонстрируют больше APA. Точно так же высоко экспрессируемые гены следуют той же схеме. [60] Рибо-секвенирование данные (секвенирование только мРНК внутри рибосом) показали, что изоформы мРНК с более короткими 3 ’UTR транслируются с большей вероятностью. [57]
Поскольку альтернативное полиаденилирование изменяет длину 3 ‘UTR, [61] он также может изменить, какие сайты привязки доступны для микроРНК в 3 ′ UTR. [19] [62] МикроРНК имеют тенденцию подавлять трансляцию и способствовать деградации мРНК, с которыми они связываются, хотя есть примеры микроРНК, которые стабилизируют транскрипты. [63] [64] Альтернативное полиаденилирование также может укорачивать кодирующую область, тем самым создавая код мРНК для другого белка, [65] [66] но это гораздо менее распространено, чем просто укорачивание нетранслируемой 3’-области. [27]
Выбор сайта поли (А) может зависеть от внеклеточных стимулов и зависит от экспрессии белков, которые принимают участие в полиаденилировании. [67] [68] Например, выражение CstF-64, подразделение фактор стимуляции расщепления (CstF), увеличивается макрофаги в ответ на липополисахариды (группа бактериальных соединений, вызывающих иммунный ответ). Это приводит к отбору слабых поли (A) сайтов и, следовательно, более коротких транскриптов. Это удаляет регуляторные элементы в 3′-нетранслируемых областях мРНК для продуктов, связанных с защитой, таких как лизоцим и TNF-α. Эти мРНК имеют более длительный период полураспада и продуцируют больше этих белков. [67] РНК-связывающие белки, отличные от тех, которые входят в аппарат полиаденилирования, также могут влиять на использование сайта полиаденилирования, [69] [70] [71] [72] сканирование Метилирование ДНК рядом с сигналом полиаденилирования. [73]
Метка деградации у эукариот
Cell Biology.ru
Справочник
Полиаденилирование
мРНК гистоновых генов и ряда других не полиаденилируется. Полиаденилированные про-mРНК подвергаются сплайсингу. После синтеза про-mРНК на расстоянии примерно 20 н в направлении 3′-конца от последовательности 5′-AAУAA-3′ происходит разрезание эндонуклеазой и к новому 3′-концу присоединяется от 30 до 300 остатков АМФ (безматричный синтез). ПолиА хвост защищает 3′-конец от гидролиза, т.к. покрыт полиА-связывающими белками. Время жизни mРНК зависит от длинны полиА-хвоста.
Polyadenylation is the covalent linkage of a polyadenylyl moiety to a messenger RNA (mRNA) molecule. It is part of the route to producing mature messenger RNA for translation, in the larger process of protein synthesis to produce proteins. In eukaryotic organisms, polyadenylation is the mechanism by which most messenger RNA molecules are terminated at their 3′ ends. The polyadenosine (poly-A) tail protects the mRNA molecule from exonucleases and is important for transcription termination, for export of the mRNA from the nucleus, and for translation. Some prokaryotic mRNAs also are polyadenylated, although the polyadenosine tail’s function is different from that in eukaryotes.
Polyadenylation occurs during and immediately after transcription of DNA into RNA in the nucleus. After transcription has been terminated, the mRNA chain is cleaved through the action of an endonuclease complex associated with RNA polymerase. The cleavage site is characterized by the presence of the base sequence AAUAAA near the cleavage site. After the mRNA has been cleaved, 50 to 250 adenosine residues are added to the free 3′ end at the cleavage site. This reaction is catalyzed by polyadenylate polymerase.
The Process of PolyadenylationCleavage and Polyadenylation Specificity Factor (CPSF) and Cleavage Stimulation Factor (CstF), both of which are multi-protein complexes, start bound to the rear of the advancing RNA polymerase II.
As the RNA polymerase II advances over the adenylation signal sequences CPSF and CstF transfer to the new pre-mRNA, CPSF binding to the AAUAAA sequence, and CstF to the GU or U rich sequence following it.
CPSF and CstF promote cleavage approximately 35 nucleotides after the end of the AAUAAA sequence. Immediately Polyadenylate Polymerase (PAP) starts writing the polyadenosine tail. Polyadenosine Binding (PAB) protein immediately binds to the new polyadenosine sequence.
CPSF dissociates, and polyadenylation by PAP continues to write an adenosine tail of approximately 50 to 250 nucleotides, depending on the organism. PAB acts as some kind of molecular ruler, specifying when polyadenylation should stop.
PAP dissociates, and PAB remains bound. Along whith the 5′ cap it is thought this helps target the mRNA for nuclear export.
Polyadenylation is initially dependent on CPSF and the AAUAAA sequence (for the first 10 As or so), after which polyadenylation is simply dependent on the existing poly A tail.
Полиаденилирование это добавление поли (А) хвост к транскрипту РНК, обычно информационная РНК (мРНК). Поли (А) хвост состоит из нескольких аденозинмонофосфаты; другими словами, это участок РНК, который имеет только аденин базы. В эукариоты, полиаденилирование является частью процесса образования зрелой мРНК для перевод. Во многих бактерииполи (А) хвост способствует деградации мРНК. Следовательно, это часть более широкого процесса экспрессия гена.
Процесс полиаденилирования начинается с транскрипция из ген прекращается. В 3′-самый сегмент новообразованной пре-мРНК сначала отщепляется набор белков; эти белки затем синтезируют поли (А) хвост на 3′-конце РНК. В некоторых генах эти белки добавляют поли (А) хвост в одном из нескольких возможных сайтов. Следовательно, полиаденилирование может производить более одного транскрипта из одного гена (альтернативное полиаденилирование), похожий на альтернативное сращивание. [1]
Поли (A) хвост важен для ядерного экспорта, трансляции и стабильности мРНК. Хвост со временем укорачивается, и, когда он достаточно короткий, мРНК подвергается ферментативной деградации. [2] Однако в некоторых типах клеток мРНК с короткими поли (A) хвостами сохраняются для последующей активации путем повторного полиаденилирования в цитозоле. [3] Напротив, когда полиаденилирование происходит у бактерий, оно способствует деградации РНК. [4] Иногда то же самое и с эукариотическими некодирующие РНК. [5] [6]
Молекулы мРНК как у прокариот, так и у эукариот имеют полиаденилированные 3′-концы, причем поли (А) хвосты прокариот обычно короче, а молекулы мРНК полиаденилированы. [7]
Содержание
Справочная информация о РНК
Ядерное полиаденилирование
Функция
При ядерном полиаденилировании поли (А) хвост добавляется к РНК в конце транскрипции. На мРНК поли (А) хвост защищает молекулу мРНК от ферментативной деградации в цитоплазма и помогает в терминации транскрипции, экспорте мРНК из ядра и трансляции. [2] Почти все мРНК эукариот полиаденилированы, [12] за исключением репликационно-зависимых гистон мРНК. [13] Это единственные мРНК у эукариот, у которых отсутствует поли (А) хвост, вместо этого заканчивающийся стебель-петля структура, за которой следует богатая пуринами последовательность, называемая нижележащим элементом гистона, который указывает место разрезания РНК, так что образуется 3′-конец мРНК гистона. [14]
Механизм
CPSF: фактор специфичности расщепления / полиаденилирования
CstF: фактор стимуляции расщепления
PAP: полиаденилатполимераза
ПАБИИ: полиаденилат-связывающий белок 2
CFI: фактор расщепления I
CFII: фактор расщепления II
РНК обычно расщепляется до терминации транскрипции, поскольку CstF также связывается с РНК-полимеразой II. [28] Благодаря плохо изученному механизму (по состоянию на 2002 г.) он дает сигнал РНК-полимеразе II, чтобы ускользнуть из транскрипта. [29] В расщеплении также участвует белок CFII, но неизвестно, каким образом. [30] Сайт расщепления, связанный с сигналом полиаденилирования, может варьироваться примерно до 50 нуклеотидов. [31]
Когда РНК расщепляется, начинается полиаденилирование, катализируемое полиаденилатполимеразой. Полиаденилат-полимераза строит поли (A) хвост, добавляя аденозинмонофосфат единиц от аденозинтрифосфат к РНК, отщепляя пирофосфат. [32] Другой белок, PAB2, связывается с новым коротким поли (A) хвостом и увеличивает сродство полиаденилатполимеразы к РНК. Когда поли (А) хвост составляет примерно 250 нуклеотиды долго фермент больше не может связываться с CPSF, и полиаденилирование прекращается, что определяет длину поли (A) хвоста. [33] [34] CPSF находится в контакте с РНК-полимеразой II, что позволяет ей сигнализировать полимеразе о прекращении транскрипции. [35] [36] Когда РНК-полимераза II достигает «терминирующей последовательности» (‘TTTATT³ на матрице ДНК и ⁵’AAUAAA³ на первичном транскрипте), сигнализируется окончание транскрипции. [37] Аппарат полиаденилирования также физически связан с сплайсосома, комплекс, удаляющий интроны из РНК. [26]
Последующие эффекты
Деаденилирование
В эукариотических соматические клеткиполи (А) хвосты большинства мРНК в цитоплазме постепенно становятся короче, а мРНК с более коротким поли (А) хвостом транслируются меньше и быстрее разрушаются. [44] Однако может пройти много часов, прежде чем мРНК распадется. [45] Этот процесс деаденилирования и деградации может быть ускорен микроРНК, комплементарными 3 ‘нетранслируемая область мРНК. [46] В незрелые яйцеклетки, мРНК с укороченными поли (A) хвостами не деградируют, а вместо этого хранятся и трансляционно неактивны. Эти короткохвостые мРНК активируются цитоплазматическим полиаденилированием после оплодотворения, во время активация яйца. [47]
Цитоплазматическое полиаденилирование
В цитозоле некоторых типов животных клеток, а именно в цитозоле, происходит полиаденилирование. линия зародыша, в начале эмбриогенез и в пост-синаптический сайты нервные клетки. Это удлиняет поли (А) хвост мРНК с укороченным поли (А) хвостом, так что мРНК будет переведено. [44] [49] Эти укороченные поли (A) хвосты часто состоят из менее 20 нуклеотидов и удлинены примерно до 80–150 нуклеотидов. [3]
В раннем эмбрионе мыши цитоплазматическое полиаденилирование материнских РНК из яйцеклетки позволяет клетке выживать и расти, даже если транскрипция не начинается до середины 2-клеточной стадии (4-клеточная стадия у человека). [50] [51] В мозге цитоплазматический полиаденилирование активно во время обучения и может играть роль в долгосрочное потенцирование, который является усилением передачи сигнала от нервной клетки к другой в ответ на нервные импульсы и важен для обучения и формирования памяти. [3] [52]
Для цитоплазматического полиаденилирования необходимы РНК-связывающие белки. CPSF и CPEB, и может включать другие РНК-связывающие белки, такие как Пумилио. [53] В зависимости от типа клетки, полимераза может быть полиаденилат-полимеразой (PAP) того же типа, что и в ядерном процессе, или цитоплазматической полимеразой. GLD-2. [54]
Альтернативное полиаденилирование
Многие гены, кодирующие белок, имеют более одного сайта полиаденилирования, поэтому ген может кодировать несколько мРНК, которые различаются по своему 3 ′ конец. [27] [55] [56] 3’-область транскрипта содержит множество сигналов полиаденилирования (PAS). Когда используются более проксимальные (ближе к 5’-концу) сайты PAS, это укорачивает длину 3 ’нетранслируемой области (3’ UTR) транскрипта. [57] Исследования как на людях, так и на мухах показали тканеспецифический APA. Нейронные ткани предпочитают использование дистальных PAS, что приводит к более длинным 3 ’UTR, а ткани семенников предпочитают проксимальные PAS, что приводит к более коротким 3’ UTR. [58] [59] Исследования показали, что существует корреляция между уровнем консервативности гена и его тенденцией к альтернативному полиаденилированию, при этом высококонсервативные гены демонстрируют больше APA. Точно так же высоко экспрессируемые гены следуют той же схеме. [60] Рибо-секвенирование данные (секвенирование только мРНК внутри рибосом) показали, что изоформы мРНК с более короткими 3 ’UTR транслируются с большей вероятностью. [57]
Поскольку альтернативное полиаденилирование изменяет длину 3 ‘UTR, [61] он также может изменить, какие сайты привязки доступны для микроРНК в 3 ′ UTR. [19] [62] МикроРНК имеют тенденцию подавлять трансляцию и способствовать деградации мРНК, с которыми они связываются, хотя есть примеры микроРНК, которые стабилизируют транскрипты. [63] [64] Альтернативное полиаденилирование также может укорачивать кодирующую область, тем самым создавая код мРНК для другого белка, [65] [66] но это гораздо менее распространено, чем просто укорачивание нетранслируемой 3’-области. [27]
Выбор сайта поли (А) может зависеть от внеклеточных стимулов и зависит от экспрессии белков, которые принимают участие в полиаденилировании. [67] [68] Например, выражение CstF-64, подразделение фактор стимуляции расщепления (CstF), увеличивается макрофаги в ответ на липополисахариды (группа бактериальных соединений, вызывающих иммунный ответ). Это приводит к отбору слабых поли (A) сайтов и, следовательно, более коротких транскриптов. Это удаляет регуляторные элементы в 3′-нетранслируемых областях мРНК для продуктов, связанных с защитой, таких как лизоцим и TNF-α. Эти мРНК имеют более длительный период полураспада и продуцируют больше этих белков. [67] РНК-связывающие белки, отличные от тех, которые входят в аппарат полиаденилирования, также могут влиять на использование сайта полиаденилирования, [69] [70] [71] [72] сканирование Метилирование ДНК рядом с сигналом полиаденилирования. [73]



