Что такое постсинаптическая мембрана
Мембрану пресинаптической терминали называют пресинаптической мембраной. Она содержит большое количество электроуправляемых кальциевых каналов. Когда потенциал действия деполяризует пресинаптическую мембрану, эти кальциевые каналы открываются, что позволяет большому числу ионов кальция войти в терминалы Количество медиатора, которое затем выделится из терминали в синаптическую щель, находится в прямой зависимости от числа вошедших ионов кальция.
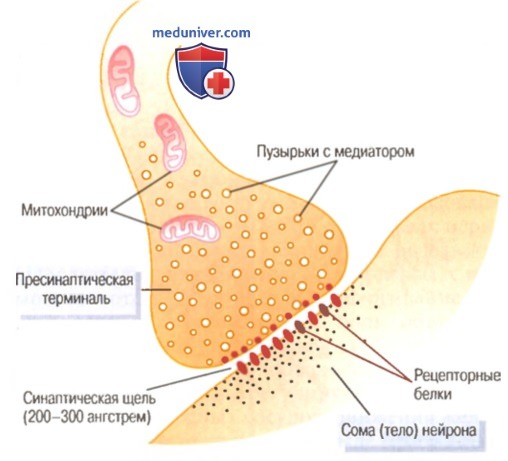 Физиологическая анатомия синапса
Физиологическая анатомия синапса
Точный механизм, с помощью которого ионы кальция вызывают выделение медиатора, не известен, но предполагают следующее.
Считают, что ионы кальция, войдя в пресинаптическую терминаль, связываются с особыми белковыми молекулами на внутренней поверхности пресинаптической мембраны, которые называют местами освобождения. Это связывание вызывает открытие мест освобождения медиатора сквозь мембрану, в результате после каждого одиночного потенциала действия несколько пузырьков выделяют свой медиатор в щель.
В каждом из пузырьков, содержащих медиатор ацетилхолин, накапливается от 2000 до 10000 его молекул, а общего числа пузырьков в пресинаптической терминали достаточно, чтобы проводить от нескольких сотен до более 10000 потенциалов действия.
Молекулы этих рецепторов имеют два важных компонента:
(1) связывающий компонент, который выдается наружу от постсинаптической мембраны в синаптическую щель, где он связывает медиатор, выделившийся из пресинаптической терминали;
(2) ионофорный компонент, проходящий сквозь постсинаптическую мембрану внутрь постсинаптического нейрона.
Ионофор, в свою очередь, может быть одним из двух типов:
(1) ионным каналом, позволяющим определенным ионам проходить через этот канал;
(2) активатором вторичного посредника, представляющим собой молекулу, которая выдается внутрь клетки в ее цитоплазму и активирует одно или более веществ внутри постсинаптического нейрона.
Эти вещества, в свою очередь, служат вторичными посредниками для усиления или угнетения специфических клеточных функций.
б) Ионные каналы. Ионные каналы в мембране постсинаптического нейрона обычно бывают двух типов:
(1) катионные каналы, которые в открытом состоянии обычно пропускают ионы Na+, но иногда также ионы К+ и/или Са 2+ ;
(2) анионные каналы, пропускающие главным образом ионы Cl- и минимальные количества других анионов.
Катионные каналы, проводящие ионы Na+, выстланы отрицательными зарядами. Эти заряды затягивают положительно заряженные ионы Na+ в канал, если диаметр канала превышает размер гидратированного иона Na+. Однако те же самые отрицательные заряды отталкивают ионы Сl- и другие анионы, препятствуя их прохождению.
Если диаметры анионных каналов достаточно велики, ионы Сl- входят в них и проходят на противоположную сторону, тогда как вход натриевых, калиевых и кальциевых катионов блокируется в основном из-за слишком больших размеров их гидратированных ионов.
Далее мы узнаем, что процесс открытия катионных каналов, позволяющий войти в клетку положительно заряженным ионам Na+, способствует возбуждению нейрона, поэтому медиаторы, ведущие к открытию катионных каналов, называют возбуждающими. И наоборот, процесс открытия анионных каналов, позволяющий войти отрицательным электрическим зарядам, тормозит нейрон. Соответственно, медиаторы, открывающие эти каналы, называют тормозными.
При активации медиатором ионного канала он обычно открывается в течение долей миллисекунды; после удаления медиатора канал закрывается так же быстро. Такое открытие и закрытие каналов обеспечивает очень быстрое управление постсинаптическими нейронами.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Что такое постсинаптическая мембрана
а) Однонаправленное проведение в химических синапсах. Одна из чрезвычайно важных характеристик химических синапсов обусловливает проведение сигналов в нервной системе в одном направлении, а именно: от пресинаптического нейрона, секретирующего медиатор, к постсинаптическому нейрону, на который медиатор действует. Эту особенность называют принципом однонаправленного проведения для химических синапсов, что существенно отличает химические синапсы от электрических, где сигналы часто распространяются в любом направлении. Механизм однонаправленного проведения очень важен, поскольку он позволяет сигналам направляться к специфическим мишеням.
Действительно, именно это специфическое проведение сигналов к различным и четко локализованным мишеням, как в пределах нервной системы, так и в терминалях периферических нервов, позволяет нервной системе выполнять ее многочисленные функции восприятия, двигательного контроля, памяти и многие другие.
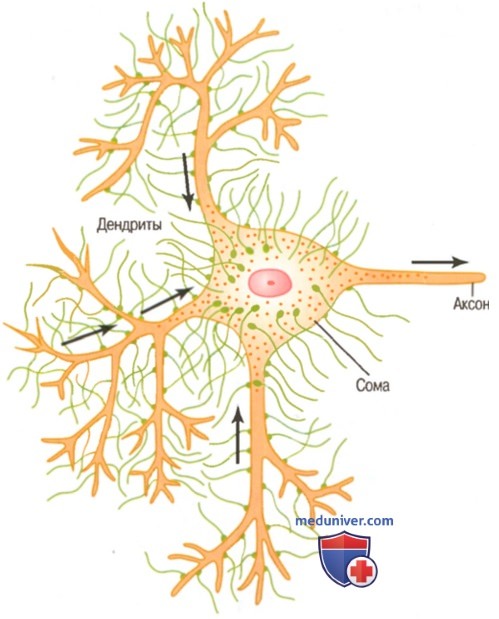 Типичный мотонейрон переднего рога спинного мозга, на соме и дендритах которого расположены многочисленные пресинаптические терминали. Хорошо виден также единственный аксон
Типичный мотонейрон переднего рога спинного мозга, на соме и дендритах которого расположены многочисленные пресинаптические терминали. Хорошо виден также единственный аксон
б) Физиологическая анатомия синапса. На рисунке выше показан типичный мотонейрон переднего рога спинного мозга. Он состоит из трех основных частей: сомы, которая является главной частью нейрона; одного аксона, который распространяется от сомы и входит в периферический нерв, выходящий из спинного мозга; дендритов, представляющих собой большое число ветвящихся отростков сомы, распространяющихся на расстояние до 1 мм в окружающие области спинного мозга.
На поверхности дендритов и сомы мотонейрона находятся от 10000 до 200000 мельчайших синаптических бугорков, называемых пресинаптинескими терминалями, причем примерно 80-95% из них — на дендритах и лишь 5-20% — на соме. Эти пресинаптические терминали являются окончаниями нервных волокон, отходящих от многих других нейронов. Далее станет очевидным, что многие из этих терминалей — возбуждающие, т.е. секретируют медиатор, который возбуждает постсинаптический нейрон. Однако другие пресинаптические терминали являются тормозными, т.е. секретируют медиатор, тормозящий постсинаптический нейрон.
Нейроны в других частях спинного и головного мозга отличаются от передних мотонейронов:
(1) размером тела клетки;
(2) длиной, диаметром и числом дендритов, причем длина колеблется в пределах от почти нуля до многих сантиметров;
(3) длиной и размером аксонов;
(4) числом пресинаптических терминалей, которое может варьировать от нескольких единиц до 200000.
Это обусловливает различную реакцию нейронов разных отделов нервной системы на входящие синаптические сигналы и, следовательно, на их способность выполнять разнообразные функции.
в) Пресинаптические терминали. Электронномикроскопические исследования пресинаптических терминалей показывают, что их анатомические формы разные, но большинство из них похожи на маленькие круглые или овальные узелки, поэтому их иногда называют терминальными узелками, пуговками, концевыми ножкамиияи синаптическими бугорками.
Физиологическая анатомия синапса
На рисунке выше показана характерная структура синапса. Видна одиночная пресинаптическая терминаль на поверхности мембраны постсинаптического нейрона. Пресинаптическая терминаль отделяется от сомы постсинаптического нейрона синаптической щелью, имеющей обычно ширину 200-300 ангстрем. Терминаль имеет две внутренние структуры, важные для возбуждающей или тормозной функции синапса: пузырьки с медиатором и митохондрии.
Пузырьки содержат медиатор, который при выделении возбуждает или тормозит постсинаптический нейрон (возбуждает, если мембрана нейрона содержит возбуждающие рецепторы, и тормозит, если мембрана содержит тормозящие рецепторы). Митохондрии обеспечивают аденозинтри-фосфат, снабжающий энергией процессы синтеза новых молекул медиатора.
Когда по пресинаптической терминали распространяется потенциал действия, деполяризация ее мембраны вызывает опорожнение небольшого числа везикул в щель. Выделенный медиатор, в свою очередь, немедленно вызывает изменение проницаемости мембраны постсинаптического нейрона, что ведет к возбуждению или торможению постсинаптического нейрона в зависимости от характеристик нейрональных рецепторов.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Что такое постсинаптическая мембрана
Область контакта между двумя нейронами называют синапсом.
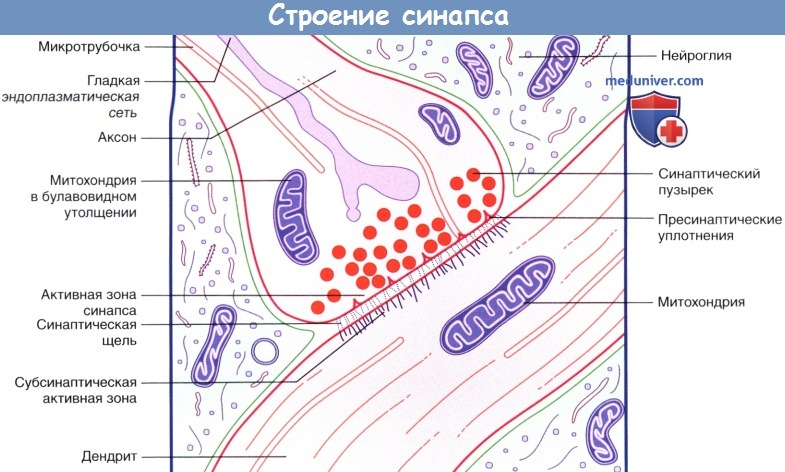 Внутреннее строение аксодендритического синапса.
Внутреннее строение аксодендритического синапса.
а) Электрические синапсы. Электрические синапсы в нервной системе млекопитающих встречаются редко. Они образованы щелевидными контактами (нексусами) между дендритами или сомами соприкасающихся нейронов, которые соединяются с помощью цитоплазматических каналов диаметром 1,5 нм. Процесс передачи сигнала происходит без синаптической задержки и без участия медиаторов.
Посредством электрических синапсов возможно распространение электротонических потенциалов от одного нейрона к другому. Вследствие тесного синаптического контакта модуляция проведения сигнала невозможна. Задача этих синапсов — осуществление одновременного возбуждения нейронов, выполняющих одинаковую функцию. Примером служат нейроны дыхательного центра продолговатого мозга, которые во время вдоха синхронно генерируют импульсы. Кроме того, примером могут служить нейронные цепи, управляющие саккадами, при которых точка фиксации взора перемещается от одного объекта внимания к другому.
б) Химические синапсы. Большинство синапсов нервной системы — химические. Функционирование таких синапсов зависит от высвобождения медиаторов. Классический химический синапс представлен пресинаптической мембраной, синаптической щелью и постсинаптической мембраной. Пресинаптическая мембрана — часть булавовидного расширения нервного окончания клетки, передающей сигнал, а постсинаптическая мембрана — часть клетки, получающей сигнал.
Медиатор высвобождается из булавовидного расширения посредством экзоцитоза, проходит через синаптическую щель и связывается с рецепторами на постсинаптической мембране. Под постсинаптической мембраной расположена субсинаптическая активная зона, в которой после активации рецепторов постсинаптической мембраны происходят разнообразные биохимические процессы.
В булавовидном расширении расположены содержащие медиаторы синаптические пузырьки, а также большое количество митохондрий и цистерны гладкой эндоплазматической сети. Применение традиционных методик фиксации при исследовании клеток позволяет различить на пресинаптической мембране пресинаптические уплотнения, ограничивающие активные зоны синапса, к которым при помощи микротрубочек направляются синаптические пузырьки.
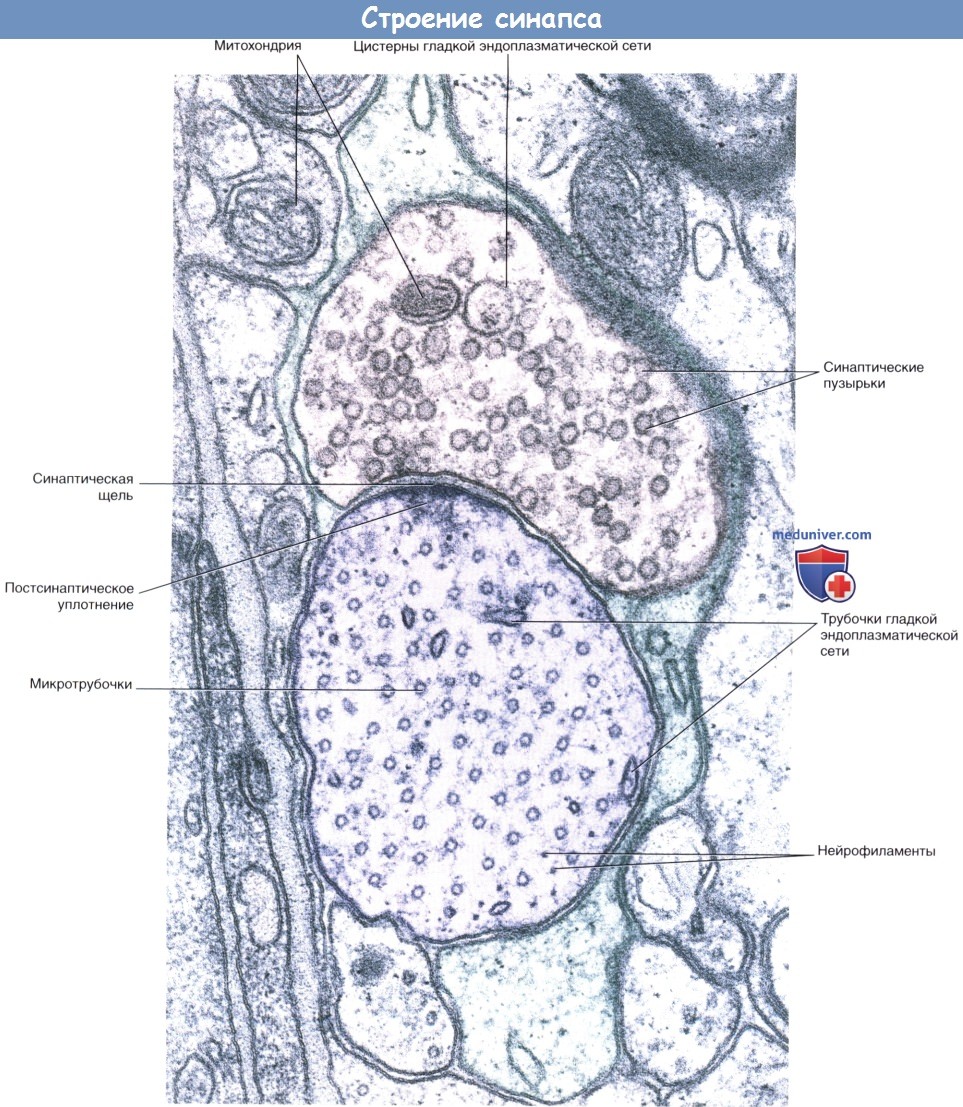 Аксодендритический синапс.
Аксодендритический синапс.
Срез препарата спинного мозга: синапс между концевым участком дендрита и, предположительно, двигательным нейроном.
Наличие округлых синаптических пузырьков и постсинаптического уплотнения характерно для возбуждающих синапсов.
Срез дендрита проведен в поперечном направлении, о чем свидетельствует наличие множества микротрубочек.
Кроме того, видны некоторые нейрофиламенты. Участок синапса окружен протоплазматическим астроцитом. 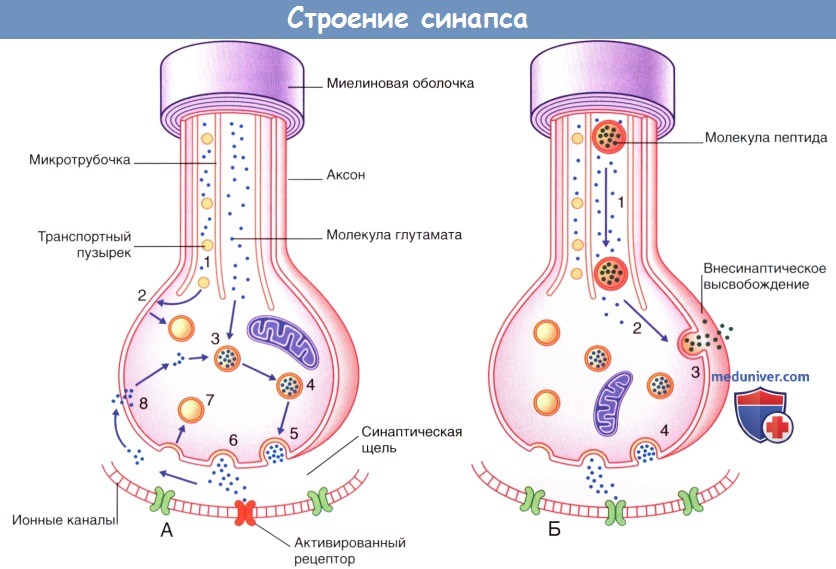 Процессы, происходящие в нервных окончаниях двух типов.
Процессы, происходящие в нервных окончаниях двух типов.
(А) Синаптическая передача небольших молекул (например, глутамата).
(1) Транспортные пузырьки, содержащие мембранные белки синаптических пузырьков, направляются вдоль микротрубочек к плазматической мембране булавовидного утолщения.
В это же время происходит перенос молекул ферментов и глутамата путем медленного транспорта.
(2) Мембранные белки пузырьков выходят из плазматической мембраны и формируют синаптические пузырьки.
(3) Глутамат погружается в синаптические пузырьки; происходит накопление медиатора.
(4) Пузырьки, содержащие глутамат, подходят к пресинаптической мембране.
(5) В результате деполяризации происходит экзоцитоз медиатора из частично разрушенных пузырьков.
(6) Высвобождающийся медиатор распространяется диффузно в области синаптической щели и активирует специфические рецепторы на постсинаптической мембране.
(7) Мембраны синаптических пузырьков транспортируются обратно в клетку путем эндоцитоза.
(8) Происходит частичный обратный захват глутамата в клетку для повторного использования.
(Б) Передача нейропептидов (например, субстанции Р), осуществляющаяся одновременно с синаптической передачей (например, глутамата).
Совместная передача этих веществ происходит в центральных нервных окончаниях униполярных нейронов, обеспечивающих болевую чувствительность.
(1) Синтезированные в комплексе Гольджи (в области перикариона) пузырьки и предшественники пептидов (пропептиды) транспортируются к булавовидному расширению путем быстрого транспорта.
(2) При их попадании в область булавовидного утолщения завершается процесс формирования молекулы пептида, и пузырьки транспортируются к плазматической мембране.
(3) Деполяризация мембраны и перенос содержимого пузырьков в межклеточное пространство путем экзоцитоза.
(4) Одновременно с этим происходит высвобождение глутамата.
1. Активация рецепторов. Молекулы медиаторов проходят через синаптическую щель и активируют рецепторные белки, расположенные парами на постсинаптической мембране. Активация рецепторов запускает ионные процессы, которые приводят к деполяризации постсинаптической мембраны (возбуждающее постсинаптическое действие) или гиперполяризации постсинаптической мембраны (тормозящее постсинаптическое действие). Изменение электротонуса передается в сому в виде затухающего по мере распространения электротонического потенциала, за счет которого происходит изменение потенциала покоя в начальном сегменте аксона.
Ионные процессы подробно описаны в отдельной статье на сайте. При преобладании возбуждающих постсинаптических потенциалов начальный сегмент аксона деполяризуется до порогового уровня и генерирует потенциал действия.
Наиболее распространенный возбуждающий медиатор ЦНС — глутамат, а тормозной — гамма-аминомасляная кислота (ГАМК). В периферической нервной системе медиатором для двигательных нейронов поперечно-полосатой мускулатуры служит ацетилхолин, а для чувствительных нейронов — глутамат.
Последовательность процессов, происходящих в глутаматергических синапсах, показана на рисунке ниже. При передаче глутамата совместно с другими пептидами высвобождение пептидов осуществляется внесинаптическим путем.
Большинство чувствительных нейронов помимо глутамата выделяет и другие пептиды (один или несколько), высвобождающиеся в различных участках нейрона; однако основная функция этих пептидов — модуляция (повышение или снижение) эффективности синаптической передачи глутамата.
Кроме того, нейротрансмиссия может происходить путем диффузной внесинаптической передачи сигнала, характерной для моноаминергических нейронов (нейронов, использующих биогенные амины для обеспечения нейротрансмиссии). Выделяют две разновидности моноаминергических нейронов. В одних нейронах осуществляется синтез катехоламинов (норадреналина или дофамина) из аминокислоты тирозина, а в других — серотонина из аминокислоты триптофана. Например, дофамин высвобождается как в синаптической области, так и из варикозных утолщений аксона, в которых также происходит синтез этого нейромедиатора.
Дофамин проникает в межклеточную жидкость ЦНС и до момента деградации способен активировать специфические рецепторы на расстоянии до 100 мкм. Моноаминергические нейроны присутствуют во многих структурах ЦНС; нарушение передачи импульса этими нейронами приводит к различным заболеваниям, среди которых выделяют болезнь Паркинсона, шизофрению и глубокую депрессию.
Оксид азота (газообразная молекула) также участвует в диффузной нейропередаче в глутаматергической системе нейронов. Избыточное влияние оксида азота оказывает цитотоксическое действие, особенно в тех участках, кровоснабжение которых нарушено за счет тромбоза артерий. Глутамат также является потенциально цитотоксическим нейромедиатором.
В отличие от диффузной нейротрансмиссии, традиционную синаптическую передачу сигнала ввиду ее относительной стабильности называют «проводниковой».
в) Резюме. Мультиполярные нейроны ЦНС состоят из сомы, дендритов и аксона; аксон образует коллатеральные и терминальные ветви. В соме расположены гладкая и шероховатая эндоплазматическая сети, комплексы Гольджи, нейрофиламенты и микротрубочки. Микротрубочки пронизывают нейрон на всем протяжении, принимают участие в процессе антероградного транспорта синаптических пузырьков, митохондрий и веществ для построения мембран, а также обеспечивают ретроградный транспорт «маркерных» молекул и разрушенных органелл.
Существует три вида химических межнейрональных взаимодействий: синаптическое (например, глутаматергическое), внесинаптическое (пептидергическое) и диффузное (например, моноаминергическое, серотонинергическое).
Химические синапсы классифицируют по анатомическому строению на аксодендритические, аксосоматические, аксоаксональные и дендро-дендритические. Синапс представлен пре- и постсинаптическими мембранами, синаптической щелью и субсинаптической активной зоной.
Электрические синапсы обеспечивают одновременную активацию целых групп нейронов, образуя между ними электрические связи за счет щелевидных контактов (нексусов).
![]() Диффузная нейротрансмиссия в головном мозге.
Диффузная нейротрансмиссия в головном мозге.
Аксоны глутаматергического (1) и дофаминергического (2) нейронов образуют плотные синаптические контакты с отростком звездчатого нейрона (3) полосатого тела.
Дофамин высвобождается не только из пресинаптической области, но и из варикозного утолщения аксона, откуда диффузно распространяется в межклеточное пространство и активирует дофаминовые рецепторы дендритного ствола и стенки перицита капилляра. 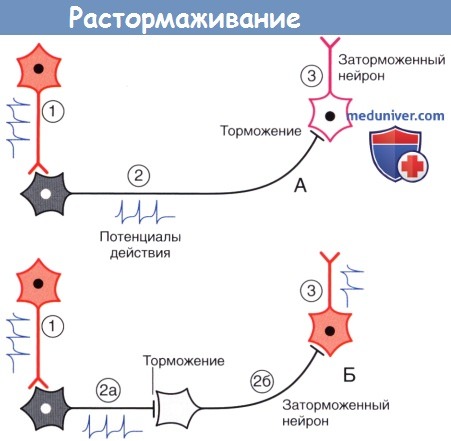 Растормаживание.
Растормаживание.
(А) Возбуждающий нейрон 1 активирует тормозной нейрон 2, который в свою очередь затормаживает нейрон 3.
(Б) Появление второго тормозного нейрона (2б) оказывает противоположное влияние на нейрон 3, поскольку происходит торможение нейрона 2б.
Спонтанно-активный нейрон 3 генерирует сигналы в условиях отсутствия тормозных влияний.
3. Торможение и растормаживание. Функционирование спонтанно-активных нейронов сдерживается под влиянием тормозных нейронов (обычно, ГАМКергических). Деятельность тормозных нейронов, в свою очередь, может быть ингибирована воздействующими на них другими тормозными нейронами, в результате чего происходит растормаживание клетки-мишени. Процесс растормаживания — важная особенность нейрональной активности в базальных ганглиях.
4. Редкие виды химических синапсов. Выделяют два типа аксоаксональных синапсов. В обоих случаях булавовидное утолщение образует тормозной нейрон. Синапсы первого типа образуются в области начального сегмента аксона и передают мощное ингибирующее влияние тормозного нейрона. Синапсы второго типа образуются между булавовидным утолщением тормозного нейрона и булавовидными утолщениями возбуждающих нейронов, что приводит к угнетению высвобождения медиаторов. Этот процесс получил название пресинаптического торможения. В этом плане традиционный синапс обеспечивает постсинаптичсекое торможение.
Дендро-дендритические (Д-Д) синапсы образуются между дендритными шипиками дендритов смежных шипиковых нейронов. Их задача — не генерирование нервного импульса, а изменение электротонуса клетки-мишени. В последовательных Д-Д-синапсах синаптические пузырьки располагаются только в одном дендритном шипике, а в реципрокном Д-Д-синапсе— в обоих. Возбуждающие Д-Д-синапсы изображены на рисунке ниже. Тормозные Д-Д-синапсы широко представлены в переключающих ядрах таламуса.
Кроме того, выделяют немногочисленные сомато-дендритические и сомато-соматические синапсы.
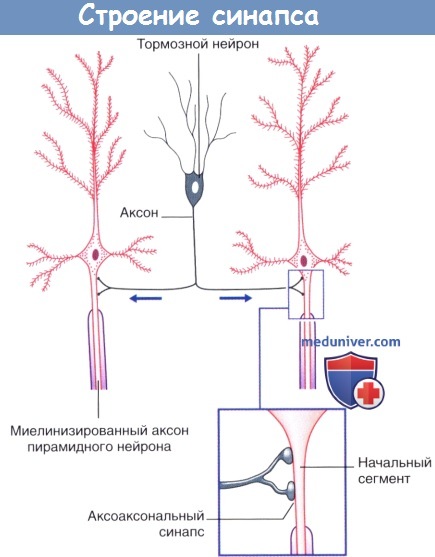 Аксоаксональные синапсы коры головного мозга.
Аксоаксональные синапсы коры головного мозга.
Стрелками указано направление проведения импульсов. 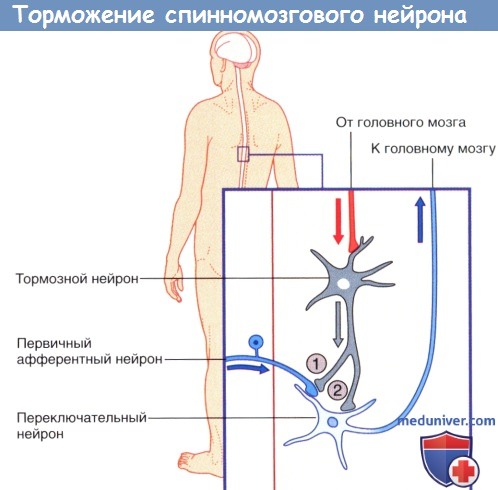 (1) Пресинаптическое и (2) постсинаптическое торможение спинномозгового нейрона, направляющегося к головному мозгу.
(1) Пресинаптическое и (2) постсинаптическое торможение спинномозгового нейрона, направляющегося к головному мозгу.
Стрелками указано направление проведения импульсов (возможно торможение переключательного нейрона под действием тормозных влияний). 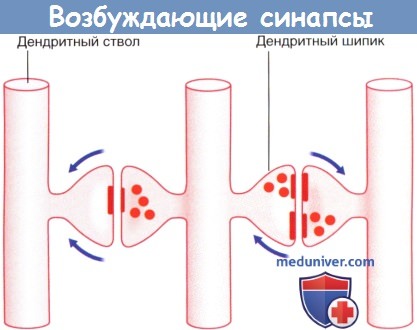 Возбуждающие дендро-дендритические синапсы. Изображены дендриты трех нейронов.
Возбуждающие дендро-дендритические синапсы. Изображены дендриты трех нейронов.
Реципрокный синапс (справа). Стрелками указано направление распространения электрото-нических волн.
Редактор: Искандер Милевски. Дата публикации: 11.11.2018
Что такое постсинаптическая мембрана
Открытие калиевых каналов позволяет положительно заряженным ионам К+ двигаться наружу, что приводит к большей отрицательности внутри клетки, чем в покое. Таким образом, оба события (вход ионов Сl- в клетку и выход ионов К+ из нее) увеличивают степень внутриклеточной отрицательности. Этот процесс называют гиперполяризацией. Увеличение отрицательности мембранного потенциала по сравнению с его внутриклеточным уровнем в покое тормозит нейрон, поэтому выход значений отрицательности за пределы исходного мембранного потенциала покоя называют тормозным постсинаптическим потенциалом (ТПСП).
б) Пресинаптическое торможение. Кроме постсинаптического торможения, вызываемого тормозными синапсами, действующими на мембране нейрона, часто происходит торможение другого типа, которое развивается на пресинаптических терминалях, прежде чем сигнал достигает синапса. Этот тип торможения, называемый пресинаптическим торможением, осуществляется следующим путем.
Причиной пресинаптического торможения является выделение тормозного медиатора на наружную поверхность пресинаптических нервных волокон, прежде чем их собственные окончания достигнут поверхности постсинаптического нейрона. В большинстве случаев тормозным медиатором является ГАМК. При этом развивается специфический эффект, связанный с открытием анионных каналов, что позволяет большому числу ионов СГ диффундировать в терминальное волокно. Отрицательные заряды этих ионов тормозят синаптическое проведение, нейтрализуя большую часть возбуждающего эффекта положительно заряженных ионов Na+, которые также входят в терминальные волокна, когда здесь возникает потенциал действия.
Пресинаптическое торможение происходит во многих сенсорных путях нервной системы. Смежные сенсорные волокна часто взаимно тормозят друг друга, что сводит к минимуму боковое распространение и смешивание сигналов в чувствительных трактах.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Постсинаптическая мембрана
Смотреть что такое «Постсинаптическая мембрана» в других словарях:
постсинаптическая мембрана — (membrana postsynaptica, LNH; пост + синапс) утолщенная поверхностная мембрана клетки в области синапса, обладающая чувствительностью к медиатору … Большой медицинский словарь
Синапс — I Синапс (греч. synapsis соприкосновение, соединение) специализированная зона контакта между отростками нервных клеток и другими возбудимыми и невозбудимыми клетками, обеспечивающая передачу информационного сигнала. Морфологически С. образован… … Медицинская энциклопедия
Химический синапс — Химический синапс особый тип межклеточного контакта между нейроном и клеткой мишенью. У данного типа синапса роль посредника (медиатора) передачи выполняет химическое вещество. Состоит из трёх основных частей: нервного окончания с… … Википедия
Возбуждение — (биологическое) реакция живой клетки на раздражение, выработанная в процессе эволюции. При В. живая система переходит из состояния относительного физиологического покоя к деятельности (например, сокращение мышечного волокна, выделение… … Большая советская энциклопедия
Нервно-мышечный синапс — 1. Пресинаптическое окончание 2. Сарколемма 3. Синаптический пузырек 4. Никотиновый ацетилхолиновый рецептор 5. Митохондрия Нервно мышечный синапс (также … Википедия
Мионевральный синапс — Нервно мышечный синапс (мионевральный синапс) эффекторное нервное окончание на скелетном мышечном волокне. Нервный отросток проходя через сарколемму мышечного волокна утрачивает миелиновую оболочку и образует сложный аппарат с цитолеммой… … Википедия
Нервно-мышечное окончание — Нервно мышечный синапс (мионевральный синапс) эффекторное нервное окончание на скелетном мышечном волокне. Нервный отросток проходя через сарколемму мышечного волокна утрачивает миелиновую оболочку и образует сложный аппарат с цитолеммой… … Википедия
ВОЗБУЖДЕНИЕ — возбуждение, совокупность физических, физико химических и функциональных изменений, возникающих в возбудимых тканях (нервной, мышечной) под влиянием раздражения. Способность клеток и тканей реагировать на раздражение процессом В. называется… … Ветеринарный энциклопедический словарь
Нервная система — совокупность структур в организме животных и человека, объединяющая деятельность всех органов и систем и обеспечивающая функционирование организма как единого целого в его постоянном взаимодействии с внешней средой. Н. с. воспринимает… … Большая советская энциклопедия
Холинэстераза — См. также: Ацетилхолинэстераза Трёхмерная модель молекулы ацетилхолинэстеразы … Википедия



