Научная электронная библиотека

Шевченко Б П, Гончаров А Г, Сеитов М С,
2.5. Гомеостаз
Гомеостаз (гр. homeo – подобный, stasis – состояние) – обозначает постоянство состава внутренней среды и некоторых функций организма (кровообращения, обмена веществ, терморегуляции и т.д.). Таким образом, термин «гомеостаз» – это не просто химическое постоянство среды или физиологических свойств организма, а особая устойчивость организма в пределах «нормы». Выход колебаний среды за пределы «нормы» ведет к патологии.
Гомеостаз по существу представляет собой эволюционное наследство адаптивных свойств организма к обычным условиям внешней среды, но эти условия могут кратковременно или долговременно выходить за пределы нормы обычных условий внешней среды. Тогда явления адаптации не только восстанавливают обычные свойства внутренней среды организма, но и кратковременно изменяют активность органов, систем (учащается пульс, дыхание, повышается температура и т.д.). В итоге организм адаптируется к внешней и внутренней средам.
В ветеринарной практике под адаптацией обычно понимают приспособление организма к измененным условиям существования. Однако следует помнить, что адаптируется животное на основе механизмов гомеостаза.
В плодном периоде развития домашних коз механизмы гомеостаза почти отсутствуют, они появляются и развиваются только в постнатальном периоде и к старости ослабевают.
Непосредственно в формировании внутренней среды организма принимает участие сердечно-сосудистая система. Кровь, наряду с другими функциями, выполняет гомеостатическую функцию, доставляет к клеткам, тканям гормоны и химические вещества, таким образом, принимает участие в формировании гомеостаза (А.В. Вальдман, 1981). От химического состава внутренней среды зависит реактивность, возбудимость как отдельных органов, систем, так всего организма.
И так гомеостаз – это большая проблема для патологов, так как означает не только сохранение постоянства внутренней среды, но и ее восстановление, и приспособление к внешней среде. Сама, в сущности, болезнь представляет проблему гомеостаза, нарушение его механизмов и пути восстановления. Однако решение многих проблем гомеостаза, особенно, организма коз, еще впереди.
Клиническая фармакология Тимогена®. Глава 1
Долгов Г.В., Куликов С.В., Легеза В.И., Малинин В.В., Морозов В.Г., Смирнов В.С., Сосюкин А.Е.
Под редакцией проф. В.С. Смирнова.
Авторский коллектив:
Опубликовано в СПб, 2003. — 103 с.
Скачать PDF
Введение
Глава 1. Механизмы пептидной регуляции гомеостаза (В.В. Малинин, В.Г. Морозов)
Глава 2. Регуляторные пептиды тимуса (В.С. Смирнов)
Глава 3. Тимоген®: структура, химический синтез, свойства (С.В. Куликов, В.С. Смирнов)
Глава 4. Тимоген® в профилактике и комплексной терапии инфекционных заболеваний (В.С. Смирнов)
Глава 5. Тимоген® в терапии бронхолегочных заболеваний (В.С. Смирнов)
Глава 6. Применение тимогена в комплексной терапии внутренних болезней (В.С. Смирнов, А.Е. Сосюкин)
Глава 7. Тимоген® в дерматологии (В.С. Смирнов)
Глава 8. Применение Тимогена® для профилактики и лечения радиационных поражений (В.И.Легеза, В.С. Смирнов)
Глава 9. Применение Тимоген® в комплексном лечении механических и термических травм (В.С. Смирнов)
Глава 10. Тимоген® в акушерско-гинекологической практике (Г.В. Долгов, В.С. Смирнов)
Глава 11. Особенности применения Тимогена® в педиатрии (В.С. Смирнов)
Заключение
Глава 1. МЕХАНИЗМЫ ПЕПТИДНОЙ РЕГУЛЯЦИИ ГОМЕОСТАЗА
В.В. Малинин, В.Г. Морозов
Механизмы, регулирующие постоянство внутренней среды (гомеостаз), представляют собой сложный комплекс нейрогуморальных процессов, позволяющих организму сохранять жизнеспособность и устойчивость в окружающей среде. При этом стабильность внутренней среды тесно связана с уровнем биологической защиты организма.
По современным представлениям регуляция гомеостаза многоклеточных систем осуществляется с помощью нейроэндокринных, иммунологических, клеточных и молекулярных механизмов. Наиболее изучена роль нервных и гормональных воздействий на процессы, позволяющие организму контролировать постоянство внутренней среды (Горизонтов, 1981). Функция иммунной системы рассматривается как висцеральная, обеспечивающая сохранение генетического постоянства клеточного состава, т.е. она является одним из гомеостатических механизмов целостного организма (Корнева, 1993).
Известно, что нервная и эндокринная системы модулируют функции иммунной системы с помощью нейротрансмиттеров, нейропептидов и гормонов, а иммунная система взаимодействует с нейроэндокринной системой с помощью цитокинов, иммунопептидов и других иммунотрансмиттеров. В настоящее время установлена роль эндогенных пептидов в формировании компенсаторно-приспособительных реакций организма в ответ на стресс и нарушения гомеостаза. Система пептидов рассматривается в качестве универсальной при нейроиммуноэндокринных взаимодействиях (Коpнева, Шхинек, 1988; Fabry et al., 1994).
При изучении механизма действия цитомединов было установлено, что эти факторы принимают непосредственное участие в процессах тканеспецифической регуляции экспрессии генов и биосинтеза. В результате этого в клетках понижается скорость накопления патологических изменений (повреждения ДНК, мутации, злокачественная трансформация и т.п.) и повышается активность репаративных процессов, направленных на восстановление клеточного гомеостаза.
Нарушение цитомединовой регуляции снижает резистентность клеток и тканей организма к дестабилизирующим факторам как внешней, так и внутренней среды. Это может служить одной из причин развития заболеваний, инволюции органов и тканей, а также ускоренного старения. Последующие работы подтвердили, что система пептидергической регуляции включает широкий спектр тканеспецифических пептидов, поддерживающих гомеостаз (Ivanov et al., 1997; Karelin et al., 1998).
В настоящее время цитомедины выделены практически из всех клеток, тканей и биологических жидкостей организма. По данным физико-химических исследований эти комплексы пептидов различаются между собой по составу, молекулярной массе и электрохимическим свойствам компонентов. На основе цитомединов разработан целый ряд новых лекарственных препаратов. Применение этих препаратов в условиях нарушенного клеточного гомеостаза позволяет восстанавливать функциональную активность различных физиологических систем организма (Морозов, Хавинсон, 1996).
Интенсивное исследование регуляторных пептидов за последние 2-3 десятилетия привело к кардинальному пересмотру представлений о механизмах регуляции физиологических функций, принципов координации процессов гомеостаза и адаптации функциональных систем организма к окружающей среде.
Оказалось, что для воздействия на физиологические процессы необязательно наличие целой молекулы. Более того, в некоторых случаях фрагменты, состоящие всего из 3-4 аминокислотных остатков, были эффективнее, чем нативные соединения. Эти данные послужили предпосылкой к формированию представлений о том, что регуляция и координирование функций организма могут осуществляться за счет процессинга полипептидов, когда в зависимости от потребностей организма от достаточно длинных полипептидных цепей отщепляются фрагменты, обладающие той или иной степенью активности, специфичности и направленности действия на определенные физиологические системы. Процессинговая регуляция обладает значительно большей степенью гибкости, позволяя в короткие сроки путем активации соответствующих пептидаз образовывать в нужном месте требуемые регуляторы из уже готового предшественника. Кроме того, в механизм процессинга заложена определенная программа последовательности включения регуляторов. Процессинговый тип регуляции в наибольшей степени присущ именно пептидным соединениям с линейной структурой, открывающей широкие возможности для изменения конформации молекулы при отщеплении хотя бы одного аминокислотного остатка с любого конца. Кроме того, при таком отщеплении могут значительно меняться другие свойства молекулы, например, степень ее гидрофобности, определяющая способность прохождения через клеточные мембраны и гистогематические барьеры и т.д. (Ерошенко и др., 1991).
Как известно, подавляющее большинство регуляторных пептидов обладает поли- функциональностью. Другими словами, одно соединение обеспечивает регуляцию различных, часто физиологически несхожих функций. В связи с этим, многие физиологические функции оказываются под контролем целого ряда регуляторных пептидов.
Все больше и больше данных свидетельствует о том, что регуляторные олигопептиды являются участниками процессов роста, развития и регенерации. Многие из них представляют собой хорошо изученные структуры, регулирующие различные физиологические функции организма (Замятнин, 1988). Предполагают, что на уровне олигопептидов существует единая система регуляции как эмбрионального развития, роста и регенерации, так и функционирования сформированного организма. По-видимому, в процессе морфогенеза большинство (если не все) функционально активные олигопептиды принимают участие в появлении новых форм и структур в ходе индивидуального развития. При этом становится очевидной условность подразделения олигопептидов на нейро-, эндокрино- или иммуноактивные и одновременно на морфогенетически активные факторы (Замятнин, 1992).
Известно, что взаимодействие лиганда с рецептором реализуется на основе их структурного соответствия (Говырин, Жоров, 1994). Для олигопептида это означает наличие определенной совокупности свойств молекулы, которую на основе применения принципов системного анализа и элементов теории информации было предложено называть сигнатурой (Чипенс, 1980). Очевидно, что это понятие включает сведения и об аминокислотной последовательности. Как отмечает Г.И. Чипенс, каждая пептидная молекула имеет бесконечное число свойств, которые проявляются и определяются только в процессе взаимодействия с другими молекулами как результат внутри- и межмолекулярных взаимодействий в условиях данной среды.
В основу анализа первичных структур белков и пептидов были положены три принципа теории информации — сигнатур, двузначности и эквивокации (Чипенс, 1980; Quastler, 1965). Согласно принципу сигнатур, взаимодействие и комплексообразование молекул определяется наборами свойств (сигнатурами) активных участков их электронных структур (носителей сигнатур). Поскольку молекула может иметь множество сигнатур, то это приводит к неопределенности биологических эффектов, которые она может индуцировать (принцип двузначности теории информации). В определенных ситуациях различные по своей химической структуре молекулы могут иметь одинаковые сигнатуры и выполнять одинаковые функции (принцип эквивокации, или неопределенности причины эффекта). На основе принципов сигнатур и эквивокации развиты представления об эквифункциональных, т.е. однонаправленно действующих, аминокислотных остатках. В зависимости от сигнатуры однонаправленно действующими могут быть самые разные по химической структуре аминокислотные остатки (Чипенс и др., 1990).
Таким образом, основываясь на представлениях о сигнатурах, а также принципах эквивокации (несколько структур ® одна сигнатура ® одна функция) и двузначности (одна структура ® несколько сигнатур ® несколько функций), можно попытаться дать общую характеристику функциональных особенностей эндогенных регуляторных олиго- пептидов, помогающую уяснить, почему структурно разные молекулы способны вызвать близкие, практически одинаковые реакции или почему одна молекулярная структура участвует в различных физиологических процессах? Очевидно, что на основании принципа двузначности несостоятельность существующих понятий и терминов может быть объяснена наличием нескольких сигнатур у одной молекулы олигопептида, что позволяет ей взаимодействовать с рецепторами нескольких типов.
Выявление двух типов функционально значимых групп – положительно заряженных и циклических (R+ и cyc), позволяет рассматривать одно из свойств сигнатуры как взаимное расположение этих групп в первичной структуре олигопептида. Исходя из этого, можно представить значительное число структур, содержащих одинаковое расположение радикалов R+ и сyc, в то время как сами эти радикалы в разных молекулах будут принадлежать аминокислотным остаткам разного типа. Хорошо известными примерами такого рода среди радикалов R+ являются взаимные замены остатков Arg и Lys у членов одного олигопептидного семейства. Более того, многочисленные возможные замены других аминокислотных остатков при сохранении расположения R+ и cyc также могут приводить к одинаковой сигнатуре при разной первичной структуре. Примерами могут служить данные по сравнению первичных структур разных олигопептидов или по семействам, полученные в результате классификации, в том числе замены близких по радикалам аминокислотных остатков в квазиконсервативной области. По-видимому, в этом и проявляется принцип эквивокации (Zamyatnin, 1991).
Основу принципа двузначности составляет высокая конформационная подвижность олигопептидов, в результате которой одна молекула принимает различные конформации (имеет несколько сигнатур), и этим обеспечивается пространственное соответствие с рецепторами различного типа.
На основании исследований физико-химических особенностей эндогенных олигопептидов сделано предположение о том, что спектр функциональной активности этих веществ в основном определяется двумя типами радикалов, формирующих сигнатуру, а состав радикалов определяет полифункциональность олигопептидов. В то же время их уникальное распределение вдоль цепи молекулы (последовательность) определяет специфичность действия. Эти два типа радикалов в принципе могут составлять основу молекулярного физиологического кода. Данные выводы могут быть использованы при прогнозировании функциональных свойств олигопептидов, основанном на рассмотрении структуры. Кроме того, обнаружение ограниченного числа функционально значимых групп, по-видимому, позволит сузить поиск новых высокоактивных соединений пептидной природы (Замятнин, 1990).
Характерным признаком регуляторных олигопептидов оказалось небольшое содержание в них аминокислотных остатков с отрицательно заряженными боковыми радикалами (Аsp и Glu). В то же время содержание остатков с положительным зарядом достоверно больше только для Arg. Достаточно часто встречаются аминокислотные остатки Pro, Phe, Tyr, Trp и Cys. Большинство этих молекул содержат циклическую химическую группу (а Cys, как правило, участвует в образовании молекулярных макроциклов). Из сказанного следует, что регуляторные олигопептиды с заданным спектром функциональной активности содержат преимущественно положительно заряженные и циклические радикалы.
Сравнение аминокислотных последовательностей пептидных препаратов, выделенных из различных органов и тканей млекопитающих, не позволяет выявить в них гомологические участки. Если же сравнивать суммарный аминокислотный состав, то можно отметить высокое содержание аминокислот с боковыми амино- и карбоксильными группами, т. е. высокую диполярность этих макромолекул. Можно предположить, что именно высокоосновные боковые группы, как, например, у тафцина, обеспечивают селективное взаимодействие этих пептидов с поверхностными рецепторами клеток, которые содержат, как правило, карбоксильные группы глутаминовой, аспарагиновой и сиаловой кислот. Иначе говоря, в основе селективного аффинного взаимодействия пептидов-регуляторов с клеточной мембраной лежат ион-ионные и ион-дипольные взаимодействия пептида с карбоксильными группами мембраны (Демин и др., 1994).
Внутриклеточной мишенью для эндогенных биологически активных пептидов, вероятно, является биохимический комплекс, осуществляющий в клетке синтез белка. Получены первые экспериментальные данные, доказывающие целесообразность введения понятия об эссенциальных аминокислотах в качестве метаболической особенности каждого органа (ткани). Это дает основание предполагать, что изменение внутриклеточной концентрации данных аминокислот (их особого сочетания в виде ди- и трипептидов) играет важную роль в регуляции рибосомального синтеза белка. Говоря об уникальных особенностях метаболизма некоторых органов, имеется в виду тот факт, что некоторые клетки, например, кардиомиоциты, синтезируют новые белковые соединения в основном из аминокислот, высвобождающихся только при катаболизме собственных белков. Из притекающей крови кардиомиоциты утилизируют лишь две аминокислоты – Asp и Glu. Данное положение подсказывает решение вопроса об эссенциальных аминокислотах в составе пептидов, высокотропных для миокарда (Кожемяки, 1992).
Проблема биогенеза регуляторных олигопептидов из белковых предшественников вблизи клеточных рецепторов является кардинальной для изучения механизма действия ростовых трансформирующих факторов, нейроактивных пептидов и белков, белковых гормонов и др.
Как и для большинства физиологически активных веществ, эффект регуляторных пептидов определяется взаимодействием со специфическими рецепторами. Расчеты показывают, что в ряде случаев эффекты, связанные с пептидами, не удается объяснить с позиций лиганд-рецепторных взаимодействий. Отсюда возникло предположение об их модулирующем влиянии, которое сводится к изменению характеристик возбудимых мембран клетки (рецептора), облегчающих реализацию эффекта основного медиатора (Гомазков, 1992).
Регуляторные пептиды и сопряженные с их функцией ферменты следует рассматривать как сложную адаптивную систему организма, организующую реализацию приспособительных реакций на всех уровнях его интеграции. Возможно, разнообразные эффекты одного пептида объясняются не его непосредственным действием, а модуляцией эффектов нервной и гуморальной регуляции.
Особый интерес вызывает исследование процессов эндоцитоза (и не только лиганд- рецепторных комплексов). Отдельные участки поверхностных мембран клеток непрерывно втягиваются внутрь и отрываются, образуя внутриклеточные пузырьки, содержащие вещества, которые находились во внешней среде или были адсорбированы на поверхности клетки. Поэтому можно допустить возможность попадания пептидов и белков внутрь клеток и без наличия специфических для них рецепторов на клеточной поверхности.
В последнее время наблюдается возрастающий интерес исследователей к парадоксальным эффектам действия сверхмалых доз (10-18-10-14 М) биологически активных веществ. Данные эффекты наблюдаются для самых разных групп веществ – гормонов и регуляторных пептидов, а также некоторых веществ непептидной природы.
Результаты экспериментов с концентрациями веществ 10-19 М и ниже довольно противоречивы, а объяснение эффектов при концентрациях ниже 10-19 М требует привлечения таких понятий, как “активированная” вода, “память молекул” (Замятнин, 1992).
Одной из особенностей действия сверхмалых доз пептидов является наличие отчетливого эффекта, несмотря на то, что во многих случаях в объекте эксперимента присутствует значительно бoльшая эндогенная концентрация того же вещества. Предполагают, что эффекты сверхмалых доз связаны с адаптационными явлениями, поскольку клетка может реагировать не на величину действующей концентрации, а на изменения концентрации вещества в малых и сверхмалых дозах (Сазанов, Зайцев, 1992). Усиление сигнала возможно не только путем изменения концентраций вторичных мессенджеров, но также и за счет активации синтеза белков, участвующих в передаче сигнала (Reibman et al., 1991). Предполагают, что для достижения эффекта достаточно того, чтобы до клеток достигли самые “быстрые” молекулы действующего вещества из общего распределения, а не все молекулы (Бурлакова и др., 1990).
Выделяют несколько основных систем, необходимых для реализации эффектов сверхмалых концентраций (доз) эндогенных и экзогенных веществ: а) системы каскадные, амплифицирующие сигнал; б) собирательные, “отлавливающие” системы; в) накопители и транспортеры сигнальных молекул; г) супераффинные рецепторы (Ашмарин и др., 1996).
Физико-химическую основу феномена высокой чувствительности организма к так называемым факторам малой интенсивности, в том числе, и сверхмалым дозам биологически активных веществ, составляют процессы колебания биомолекул, перехода одного типа энергии в другой, резонансные эффекты взаимодействия и некоторые иные механизмы. При этом с кибернетических позиций возможен перевод сложной системы на иной уровень реагирования (например, выход из патологического состояния) путем информационных воздействий на нее, которые в своей основе имеют характер слабых по силе сигналов (Подколзин и др., 1994).
Как было отмечено, пептидная регуляция осуществляет связь между нервной, эндокринной, иммунной и, по-видимому, другими системами, участвующими в поддержании гомеостаза. Полифункциональность пептидов и каскадный механизм реализации биологических эффектов определяют те процессы, которые происходят в организме как после экзогенного введения пептида, так и после его эндогенного образования.
Таким образом, пептидная биорегуляция – новое научное направление, связанное с изучением молекулярных и клеточных механизмов, управляющих гомеостазом, разработкой способов и средств восстановления физиологических функций организма с целью предупреждения и лечения заболеваний. Дальнейшее развитие этого направления позволит по-новому подойти к изучению функций организма в норме и патологии, а также объяснить механизмы регуляции гомеостаза на уровне клеток и макромолекул.
Что такое регуляция гомеостаза
Кислотно-основное состояние (КОС) организма является одним из важнейших и наиболее строго стабилизируемых параметров гомеостаза. От соотношения водородных и гидроксильных ионов во внутренней среде организма зависят активность ферментов, гормонов, интенсивность и направленность окислительно-восстановительных реакций, процессы обмена белков, углеводов и жиров, функции различных органов и систем, постоянство водного и электролитного обмена, проницаемость и возбудимость биологических мембран и т.д. Активность реакции среды влияет на способность гемоглобина связывать кислород и отдавать его тканям.
Активную реакцию среды принято оценивать по содержанию в жидкостях ионов водорода.
Величина рН является одним из самых «жестких» параметров крови и колеблется у человека в норме в очень узких пределах – рН артериальной крови составляет 7,35–7,45; венозной – 7,32–7,42. Более значительные изменения рН крови связаны с патологическими нарушениями обмена. В других биологических жидкостях и в клетках рН может отличаться от рН крови.
Сдвиги рН крови за указанные границы приводят к существенным сдвигам окислительно-восстановительных процессов, изменению активности ферментов, прницаемости биологических мембран, обусловливают нарушения со стороны функции сердечно-сосудистой, дыхательной и других систем; сдвиг на 0,3 может вызвать коматозные состояния, а на 0,4 – зачастую несовместим с жизнью.
Кислотно-основное состояние поддерживается мощными гомеостатическими механизмами. В их основе лежат особенности физико-химических свойств буферных систем крови и физиологические процессы, в которых принимают участие системы внешнего дыхания, почки, печень, желудочно-кишечный тракт и др.
Химические буферные системы образуют первую линию защиты против изменений рН жидкости организма, действуют для быстрого их предотвращения.
Буферной системой называют смеси, которые обладают способностью препятствовать изменению рН среды при внесении в нее кислот или оснований. Буферные системы не удаляют H+ из организма, а «связывают» его своим щелочным компонентом до окончательного восстановления КОС. Буферными свойствами обладают смеси, которые состоят из слабой кислоты и ее соли, содержащей сильное основание, или из слабого основания и соли сильной кислоты.
Наиболее емкими буферными системами крови являются бикарбонатный, фосфатный, белковый и гемоглобиновый. Первые три системы особенно важную роль играют в плазме крови, а гемоглобиновый буфер, самый мощный, действует в эритроцитах.
Бикарбонатный буфер является наиболее важной внеклеточной буферной системой и состоит из слабой угольной кислоты Н2СО3 и соли ее аниона – сильного основания 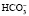 . Угольная кислота образуется в результате взаимодействия углекислого газа и воды: CO2 + H2O ↔ H2CO3. Угольная кислота в свою очередь диссоциирует на водород и бикарбонат: H2CO3 ↔ H+ + HCO3-.
. Угольная кислота образуется в результате взаимодействия углекислого газа и воды: CO2 + H2O ↔ H2CO3. Угольная кислота в свою очередь диссоциирует на водород и бикарбонат: H2CO3 ↔ H+ + HCO3-.
В нормальных условиях (при рН крови около 7,4) в плазме бикарбоната в 20 раз больше, чем углекислоты.
Емкость бикарбонатной системы составляет 53 % всей буферной емкости крови. При этом на бикарбонат плазмы приходится 35 % и на бикарбонат эритроцитов 18 % буферной емкости.
При образовании в плазме избытка кислореагирующих продуктов ионы водорода соединяются с анионами бикарбоната (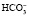 ). Образующийся при этом в плазме избыток углекислоты поступает в эритроциты и там с помощью угольной ангидразы разлагается на углекислый газ и воду. Углекислый газ выделяется в плазму, возбуждает дыхательный центр и избыток СО2 удаляется из организма через легкие. Это быстрое преобразование бикарбонатом любой кислоты в угольную, которая легко удаляется легкими, делает бикарбонатный буфер самой лабильной буферной системой.
). Образующийся при этом в плазме избыток углекислоты поступает в эритроциты и там с помощью угольной ангидразы разлагается на углекислый газ и воду. Углекислый газ выделяется в плазму, возбуждает дыхательный центр и избыток СО2 удаляется из организма через легкие. Это быстрое преобразование бикарбонатом любой кислоты в угольную, которая легко удаляется легкими, делает бикарбонатный буфер самой лабильной буферной системой.
Бикарбонатный буфер способен нейтрализовать и избыток оснований. В этом случае ионы ОНˉ будут связаны углекислотой и вместо самого сильного основания ОНˉ образуется менее сильное 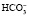 , избыток которого в виде бикарбонатных солей выделяется почками.
, избыток которого в виде бикарбонатных солей выделяется почками.
До тех пор, пока количество угольной кислоты и бикарбоната натрия изменяется пропорционально и соотношение между ними сохраняется 1:20, рН крови остается в пределах нормы.
Фосфатный буфер представлен солями одно- и двузамещенных фосфатов. Фосфатная буферная система обеспечивает 5 % буферной емкости крови, является основной буферной системой клеток.
Однозамещенная соль обладает кислыми свойствами, так как при диссоциации дает ион 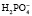 , который далее способен выделять ион водорода: NаН2РО4 ⇒ Nа+ +
, который далее способен выделять ион водорода: NаН2РО4 ⇒ Nа+ + 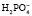 ;
; 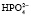 ⇒Н+ +
⇒Н+ + 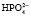 . Двузамещенный фосфат обладает свойствами основания, так как диссоциирует с образованием иона
. Двузамещенный фосфат обладает свойствами основания, так как диссоциирует с образованием иона 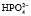 , который может связывать ион водорода:
, который может связывать ион водорода: 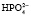 + Н+ ⇒
+ Н+ ⇒ 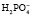 .
.
При нормальном рН в плазме соотношение фосфатных солей NаН2РО4: Nа2НРО4 = 1:4. Этот буфер имеет значение в почечной регуляции КОС, а также в регуляции реакции некоторых тканей. В крови же его действие главным образом сводится к поддержанию постоянства и воспроизводства бикарбонатного буфера.
Белковая буферная система является довольно мощным буфером, который способен проявлять свои свойства за счёт амфотерности белков. Белковая буферная система обеспечивает 7 % буферной емкости крови. Белки плазмы крови содержат достаточное количество кислых и основных радикалов, поэтому эта буферная система действует в зависимости от среды, в которой происходит диссоциация белков.
Гемоглобиновый буфер является самой емкой буферной системой. На ее долю приходится до 75 % всей буферной емкости крови. Свойства буферной системы гемоглобину придает главным образом его способность постоянно находиться в виде двух форм – восстановленного (редуцированного) гемоглобина ННb и окисленного (оксигемоглобина) НbО2.
Гемоглобиновый буфер, в отличие от бикарбонатного, в состоянии нейтрализовать как нелетучие, так и летучие кислоты. Окисленный гемоглобин ведёт себя как кислота, увеличивая концентрацию ионов водорода, а восстановленный (дезоксигенированный) – как основание, нейтрализуя H+.
Гемоглобин является классическим примером белкового буфера и эффективность его достаточно высока. Гемоглобин в шесть раз более эффективен как буфер, чем плазменные протеины.
Переход окисленной формы гемоглобина в восстановленную форму предупреждает сдвиг рН в кислую сторону во время контакта крови с тканями, а образование оксигемоглобина в легочных капиллярах предотвращает сдвиг рН в щелочную сторону за счет выхода из эритроцитов СО2 и иона хлора и образования в них бикарбоната.
Система аммиак/ион аммония (NH3/NH4+) – действует преимущественно в моче.
Помимо буферных систем в поддержании постоянства рН активное участие принимают физиологические системы, среди которых основными являются легкие, почки, печень, желудочно-кишечный тракт.
Система дыхания играет значительную роль в поддержании кислотно-щелочного баланса организма, однако для нивелирования сдвига рН крови им требуется 1–3 минуты. Роль легких сводится к поддержанию нормальной концентрации углекислоты, и основным показателем функционального состояния легких является парциальное напряжение углекислого газа в крови. Легочные механизмы обеспечивают временную компенсацию, так как при этом происходит смещение кривой диссоциации оксигемоглобина влево и уменьшается кислородная емкость артериальной крови.
При устойчивом состоянии газообмена легкие выводят углекислого газа около 850 г в сутки. Если напряжение углекислого газа в крови повышается сверх нормы на 10 мм рт. ст., вентиляция увеличивается в 4 раза.
Роль почек в регуляции активной реакции крови не менее важна, чем деятельность дыхательной системы. Почечный механизм компенсации более медленный, чем респираторный. Полноценная почечная компенсация развивается только через несколько дней после изменения pH.
Экскреция кислот при обычной смешанной пище у здорового человека превышает выделение оснований, поэтому моча имеет кислую реакцию (рН 5,3–6,5) и концентрация в ней ионов водорода примерно в 800 раз выше, чем в крови. Почки вырабатывают и выделяют с мочой количество ионов водорода, эквивалентное их количеству, непрерывно поступающему в плазму из клеток организма, совершая при этом замену ионов водорода, секретируемых эпителием канальцев, на ионы натрия первичной мочи. Этот механизм осуществляется с помощью нескольких химических процессов.
Первым из них является процесс реабсорбции натрия при превращении двузамещенных фосфатов в однозамещенные. При истощении фосфатного буфера (при рН мочи ниже 4,5) реабсорбция натрия и бикарбоната осуществляется за счет аммониогенеза.
Второй процесс, который обеспечивает задержку натрия в организме и выведение излишка ионов водорода, – это превращение в просвете канальцев бикарбонатов в угольную кислоту.
Третьим процессом, который способствует сохранению натрия в организме, является синтез в дистальных почечных канальцах аммиака (аммониогенез) и использование его для нейтрализации и выведения кислых эквивалентов с мочой.
Образовавшийся свободный аммиак легко проникает в просвет канальцев, где, соединяясь с ионом водорода, превращается в плохо диффундирующий аммонийный катион 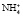 , не способный вновь вернуться в клетку стенки канальца.
, не способный вновь вернуться в клетку стенки канальца.
В общем итоге концентрация водородных ионов в моче может превышать концентрацию водородных ионов в крови в несколько сотен раз.
Это свидетельствует об огромной способности почек выводить из организма ионы водорода.
Почечные механизмы регуляции КОС не могут скорректировать рН в течение нескольких минут, как респираторный механизм, но они функционируют в течение нескольких дней, пока рН не вернется к нормальному уровню.
Регуляция КОС с участием печени. Печень окисляет до конечных продуктов недоокисленные вещества крови, оттекающей от кишечника; синтезирует мочевину из азотистых шлаков, в частности из аммиака и из хлорида аммония, поступающих из желудочно-кишечного тракта в кровь портальной вены; печени присуща выделительная функция и поэтому при накоплении в организме избыточного количества кислых или щелочных продуктов метаболизма они могут выделяться с желчью в желудочно-кишечный тракт. При избытке кислот в печени усиливается их нейтрализация и одновременно тормозится образование мочевины. Неиспользованный аммиак нейтрализует кислоты и увеличивает выведение аммонийных солей с мочой. При возрастании количества щелочных валентностей мочекинообразование возрастает, а аммониогенез снижается, что сопровождается уменьшением выведения с мочой аммонийных солей.
Концентрация водородных ионов в крови зависит также от деятельности желудка и кишечника. Клетки слизистой желудка секретируют соляную кислоту в очень высокой концентрации. При этом из крови ионы хлора выделяются в полость желудка в соединении с ионами водорода, образующимися в эпителии желудка с участием карбоангидразы. Взамен хлоридов в плазму в процессе желудочной секреции поступает бикарбонат.
Поджелудочная железа активно участвует в регуляции рН крови, так как она генерирует большое количество бикарбоната. Образование бикарбоната тормозится при избытке кислот и усиливается при их недостатке.
Кожа может в условиях избытка нелетучих кислот и оснований выделять последние с потом. Это имеет особое значение при нарушении функции почек.
Костная ткань. Это наиболее медленно реагирующая система. Механизм ее участия в регуляции рН крови состоит в возможности обмениваться с плазмой крови ионами Са2+ и Na+ в обмен на протоны Н+. Происходит растворение гидроксиапатитных кальциевых солей костного матрикса, освобождение ионов Са2+ и связывание ионов НРО42– с Н+ с образованием дигидрофосфата, который уходит с мочой. Параллельно при снижении рН (закисление) происходит поступление ионов H+ внутрь остеоцитов, а ионов калия – наружу.
Оценка кислотно-основного состояния организма
При изучении кислотно-щелочного баланса наибольшее значение имеет исследование крови. Показатели в капиллярной крови близки к показателям артериальной. В настоящее время показатели КОС определяют эквилибрационным микрометодом Аструпа. Данная методика позволяет, помимо истинного рН крови, получить показатель напряжения СО2 в плазме (рСО2), истинный бикарбонат крови (АВ), стандартный бикарбонат (SB), сумму всех оснований крови (ВВ) и показатель дефицита или избытка оснований (ВЕ).



