Синаптонемный комплекс

СОДЕРЖАНИЕ
Состав [ править ]
У человека были охарактеризованы три специфических компонента синаптонемного комплекса: SC-белок-1 (SYCP1), SC-белок-2 (SYCP2) и SC-белок-3 ( SYCP3 ). SYCP1 гена на хромосоме 1p13; ген SYCP2 находится на хромосоме 20q13.33; а ген SYCP3 находится на хромосоме 12q. [3]
Синаптонемный комплекс был описан Монтроузом Дж. Мозесом в 1956 году для первичных сперматоцитов раков и Д. Фосеттом в сперматоцитах голубя, кошки и человека. [4] Как видно под электронным микроскопом, синаптонемный комплекс образован двумя «боковыми элементами», в основном образованными SYCP3 и, во вторую очередь, SYCP2, «центральным элементом», который содержит по крайней мере два дополнительных белка и аминоконцевую область SYCP1. и «центральная область», расположенная между двумя боковыми элементами, которая содержит «поперечные филаменты», состоящие в основном из белка SYCP1. [3]
SC можно увидеть с помощью светового микроскопа с использованием окрашивания серебром или с помощью методов иммунофлуоресценции, которые маркируют белки SYCP3 или SYCP2.
Сборка и разборка [ править ]
В процессе развития клетки синаптонемный комплекс исчезает на поздней профазе мейоза I. Он образуется во время зиготены.
Необходимость у эукариот [ править ]
Внешние ссылки [ править ]
от 3D-Structured Illumination, фотография доктора Чунг-Джу Рэйчел Ван из Калифорнийского университета в Беркли, факультет молекулярной и клеточной биологии Беркли, Калифорния, США, занявшая второе место в конкурсе цифровых изображений Olympus Bioscapes 2009 года.
СОДЕРЖАНИЕ
Состав
У человека были охарактеризованы три специфических компонента синаптонемного комплекса: SC-белок-1 (SYCP1), SC-белок-2 (SYCP2) и SC-белок-3 ( SYCP3 ). SYCP1 гена на хромосоме 1p13; ген SYCP2 находится на хромосоме 20q13.33; а ген SYCP3 находится на хромосоме 12q.
Синаптонемный комплекс был описан Монтроузом Дж. Мозесом в 1956 г. в первичных сперматоцитах раков и Д. Фосеттом в сперматоцитах голубя, кошки и человека. Как видно под электронным микроскопом, синаптонемный комплекс образован двумя «латеральными элементами», в основном образованными SYCP3 и, во вторую очередь, SYCP2, «центральным элементом», который содержит по крайней мере два дополнительных белка и аминоконцевую область SYCP1, и «центральная область», расположенная между двумя боковыми элементами, которая содержит «поперечные филаменты», состоящие в основном из белка SYCP1.
SC можно увидеть в световой микроскоп с использованием окрашивания серебром или с помощью методов иммунофлуоресценции, которые маркируют белки SYCP3 или SYCP2.
Сборка и разборка
В процессе развития клетки синаптонемный комплекс исчезает на поздней профазе мейоза I. Он образуется во время зиготены.
Необходимость у эукариот
Внешние ссылки
от 3D-Structured Illumination, фотография д-ра Чунг-Джу Рэйчел Ван из Калифорнийского университета в Беркли, факультет молекулярной и клеточной биологии Беркли, Калифорния, США, занявшая второе место в конкурсе цифровых изображений Olympus Bioscapes 2009 года.
Функция синаптонемного комплекса.
Занятие № 13.
Мейоз.
Синаптонемный комплекс (СК).
Эффект Шульца-Редфильда.
Http://obi.img.ras.ru/humbio/MUTATION/0006e7bd.htm
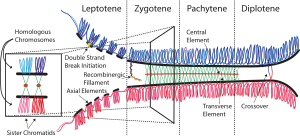
Для генетической рекомбинации необходимо тесное сближение рекомбинирующих хромосом. Синаптонемный комплекс, который формируется перед самой пахитеной и распадается сразу после нее, удерживает гомологичные хромосомы рядом, скрепляя их по всей длине. Синаптонемный комплекс представляет собой длинное белковое образование, напоминающее веревочную лестницу, к противоположным сторонам которого прилегают два гомолога. Сестринские хроматиды каждого гомолога остаются тесно сближенными, а их ДНК образует многочисленные петли по ту же сторону от белковой «лестницы». Таким образом, хотя гомологичные хромосомы в синаптонемальном комплексе сближены по всей длине, материнские и отцовские хроматиды, которые в последствии будут обмениваться участками, остаются по разные стороны от «лестницы», причем разделяющее их расстояние превышает 100 нм.
Как показывают цитологические исследования, конъюгации хромосом предшествует формирование белковой нити вдоль каждого из гомологов. По мере осуществления конъюгации эти нити, по-видимому, сближаются, превращаясь в боковые элементы синаптонемного комплекса и образуя две стороны белковой «лестницы». И первоначальные нити, и эти боковые элементы содержат белок, который очень хорошо окрашивается серебром, что позволяет видеть эти структуры как с помощью светового микроскопа, так и на электронных микрофотографиях.
Латеральные элементы синаптонемного комплекса.
Латеральные элементы формируются путем последовательного присоединения уже готовых блоков. Последнее приводит к тому, что в начале мейотической профазы отдельные участки гомологичных хромосом могут иметь уже сформированные латеральные элементы, в то время как в других участках хромосом они еще отсутствуют. Обычно латеральные элементы выглядят как одиночные линейные структуры, хотя в действительности каждый из гомологов состоит из двух сестринских хроматид. В некоторых случаях, например, в случае асинапсиса латерального элемента, наблюдается его локальное или полное разделение на два элемента, которые обычно более сближены, чем латеральные элементы синаптонемного комплекса.
Функция синаптонемного комплекса.
Сходные взгляды на функцию синаптонемного комплекса были высказаны Эгелем-Митани еще в 1982 году (Egel-Mittani et al.,1982); к сожалению, эти мысли не стали популярными.
Дата добавления: 2018-09-22 ; просмотров: 468 ; Мы поможем в написании вашей работы!
Белковые механизмы мейоза

Об авторе
Юрий Федорович Богданов, доктор биологических наук, профессор, главный научный сотрудник Института общей генетики им. Н. И. Вавилова РАН. Область научных интересов: клеточная биология, цитогенетика, проблемы организации эукариотической хромосомы, проблемы генетического контроля мейоза и биологии половых клеток.
Мейоз — важнейший процесс клеточного деления, происходящего накануне формирования половых клеток и открытый еще в конце XIX в., долгое время оставался предметом пристального внимания весьма узкого круга цитологов. Он попал в поле зрения молекулярных биологов лишь в 90-х годах XX в. Бурному развитию исследований в этой области способствовали работы по молекулярной генетике модельных объектов, а также появление новых иммуноцитохимических методов, которые дали в руки исследователей удобный способ изучения белков, участвующих в мейозе [1].
У всех эукариот во время мейоза формируется субмикроскопическая структура, получившая название синаптонемный комплекс (от греч. synaptos — соединенный, пета — нить). Исследование молекулярной организации этого комплекса и его роли в мейозе показало, что он нужен для рекомбинации хромосом и редукции их числа. Об этом и пойдет речь в данной статье.

Но сначала напомним основные сведения о мейозе, состоящем из двух делений: мейоза I и мейоза II. В результате редукционного деления (мейоза I) число хромосом в дочерних клетках уменьшается в два раза по сравнению с набором хромосом родительской клетки. Это происходит потому, что количество ДНК в хромосомах удваивается только один раз перед мейозом I (рис. 1). Двукратная редукция числа хромосом в ходе формирования половых клеток позволяет при оплодотворении восстановить исходное (диплоидное) число хромосом и сохранить его постоянство. Для этого необходимо строгое разделение пар гомологичных хромосом между половыми клетками. При ошибках возникает анеуплоидия — нехватка или избыток хромосом, и этот дисбаланс приводит к гибели зародыша или тяжелым аномалиям развития (у человека к так называемым хромосомным болезням).
Структура и функция синаптонемного комплекса

Синаптонемный комплекс представляет собой две белковые оси гомологичных хромосом, соединяющихся с помощью белковой «застежки-молнии» (рис. 2). Зубцы «застежки» — это палочковидные димеры из параллельно уложенных и одинаково ориентированных белковых молекул с длинной α-спиралью в середине молекулы. У дрожжей S. cerevisiae — это белок Zip1, у млекопитающих и человека — SCP1 (SYCP1). Эти белки своими С-концами закреплены на хромосомных осях (латеральных элементах комплекса), а N-концами направлены навстречу друг другу, внутрь центрального пространства (рис. 3). На N-концах молекул находятся заряженные «шпоры» — чередующиеся пики плотностей положительных и отрицательных зарядов аминокислот (рис. 4), комплементарное взаимодействие которых обеспечивает прочную электростатическую связь зубцов.
Так называемое центральное пространство комплекса (щель между белковыми осями, заполненная зубцами «застежки», шириной около 100 нм), как и весь комплекс (его сечение — порядка 150–200 нм) в обычном световом микроскопе не видны, поскольку весь комплекс замаскирован хроматином. Впервые синаптонемный комплекс увидели на ультратонких (толщиной 0,8 мкм) срезах семенников речного рака и мыши с помощью просвечивающего электронного микроскопа. Его обнаружили в 1956 г. независимо друг от друга два американских исследователя — М. Мозес и Д. В. Фоссет [2].

Теперь при исследовании комплекса используют так называемый метод микроспредирования. Клетки семенников (или пыльников растений) после гипотонического шока помещают на пластиковую подложку, нанесенную на предметное стекло. Содержимое лопнувшей клетки фиксируется слабым раствором формальдегида и контрастируется солями тяжелых металлов (лучше всего — AgNО3). Стекло просматривают в фазовоконтрастном микроскопе и по косвенным признакам выбирают клетки, которые должны содержать комплекс. Кружочек пленки с нужной клеткой подхватывают на металлическую сеточку и помещают ее в электронный микроскоп (рис. 5). По необходимости перед контрастированием клетки обрабатывают антителами к интересующим исследователя белкам. Эти антитела метят калиброванными гранулами коллоидного золота, которые хорошо видны в электронном микроскопе.

В ходе профазы мейоза I синаптонемный комплекс удерживает параллельно расположенные гомологичные хромосомы почти до момента их построения на экваторе клетки (метафаза I). Хромосомы соединяются с помощью синаптонемного комплекса на некоторое время (от 2 ч у дрожжей до 2–3 сут. у человека), в течение которого между гомологичными хромосомами совершается обмен гомологичными участками ДНК — кроссинговер [3]. В кроссинговере, происходящем с частотой не менее одного события (чаще — два, реже три или четыре) на пару гомологичных хромосом, участвуют десятки специфичных для мейоза белков-ферментов.

Молекулярный механизм кроссинговера и его генетические последствия — это две большие темы, выходящие за рамки задач данного рассказа. Нас этот процесс интересует потому, что в результате него гомологичные хромосомы прочно связываются перекрещенными молекулами ДНК (хиазмами) и необходимость попарного удержания хромосомы с помощью синаптонемного комплекса отпадает (после завершения кроссинговера комплекс исчезает). Гомологичные хромосомы, соединенные хиазмами, выстраиваются на экваторе веретена клеточного деления и расходятся с помощью нитей веретена клеточного деления в разные клетки. После завершения мейоза число хромосом в дочерних клетках уменьшается вдвое.
Итак, только накануне мейоза I структура хромосом радикально меняется. Очень специфическая внутриядерная и межхромосомная структура — синаптонемный комплекс — возникает один раз в жизненном цикле организма на короткое время для попарного соединения гомологичных хромосом и кроссинговера, а затем демонтируется. Эти и многие другие события в ходе мейоза на молекулярном и субклеточном (ультраструктурном) уровнях обеспечиваются работой многочисленных белков, выполняющих структурные, каталитические и кинетические (моторные) функции.
Белки синаптонемного комплекса
Еще в далекие 70-е годы мы получили косвенные доказательства того, что синаптонемный комплекс формируется путем самосборки его элементов, которая может происходить и в отсутствие хромосом. Эксперимент поставила сама природа, а нам удалось его наблюдать. Оказалось, что у свиной аскариды в цитоплазме клеток, готовящихся к мейозу I, появляются пакеты или «штабеля» абсолютно правильно уложенных морфологических элементов синаптонемного комплекса (хотя в цитоплазме нет хромосом: они — в ядре). Поскольку на стадии подготовки клеток к мейозу в клеточных ядрах еще нет синаптонемного комплекса, появилось предположение о несовершенстве контроля очередности событий мейоза у этого примитивного организма. Избыток новосинтезированных белков в цитоплазме приводит к их полимеризации и возникновению структуры, не отличающейся от синаптонемного комплекса [3]. Эта гипотеза получила подтверждение только в 2005 г. благодаря работе интернациональной группы исследователей, работающих в Германии и Швеции. Они показали, что если ген, кодирующий белок зубцов «застежки-молнии» млекопитающих (SCP1), внедрить в соматические клетки, растущие на искусственной питательной среде, и активировать его, то внутри культивируемых клеток возникает мощная сеть из белков SCP1, «застегнутых» между собой так же, как в центральном пространстве комплекса. Формирование слоя из сплошных белковых «застежек-молний» в культуре клеток означает, что предсказанная нами способность белков комплекса к самосборке доказана [4].
В 1989 и в 2001 гг. сотрудники нашей лаборатории О. Л. Коломиец и Ю. С. Федотова исследовали естественный «демонтаж» синаптонемных комплексов на завершающих этапах их существования. Этот многоэтапный процесс лучше всего удалось проследить на материнских клетках пыльцы в пыльниках ржи, где есть частичная синхронность мейоза [5]. Выяснилось, что латеральные элементы комплекса демонтируются путем постепенного «раскручивания» белковой суперспирали, имеющей три уровня упаковки (рис. 6).

Основа протяженных латеральных элементов — комплекс из четырех белков когезинов (от англ. cohesion — сцепление). Накануне мейоза в хромосомах появляется специфичный белок когезин Rec8, который заменяет соматический когезин Rad21. Затем к нему присоединяются три других белка-когезина, присутствующие и в соматических клетках, но вместо соматического когезина SMC1 появляется специфический для мейоза белок SMC1b (его N-конец на 50% отличается от N-конца соматического белка SMC1). Этот когезиновый комплекс располагается внутри хромосомы между двумя сестринскими хроматидами, удерживая их вместе. С комплексом когезинов связываются мейоз-специфичные белки, которые становятся мажорными белками хромосомных осей и превращают их (эти оси) в латеральные элементы синаптонемного комплекса [6]. У млекопитающих мажорные белки синаптонемного комплекса — SCP2 и SCP3, у дрожжей белки Hop1 и Red1, а мейоз-специфичный белок — Rec8.
Эволюционный парадокс белков
У млекопитающих и дрожжей белки синаптонемного комплекса имеют разные аминокислотные последовательности, но их вторичная и третичная структуры одинаковы. Так, белок «застежки-молнии» SCP1 у млекопитающих и негомологичный ему белок Zip1 у дрожжей построены по единому плану. Они состоят из трех аминокислотных доменов: центральный — α-спираль, способная к формированию спирали второго порядка (суперспирализации), и два концевых домена — глобулы. Мажорные белки SCP2 и SCP3, не имеющие никакой гомологии с белками Hop1 и Red1 дрожжей и, видимо, с еще недостаточно изученными белками комплекса у растений, также строят морфологически и функционально одинаковые структуры синаптонемного комплекса [7]. Это значит, что первичная структура (последовательность аминокислот) этих белков — эволюционно нейтральный признак.
Итак, негомологичные белки у эволюционно далеких организмов строят синаптонемный комплекс по единому плану. Объясняя этот феномен, воспользуюсь аналогией со строительством домов из разных материалов, но по единому плану Важно, чтобы в таких домах были стены, перекрытия, крыша и чтобы строительные материалы соответствовали условиям прочности. Равным образом, при формировании синаптонемного комплекса необходимы латеральные элементы («стены»), поперечные филаменты (зубцы «застежки-молнии») — «перекрытия» и центральное пространство (помещение для «кухни»). Там должны поместиться «кухонные роботы» — комплексы ферментов рекомбинации, собранные в так называемые «рекомбинационные узелки».
Ширина центрального пространства синаптонемного комплекса у дрожжей, кукурузы и человека составляет примерно 100 нм. Это обусловлено длиной односпиральных участков ДНК, покрытых белком рекомбинации Rad51. Этот белок относится к группе ферментов (подобных бактериальному белку рекомбинации RecA), которые сохраняют гомологию со времен появления рекомбинации молекул ДНК (примерно 3,5 млрд лет назад). Неизбежность гомологии белков рекомбинации у далеких организмов определяется их функцией: они взаимодействуют с двойной спиралью ДНК (одинаковой у бактерий и млекопитающих), разделяя ее на односпиральные нити, покрывают их белковым чехлом, переносят одну нить в гомологичную хромосому и там снова восстанавливают двойную спираль. Естественно, что большинство ферментов, участвующих в этих процессах, сохраняют гомологию более 3 млрд лет. В противоположность им синаптонемные комплексы, появившиеся у эукариот после возникновения мейоза (около 850 млн лет назад), построены из негомологичных белков. но схема их доменного строения одинакова. Откуда взялась эта схема?
Подсказкой служит упомянутый белок Rec8, с которого начинается формирование хромосомных осей в цикле мейоза и который есть у всех изученных организмов. Можно предположить, что строительным материалом для осей мейотических хромосом и латеральных элементов синаптонемного комплекса могут быть любые итермедиатные белки, которые способны образовывать волокнистую структуру (SCP2, Hop1 и др.), взаимодействовать с когезином Rec8 и «осаждаться» на нем, как бетон на металлической арматуре.
В последние годы, испытывая трудности в проведении экспериментальной работы из-за недостаточного финансирования, мы стали активно использовать методы биоинформатики. Нас интересовал белок «застежки-молнии» у дрозофилы. Учитывая сходство вторичной и третичной структур белков Zip1 дрожжей и SCP1 человека, мы предположили, что белок «застежки-молнии» у дрозофилы имеет такое же строение. Мы приступили к работе в 2001 г., когда геном дрозофилы уже был секвенирован и стало известно, что в нем имеется примерно 13 тыс. потенциальных генов. Как же найти ген для искомого нами белка?
Среди 125 известных к тому времени генов мейоза у дрозофилы мы предвидели лишь одного кандидата на эту роль. Дело в том, что мутация гена c(3)G лишала хромосомы способности соединяться попарно с помощью «застежки-молнии» и вступать в рекомбинацию. Мы предположили, что у мутантов дефектен белок, формирующий субмикроскопические зубцы «застежки». Вторичная структура и конформация искомого белка должна быть аналогична белкам Zip1 и SCP1.
Зная, что ген c(3)G находится у дрозофилы в хромосоме 3, мы искали в базе данных об этом районе (составляющем 700 тыс. пар нуклеотидов) такую открытую рамку считывания, которая могла бы кодировать похожий белок. Мы понимали, что при отсутствии гомологии в первичной структуре искомого белка и дрожжевого их размер, организация (из трех доменов) и способность центрального домена формировать α-спираль определенной длины (около 40 нм) должны быть аналогичными. Об этом говорило сходство электронно-микроскопической картины синаптонемного комплекса в мейозе у дрожжей и у дрозофилы.
Просмотрели открытые рамки считывания почти для 80 генов в районе поиска. С помощью компьютерных программ, позволяющих прогнозировать вторичную структуру виртуального белка, его физико-химические свойства и распределение электростатических зарядов в молекулах, Т. М. Гришаева нашла такую рамку считывания на границе зоны локализации гена c(3)G. (Это не очень точно предсказали японские генетики на микроскопической карте хромосом.) Им оказался ген CG1J604 по геномной карте компании «Селера».
Мы заключили, что этот виртуальный ген должен быть давно известным геном c(3)G и кодировать белок, аналогичный белку Zip1 дрожжей. В ответ на наше сообщение [8] мы получили электронное письмо из США от С. Хоули. Он экспериментально доказал, что ген c(3)G кодирует белок, формирующий «застежку-молнию» между хромосомами в мейозе у дрозофилы [9]. Результаты наших работ совпали, но экспериментальная работа группы Хоули заняла около семи лет, а наша компьютерная работа силами трех человек — лишь около трех месяцев. Статьи вышли из печати одновременно. В 2003 г. мы опубликовали метод наших компьютерных поисков и привели примеры аналогичных виртуальных белков у других организмов [10]. Эту работу сейчас охотно цитируют зарубежные коллеги, и наш метод успешно работает в их руках в сочетании с экспериментальной проверкой. Так, в 2005 г. группа английских биологов обнаружила ген и белок зубцов «застежки-молнии» у растения Arabidopsis thaliana [11].
В заключение приведу пример еще одной находки в области молекулярной биологии мейоза, но надо начать с митоза. Для того чтобы в анафазе митоза хроматиды разошлись, нужно разрушить «склеивающий» их когезин. Гидролиз когезинов во время митоза — это генетически программируемое событие. А вот в метафазе мейоза I, когда гомологичные хромосомы выстроены на экваторе клетки и белковое веретено готово растащить их к полюсам, гидролиз когезинов оказывается невозможным. Именно поэтому обе хроматиды каждой хромосомы, склеенные между собой в области кинетического центра хромосом (кинетохора), направляются к одному полюсу (см. рис. 1). В конце 90-х годов японские исследователи, изучая мейоз у дрожжей, установили, что в районе кинетохора когезины защищены белком, названным ими шугошином (корень этого термина взят из лексикона самураев и означает защиту). Очень быстро мировое сообщество исследователей мейоза пришло к выводу, что аналогичные белки-шугошины есть у дрозофилы, у кукурузы и у других объектов. При этом гены, «запрещающие» разъединение хроматид в мейозе I у дрозофилы, были известны лет за 10 до этого, но их белковый продукт не был расшифрован. А в 2005 г. группа американских исследователей из Калифорнийского университета в Беркли, среди которых и наша соотечественница и моя давняя коллега по исследованию мейоза И. Н. Голубовская, сообщила, что во время метафазы I мейоза в хромосомах кукурузы шугошин ZmSGO1 расположен по обе стороны от кинетохоров, причем появляется он в этом районе только в том случае, если там уже есть когезин Rec8, которого он и защищает от гидролиза (но только в мейозе I). Эти результаты получены с помощью флюоресцирующих антител к белкам и конфокального микроскопа [12]. Остается добавить, что японские исследователи тут же сообщили, что шугошин защищает Rec8 от гидролиза, если шугошин дефосфорилирован. Фосфорилирование и дефосфорилирование, так же как ацетилирование и деацетилирование, — важные модификации, меняющие свойства белковых молекул.
Прикладной аспект

Все рассказанное — красивая фундаментальная наука, а можно ли использовать эти знания в практических целях? Можно. Еще в середине 80-х годов британские исследователи и наша лаборатория на разных экспериментальных моделях доказали, что, используя микроспреды синаптонемных комплексов, можно выявить в два раза больше хромосомных перестроек (делеций, транслокаций, инверсий) по сравнению с традиционным методом анализа хромосом на стадии метафазы (рис. 7). Дело в том, что синаптонемный комплекс — скелетная структура мейотических хромосом в профазе. В это время хромосомы примерно в 10 раз длиннее, что значительно повышает разрешающую способность анализа. Однако исследовать запутанные в клубок профазные хромосомы практически невозможно, а жесткие скелетные структуры си-наптонемного комплекса не боятся распластывания, и, кроме того, электронный микроскоп способен различать миниаберрации, недоступные световому микроскопу.
Мы задались вопросом: можно ли установить причину стерильности потомства облученных мышей, изучая не хромосомы, а синаптонемный комплекс? Оказалось, что у стерильных мышей, унаследовавших от родителей хромосомные транслокации, эти перестройки выявляются с помощью комплекса в 100% исследуемых клеток, а при обычных методах «метафазного» анализа — лишь в 50% клеток [13]. Группа испанских исследователей обследовала более 1 тыс. мужчин, страдающих бесплодием. У трети из них причину бесплодия ранее не удавалось установить, а изучение синаптонемного комплекса из клеток семенников этих пациентов позволило половине из них поставить диагноз: причина бесплодия в отсутствии синаптонемного комплекса, из-за чего сперматоциты (клетки-предшественники сперматозоидов) не развиваются, т. е. наблюдался «арест» процесса мейоза и всего сперматогенеза [14]. Аналогичные результаты получены О. Л. Коломиец совместно с врачами из Харькова. Исследование синаптонемного комплекса в сочетании с другими методами анализа повышает процент выявления причин бесплодия у обследованных пациентов-мужчин с 17 до 30% [15]. Некоторые английские клиники уже в 90-х годах XX в. активно использовали подобные методы. Такая диагностика, конечно, требует высокой теоретической и практической квалификации врачей и использования электронных микроскопов. Российские лаборатории еще не достигли такого уровня, за исключением Института общей генетики им. Н. И. Вавилова РАН (Москва) и Института цитологии и генетики СО РАН (Новосибирск).
Можно думать, что интенсивные исследования механизмов мейоза неизбежно приведут к применению полученных знаний в тех областях биологии и медицины, которые связаны с фертильностью живых организмов, включая человека. Однако закон применения научных достижений на практике неизменен: «внедрять» что-либо силой — бесполезно. Практики сами должны следить за достижениями науки и использовать их. Именно такой подход применяют передовые фармакологические и биотехнологические фирмы.
От открытия мейоза (1885) до открытия синаптонемного комплекса (1956) прошло примерно 70 лет, а с 1956 г. до открытия белков синаптонемного комплекса (1986) — еще 30. За последующие 20 лет мы узнали структуру этих белков, кодирующие их гены, взаимодействие белков при построении и работе синаптонемных комплексов, в частности, их взаимодействие с белками-ферментами рекомбинации ДНК и т. д., т. е. больше, чем за предшествующий 30-летний период описательных цитологических исследований. Возможно, для расшифровки основных молекулярных механизмов мейоза потребуется не более двух десятков лет. История науки, как и всей цивилизации, характеризуется «сжатием времени», нарастающим уплотнением событий и открытий.



