Что такое вирусный вектор
Идеальный вектор для генотерапии должен быть безопасным, легко изготавливаемым, легко вносимым в подходящую целевую ткань, при этом он должен экспрессировать нужный ген пожизненно. В настоящее время нет ни одного известного вирусного или невирусного вектора, удовлетворяющего всем этим критериям.
На самом деле, вероятно, нет единственного вектора, удовлетворительного во всех отношениях для всех типов генотерапии, и потребуется набор векторов. Здесь мы кратко рассмотрим три широко используемых класса вирусных векторов, производных от ретровирусов, аденовирусов и адено-ассоциированных вирусов. Основное преимущество вирусных векторов — то, что они способны проникнуть фактически в каждую клетку в целевой популяции.
Один из наиболее широко используемых классов векторов — производные от ретровирусов, простых РНК-вирусов всего с тремя структурными генами, которые могут быть удалены и заменены нужным геном. Текущее поколение ретровирусных векторов создано так, чтобы лишить их способности к репликации.
Другие их достоинства: нетоксичны в клетке; в геном хозяина внедряется (с передаваемым геном) только небольшое количество копий вирусной ДНК; встроенная ДНК стабильна; ретровирусные векторы могут встраивать вплоть до 8 килобаз дополнительной ДНК, что достаточно для многих передаваемых генов.
Основное ограничение большинства ретровирусных векторов в том, что для интеграции вируса в ДНК хозяина целевая клетка должна делиться, а это ограничивает использование таких векторов для неделящихся клеток, например нейронов. Тем не менее ретровирусы одного класса — лентивирусы, включающие ВИЧ, способны встраивать свою ДНК во множество медленно делящихся и даже неделящихся клеток, включая нейроны. Эти векторы могут оказаться пригодными для лечения неврологических заболеваний.
Адено-ассоциированные вирусы имеют большое преимущество — они не имеют никаких неблагоприятных эффектов у больных и широко распространены в популяциях человека. Кроме того, они заражают как делящиеся, так и неделящиеся клетки и могут существовать в виде эписом или стабильно интегрироваться в хромосому хозяина.
Их основной недостаток состоит в том, что имеющиеся на настоящий момент адено-ассоциированные вирусные векторы могут встраивать не более 5 килобаз дополнительной ДНК.
Аденовирусные векторы имеют свои преимущества — их можно получать в высоком титре; они заражают множество типов клеток, как делящихся, так и неделящихся; они могут встраивать в себя от 30 до 35 килобаз ДНК. Тем не менее, помимо других ограничений, их применение недавно было связано по крайней мере с одной смертью при испытании генотерапии вследствие развития сильной иммунной реакции. Следовательно, возможность их использования в целях генотерапии в настоящее время тщательно проверяется.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Характеризация вирусных векторов: анализ гомогенности rAAV
Решение для измерения гомогенности вектора на основе rAAV: чистота (и более) – анализ вирусных частиц в растворе.
Векторы на основе рекомбинантных аденоассоциированных вирусов (rAAV) открывают перспективы для развития новых методов генной терапии, способных спасать жизни.
Классические методы, такие как электронная микроскопия и саузерн-блоттинг, позволяют охарактеризовать rAAV по гетерогенности и агрегации. Однако они не обеспечивают достаточного разрешения для количественной оценки гомогенности и нагрузки вирусными частицами. Когда речь идет о создании векторных препаратов на основе rAAV для потенциального клинического применения, достижение необходимого разрешения измерений всегда вызывает трудности.
 При исследованиях в области генной терапии, быстрое получение отрицательного результата особенно важно на ранних этапах разработки продукта, когда ставки очень высоки. Но до настоящего времени, когда речь заходила о создании векторов на основе rAAV для клинического применения, этого невозможно было достичь с помощью обычных технологий.
При исследованиях в области генной терапии, быстрое получение отрицательного результата особенно важно на ранних этапах разработки продукта, когда ставки очень высоки. Но до настоящего времени, когда речь заходила о создании векторов на основе rAAV для клинического применения, этого невозможно было достичь с помощью обычных технологий.
Каково главное препятствие? Отсутствие метода количественного определения с высоким разрешением для контроля терапевтического качества с учетом:
Новое решение: анализ в растворе с применением аналитического ультрацентрифугирования (AUC).
Исследователи из Genethon и их коллеги установили, что благодаря возможности проведения безматричного анализа векторных препаратов на основе rAAV, независимо от серотипа и трансгенов, аналитическое ультрацентрифугирование позволяет:
Просмотрите этот вебинар и узнайте, как Кристин Ле Бек из компании Genethon и ее исследовательская группа использовали AUC для изучения характеристик векторов scAAV и ssAAV – в частности определяли гомогенность и нагрузку вирусными частицами.
Вирус, дарующий зрение
Автор
Редакторы
Статья на конкурс «Био/Мол/Текст»: Слепота — не всегда приговор, ведь вирусы могут помочь восстановить утраченное зрение. Но какие это вирусы, как их применять? О том, что такое вирусные векторы и как именно они участвуют в восстановлении зрения, вы узнаете в этой статье.

Конкурс «Био/Мол/Текст»-2020/2021
Эта работа опубликована в номинации «Свободная тема» конкурса «Био/Мол/Текст»-2020/2021.
Генеральный партнер конкурса — ежегодная биотехнологическая конференция BiotechClub, организованная международной инновационной биотехнологической компанией BIOCAD.

Спонсор конкурса — компания SkyGen: передовой дистрибьютор продукции для life science на российском рынке.
Спонсор конкурса — компания «Диаэм»: крупнейший поставщик оборудования, реагентов и расходных материалов для биологических исследований и производств.
Люди всегда боялись различных вирусов, а с появлением нового SARS-CoV-2 этот страх только усилился. Однако не все вирусы оказывают отрицательное воздействие на организм человека, а учитывая свойства и особенности некоторых из них, можно даже вылечить заболевания. В этой статье речь пойдет про аденоассоциированные вирусы и их применение в офтальмологии.
Знакомство с аденоассоциированными вирусами
Геном вирусов
Аденоассоциированные вирусы (AAV) — это небольшие, безоболочечные вирусы, принадлежащие к роду Dependovirus семейства Parvoviridae. Первый аденоассоциированный вирус был открыт в 1965 году как загрязнитель препаратов аденовируса (Ad) [1]. Вирусные частицы выявили случайно во время микроскопии лабораторных препаратов аденовируса, исходно полученных от обезьян (макак резус) с целью производства вакцины.
Первоначально считалось, что это клеточный мусор, однако впоследствии было доказано, что AAV является отдельным вирусом, который по многим структурным, иммунологическим и генетическим показателям отличается от аденовируса.
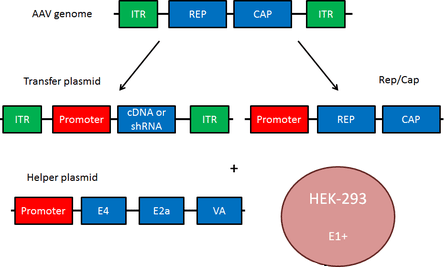
Рисунок 1. Плазмидная система на основе AAV
Геном аденоассоциированного вируса (рис. 1, вверху) представлен линейной одноцепочечной ДНК с двумя инвертированными концевыми повторами (ITR), которые в основном необходимы для репликации и служат сигналом упаковки. AA-вирусы имеют три гена. Первый ген rep с помощью двух промоторов и альтернативного сплайсинга кодирует четыре регуляторных белка, которые получили названия Rep78, Rep68, Rep52 и Rep40 [1]. Цифры соответствуют их молекулярной массе в кДа. Эти белки участвуют в репликации генома AAV. Также было доказано, что эти четыре Rep-белка обладают хеликазной активностью и способностью связывать АТФ.
Ген cap посредством альтернативного сплайсинга и инициации трансляции дает три капсидных белка: VP1 (белок вириона 1), VP2 и VP3 с молекулярной массой 87, 72 и 62 кДа соответственно. Эти капсидные белки образуют белковую оболочку вируса из 60 субъединиц в соотношении 1:1:10 (VP1:VP2:VP3). VР3 также активирует сборку новых вирионов [1].
На сегодняшний день выявлено 11 серотипов аденоассоциированных вирусов, из которых наиболее хорошо изучен серотип AAV2. Он обладает высоким уровнем тропизма по отношению к клеткам скелетных мышц, нейронам, гладкомышечным клеткам сосудов и гепатоцитам. До 1990-х годов геном AA-вирусов изучали на примере AAV2. Структура генома данного серотипа представлена на рисунке 2.
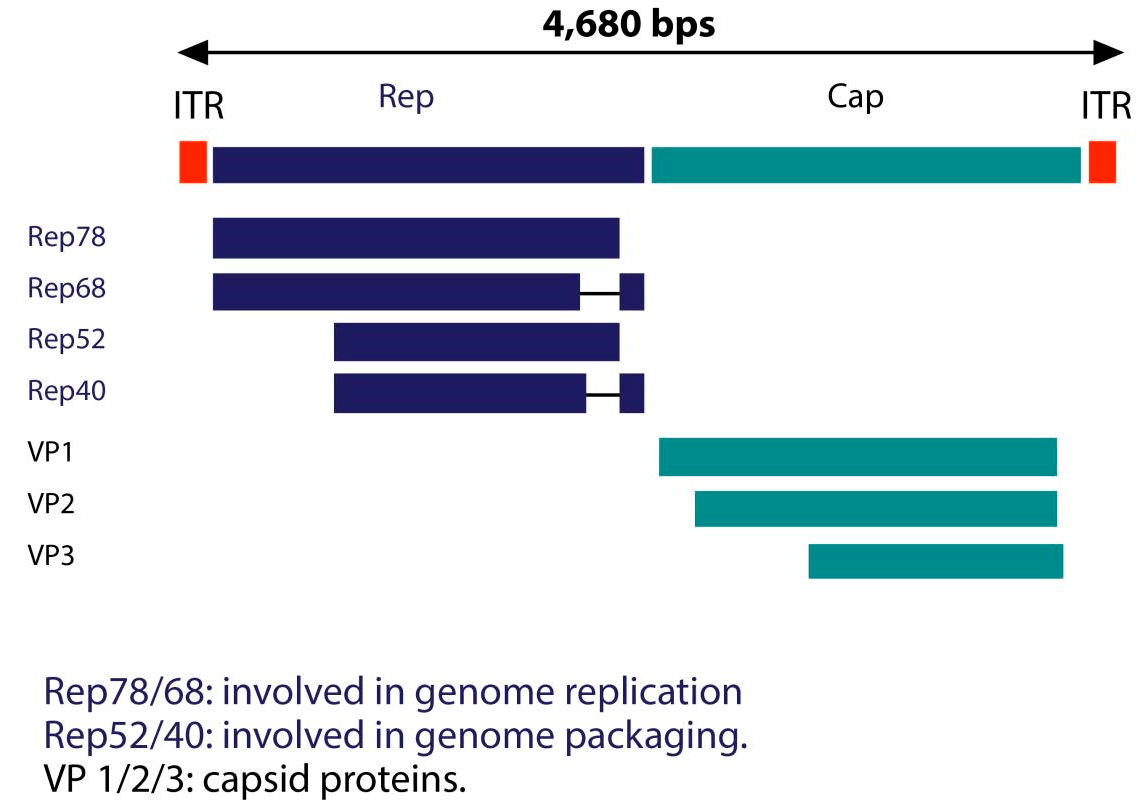
Рисунок 2. Структура генома AAV2
Жизненный цикл вируса
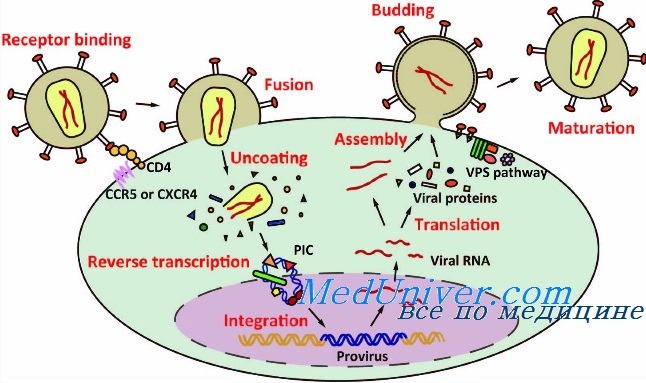
AAV связываются с клеткой хозяина за счет использования гепарансульфатных протеогликанов в качестве сайтов для стыковки. Благодаря тому, что аденоассоциированные вирусы используют достаточно распространенный фактор передачи инфекции, они имеют широкий круг хозяев и могут инфицировать клетки самых разных типов. Вирус проникает внутрь клетки через плазматическую мембрану посредством эндоцитоза, после чего попадает в цитозоль и захватывается эндосомой. Механизм транспортировки AAV в ядро до сих пор полностью не изучен. Однако недавно благодаря визуализации молекул AVV флуоресцентными метками удалось отследить траекторию движения вирусных вирионов в клетке. Выяснили, что в активном внутриклеточном транспорте вирусов участвуют микротрубочки и моторные белки.
После проникновения через ядерные поры в ядро клетки-хозяина, данный вирус может следовать по одному из двух путей своего цикла: литическому или лизогенному. Первый развивается в клетках, инфицированных вспомогательным вирусом, например, таким как аденовирус или вирус простого герпеса (ВПГ), тогда как второй — при их отсутствии. Если инфицированная клетка содержит в себе вспомогательные вирусы, активизируется программа экспрессии генов AAV, что дает возможность вирусам реплицироваться с использованием полимеразы клетки-хозяина.
В процессе жизненного цикла по лизогенному пути AA-вирус интегрируется в геном хозяина в конкретном месте — сайте AAVS1 на 19 хромосоме человека. Это сайт-специфическая интеграция, которая происходит с участием ITR AAV и белков Rep (Rep78, Rep68). Сборка капсида происходит в ядре инфицированных клеток. Процессы образования капсида генома пока малоизвестны. Однако установлено, что их образование происходит в результате взаимодействия белка, активирующего сборку (VP3), и капсидных белков. После сборки капсидов в нуклеоплазме происходит упаковка сформированной ДНК.
Когда вирус-помощник вызывает лизис клетки-хозяина, высвобождаются новые собранные вирионы AAV.
Жизненый цикл и трансдукция AA-вирусов на примере рекомбинантного AAV (rAAV), который используется в качестве вектора в генной терапии, представлен на рисунке 3.
![]()
Рисунок 3. Трансдукция AAV на примере рекомбинантного AAV (rAVV)
Процесс осуществления зрения
Зрительный аппарат состоит из глазного яблока и вспомогательного аппарата, который включает в себя веки, ресницы, слезные железы и мышцы глазного яблока.
Важные зрительные процессы, а именно преобразование фотонов в энергию (которая затем используется в качестве импульса зрительного сигнала) и построение изображения, происходят в сетчатке глаза.
Сетчатка состоит из фоторецепторов и нервных клеток. Нервные клетки представлены биполярными нейронами и ганглиозными клетками, которые образуют зрительный нерв (рис. 4).
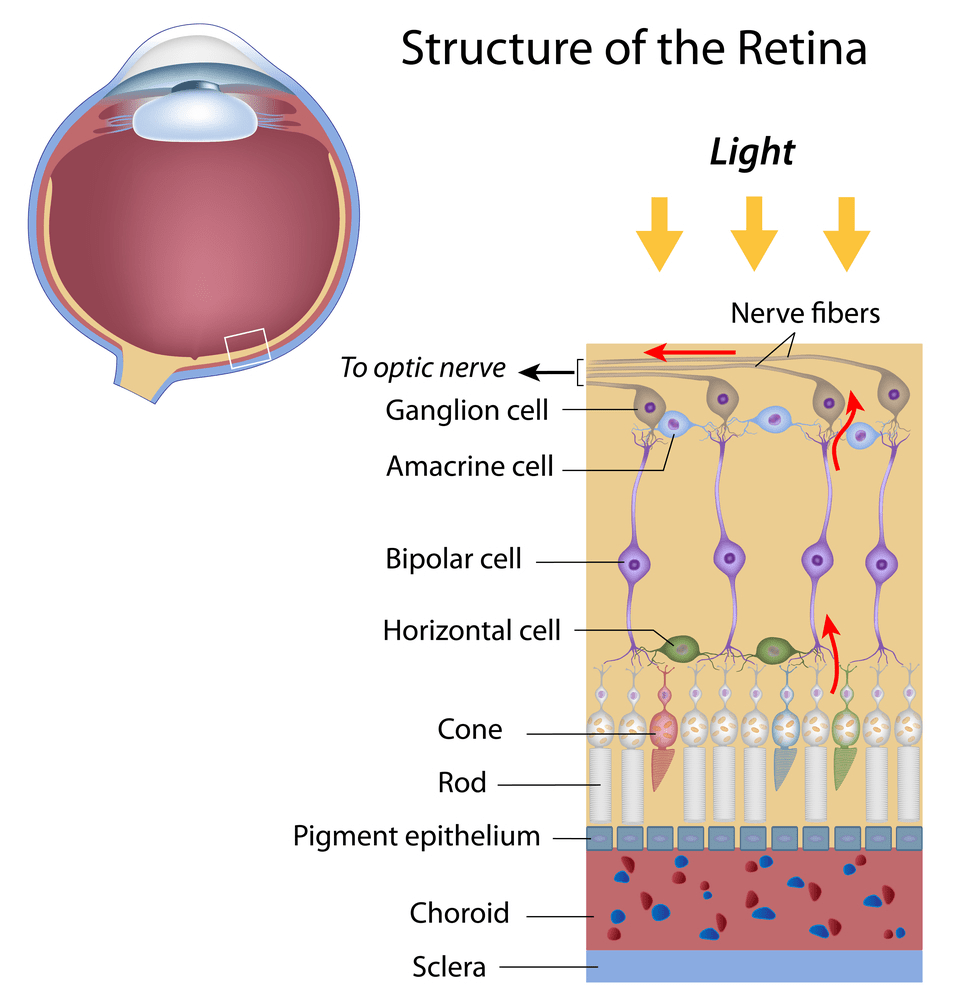
Рисунок 4. Строение сетчатки
Фоторецепторы представлены палочками и колбочками. Палочки обладают высокой светочувствительностью и позволяют видеть при плохом освещении. Эти клетки содержат вещество родопсин, благодаря которому они быстро возбуждаются слабым светом, но при этом воспринимать цвет не могут. Подробнее про молекулярную основу зрения и действие родопсина написано в статье «Зрительный родопсин — рецептор, реагирующий на свет» [2].
Механизм работы родопсина
При поглощении света происходит процесс фотолиза родопсина, в ходе которого 11-цис-ретиналь полностью изомеризуется в транс-форму, что меняет ионный транспорт в фоторецепторе. Цикл преобразования родопсина представлен на рисунке 5.
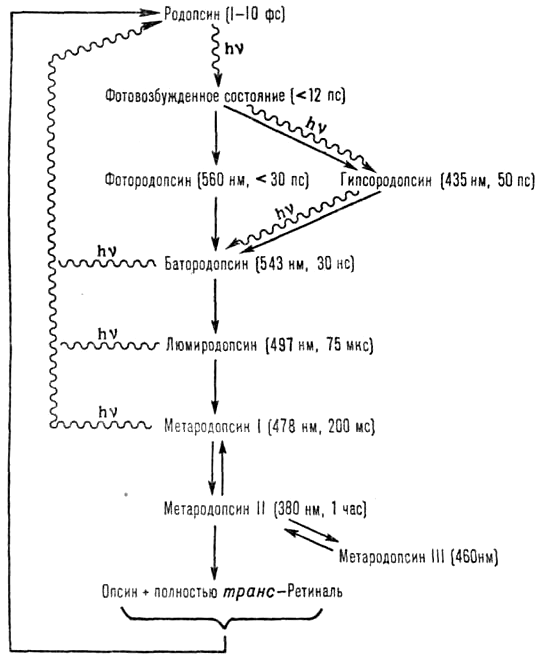
Рисунок 5. Цикл преобразования родопсина
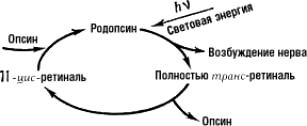
Рисунок 6. Изомеризация 11-цис-ретиналя в транс-форму
Под действием света меняется конфигурация белковой части родопсина и происходит активация G-белка трансдуцина, который активирует фермент цГМФ-фосфодиэстеразу. В результате активации этого фермента в клетке снижается концентрация цГМФ и закрываются цГМФ-зависимые натриевые каналы. Так как ионы натрия постоянно вытесняются из клетки АТФазой, их концентрация внутри клетки падает, что, в свою очередь, приводит к ее гиперполяризации. В результате фоторецептор выделяет меньше тормозного медиатора глутамата, а в биполярной нервной клетке возникают нервные импульсы (рис. 6).
Колбочки обеспечивают нам цветное зрение. Причем есть три вида колбочек, которые отвечают за красный, синий и зеленый цвета. При их взаимодействии глаз воспринимает всевозможные цвета. Зрительный пигмент колбочек — иодопсин — также содержит остаток ретиналя. Однако его белковый компонент отличается от опсина палочек. Процесс превращения иодопсина схож с превращениями родопсина. Электрические сигналы, которые производят колбочки и палочки, прежде чем они попадут в мозг по зрительному нерву, обрабатываются сначала другими клетками сетчатки — биполярными и ганглиозными клетками. Существуют еще два дополнительных слоя промежуточных нейронов. Горизонтальные клетки передают сигналы туда и обратно между клетками фоторецепторов, биполярным клеткам и друг другу. Амакриновые клетки (клетки сетчатки) взаимодействуют с биполярными клетками, ганглиозными клетками, а также друг с другом. Оба вида таких промежуточных нейронов играют главную роль в обработке визуальной информации на уровне сетчатки перед тем, как она передается в мозг для конечной обработки.
Большие шаги науки
В этой части статьи я хочу поведать про достижения генной инженерии в области офтальмологии.
Согласно статистике, которую приводит Всемирная организация здравоохранения, около 1,3 млрд человек в мире живут с той или иной формой нарушения зрения, тогда как 36 млн человек поражены слепотой [3]. Одной из причин слепоты является дегенерация сетчатки в силу возраста или в результате каких-то врожденных мутаций. Эти нарушения часто приводят к гибели светочувствительных клеток (палочек и колбочек), при этом другие клетки (ганглионарные и биполярные), которые принимают и передают сигналы от светочувствительных клеток, повреждаются редко. Такая вот интересная особенность. Ученые с помощью генной инженерии пытаются придать оставшимся клеткам дополнительно функции светочувствительных клеток, чтобы компенсировать потерю последних. Вирусные векторы активно используют также и для лечения других заболеваний, таких как гемофилия, анемия, спинальная мышечная атрофия и рак: об этом можно подробнее почитать на «Биомолекуле» в статьях «Генная терапия против рака» [4] и «Сводка с генотерапевтических фронтов. Новая стратегия нейтрализации гемофилии» [5].
Слепые тоже смогут видеть
Подробнее об особенностях препаратов для генной терапии на основе вирусных векторов можно прочитать в статье «Генная терапия: познакомьтесь с лекарствами будущего» [6].
Последние исследования и клинические испытания подтверждают терапевтическую эффективность вирусных векторов в лечении дегенеративных процессов в сетчатке у людей. Например, недавно Европейская медицинская ассоциация одобрила препарат Luxturna на основе AAV2-вектора [7], который предназначен для лечения редкой формы аутосомно-рецессивной слепоты, вызванной мутациями в гене RPE65.
Как разрабатываются новые генные терапии в офтальмологии
Изменения в RPE65 — далеко не единственно возможная причина проблем с сетчаткой. К прогрессирующей потере зрения может приводить и нарушение синтеза опсина, вызванное мутацией в гене PDE. Чтобы найти решение этой проблемы ученые недавно провели эксперимент на мышах, в ходе которого инактивированный AA-вирус использовали в качестве вектора для доставки гена опсина в ДНК светочувствительных клеток, содержащихся в колбочках.
Подробнее о том, как можно встроить в вирусный вектор нужный ген, описано в статье спецпроекта «12 методов в картинках»: «Генная инженерия. Часть II: инструменты и техники» [8].
Тут важно отметить, что все новые методы терапии отрабатывают на разных лабораторных животных, а затем успешный препарат может выйти в клинические испытания на людях. Так что работа ученых весьма многообещающая!
Что же делали в этом исследовании? Эксперимент проводили с мышами, у которых была мутация в гене PDE6β [9]. Из-за мутаций гена нарушается синтез опсина средней длины волны (MW-опсин), в результате чего фоторецепторные клетки теряют свою зрительную способность. Ген MW-опсина с флуоресцентной (YFP) C-концевой меткой для отслеживания экспрессии был упакован в AAV2 и введен интравитреально (в стекловидное тело глаза) на 45–60 сутки после рождения мышей. Модифицированный вирус избирательно связывается с ганглионарными клетками сетчатки, за счет своего тропизма к нейронам. Закрепившись на поверхности, он вносит внутрь клетки ДНК, кодирующую ген опсина. В результате ганглионарные клетки получают способность синтезировать чувствительный белок, начинают регистрировать свет и снова передавать зрительные сигналы дальше по цепочке.
Для оценки результатов подопытных мышей через 8 часов после введения AAV поместили в коробку, где находились два объекта. Мышь оставляли у стены, достаточно далеко от объектов, которые также находились на некотором удалении друг от друга. Движения мышей отслеживали в течение 10 минут при выходе на арену. На рисунке 7 вы можете видеть, как изменилось поведение мышей в незнакомой клетке после введения гена.

Рисунок 7. Перемещение мыши в незнакомой клетке. а — Мышь слепая от рождения. б — Мышь с восстановленным зрением. в — Клинически здоровая мышь.
Животные, которым ввели ген опсина, двигались в 1,6 раз дальше и в 1,59 раза быстрее, чем слепые мыши.
Для дальнейшего анализа проводили оценку исследовательского поведения животных в новой клетке. Инъецированные мыши и мыши, синтезирующие родопсин, действовали аналогично. Однако мыши с MW-опсином проходили более короткие пути и достигали первого и второго объекта быстрее по сравнению с мышами, не получавшими лечения. Эти результаты предполагают, что MW-опсин восстанавливает фоторецепторные клетки и обеспечивает ранее слепым животным различение объектов при естественном освещении.
На мышах уже тестировали модели лечения и других глазных заболеваний, таких как врожденный амавроз Лебера и дегенерация желтого пятна [10]. Интересно и другое исследование, где изучали мутации в гене ABCA4 [10], которые вызывают болезнь Штаргардта — форму аутосомно-рецессивной ювенильной дегенерации желтого пятна. Субретинальная инъекция вектора AAV2/5, несущего ABCA4, привела у мышей к снижению содержания липофусцина и улучшению морфологии и функции сетчатки на срок до пяти месяцев.
Благодаря позитивным результатам лечения на мышиных моделях, уже начаты клинические испытания на людях. Пациентам с тем или иным недугом субретинальным путем вводят вирусные векторы для проверки переносимости и безопасности. Побочных эффектов со стороны иммунной системы пока не наблюдалось [11], и у трех из семи пациентов отмечалось повышение остроты зрения в течение 12-месячного испытательного периода.
Заключение
Благодаря этим опытам мы видим, что применение AAV-векторов хорошо зарекомендовало себя в решении целого ряда нарушений зрительных функций. Они позволяют точно доставлять гены в клетки-мишени, не вызывая побочных действий у пациента в большинстве случаев. Вирусные векторы — это одно из перспективных направлений генной инженерии. Конечно, необходимо провести еще ряд клинических исследований, чтобы окончательно удостовериться в безопасности и эффективности векторной терапии. Но, возможно, уже в скором времени для лечения слепоты разной этиологии мы будем использовать вирусы.
Вирусные векторы для доставки генетического материала в клетку и их использование в нейробиологии (обзор)
В настоящее время вирусные векторы — распространенный инструмент для доставки генетического материала в клетку. Именно благодаря особенностям жизненного цикла вирусов первые векторы (носители трансгенов) стали разрабатывать на их основе [1]. Вирусы переносят чужеродные гены, которые способны экспрессироваться в зараженных клетках [2]. Сейчас вирусы эволюционировали в специализированные молекулярные механизмы, способные эффективно транспортировать их геномы внутрь клеток, которые они заражают [3].
Разнообразие вирусных векторов велико, и каждый имеет свои преимущества и недостатки. Сейчас значительные усилия направляются на разработку вирусных векторов с улучшенными характеристиками безопасности и большей эффективностью доставки нуклеиновой кислоты в клетки, а также на обеспечение длительной и тканеспецифичной экспрессии введенного генетического материала. Потенциально вирусные векторы могут применяться как один из способов доставки генов в генной терапии новообразований и наследственных заболеваний.
Рекомбинантные аденоассоциированные вирусы (рААВ) являются одними из наиболее перспективных векторов доставки для генной терапии благодаря своим непатогенным свойствам, отсутствию иммуногенности со стороны хозяина и тропности к большинству клеток и тканей [4]. Аденоассоциированные вирусные векторы по своим характеристикам приближаются к идеальному вектору [5].
Вирусные векторы нашли свое применение и в нейробиологии в качестве систем эффективной доставки генов в нейроны и другие нейрональные клетки in vitro и in vivo. Вирусные векторы позволяют вести наблюдение за нейробиологическими функциями, изменять экспрессию целевых генов, метить клетки для определения их судьбы и менять физиологическое состояние специфических клеточных популяций. Использование вирусных векторов для доставки генов в нервную систему имеет большие перспективы как для фундаментальных исследований, так и для терапевтического применения.
Типы вирусных векторов, используемых в нейробиологии
Существуют вирусные и невирусные системы доставки генетического материала. Вирусные системы включают векторы, разработанные на основе ретровирусов, аденовирусов, аденоассоциированных вирусов, лентивирусов и вирусов простого герпеса. Невирусные системы включают «голую» ДНК и липиды или полиэтилгликоль [4]. Независимо от того, какой геном имеют вирусные векторы в своем составе — ДНК или РНК, у них похожий жизненный цикл, который начинается с взаимодействия со специфическим рецептором на поверхности клетки. После адсорбции вириона на клеточной поверхности в процессе проникновения геном вирусов подвергается «раздеванию». При реализации внутриклеточной стадии жизненного цикла вирус осуществляет три молекулярных процесса: репликацию геномной нуклеиновой кислоты, транскрипцию и трансляцию. На каждой стадии вирус вмешивается в клеточные синтетические механизмы и подчиняет их своим задачам, создавая приоритеты для вирусных нуклеиновых кислот [6].
Ни ДНК, ни РНК не могут быть использованы в «голом» виде для доставки генов в клетки-мишени. Сахарофосфатный остов молекул нуклеиновых кислот располагается по их периферии полярными группами наружу, придавая им анионные свойства [7]. При физиологических значениях рН нуклеиновая кислота несет отрицательный заряд, отталкивающий ее от отрицательно заряженной наружной поверхности клеточной мембраны. Еще одно ограничение при проникновении в клетку нуклеиновой кислоте создает ее гидрофильность [8]. Все ее гидрофобные основания «повернуты» вовнутрь молекулы, поэтому она не может проникнуть через гидрофобный барьер клетки-мишени [9]. В сыворотке крови нуклеиновая кислота быстро деградирует под воздействием нуклеаз. Период полужизни немодифицированной интерферирующей РНК в сыворотке крови укладывается в 5–60 мин, для ДНК он составляет не более 10 мин [10]. Кроме того, нуклеиновые кислоты не способны специфически узнавать клетки-мишени. Поэтому для доставки генов в эукариотические клетки с начала 1980-х гг. разрабатываются векторные генетические конструкции [11].
Ретровирусные векторные системы
Ретровирусы относятся к группе вирусов, РНК-геном которых в инфицированных клетках конвертируется в ДНК. Геном ретровирусов образован плюс-цепью РНК и включает три структурных гена (gag, pol и env), фланкированных длинными терминальными повторами (LTR, long terminal repeat) [12]. В LTR содержатся регуляторные элементы, которые играют роль во встраивании ретровируса и необходимы для интеграции ДНК-копии генома вируса с геномом хозяина, а также определяют начало и конец вирусного генома. LTR также контролируют экспрессию вирусных генов [13]. Оболочка ретровирусов состоит из цитоплазматической мембраны зараженной клетки и вирусных белков [14]. Особенностью жизненного цикла ретровирусов является обратная транскрипция в зараженной клетке на матрице вирусной РНК с образованием двухцепочечной ДНК (провирус), которая затем встраивается в клеточный геном. Вирус встраивается в клеточный геном случайным образом, поэтому возрастает риск инсерционного мутагенеза. Так как эти вирусы заражают только делящиеся клетки (для проникновения в ядро необходимо разрушение ядерной оболочки клетки, происходящее в ходе митоза), ретровирусные векторы используют в основном для трансфекции клеток ex vivo или для лечения злокачественных опухолей [15]. Сами ретровирусные векторы получают на основе провируса, из которого удаляются гены gag, pol и env для предотвращения репродукции вируса и освобождения места для желаемого генетического материала. В вектор на основе ретровируса может быть включено до 8 тыс. пар оснований ДНК-вставки. Для репродукции вируса удаленные вирусные гены (gag, pol, env), которые кодируют вирусные белки и обеспечивают репродукцию вируса, встраивают в геном упаковывающей клеточной линии на разные хромосомы так, чтобы снизить вероятность обратной рекомбинации вирусных генов в исходный вирусный геном и образования вирусов, которые могут репродуцироваться [16].
Лентивирусные векторные системы
Лентивирусы относятся к семейству ретровирусов и в отличие от других ретровирусов инфицируют не только делящиеся, но и неделящиеся клетки. Наиболее исследованным лентивирусом является вирус иммунодефицита человека (ВИЧ). В связи со способностью лентивирусов включать большое количество генетического материала (до 8 тыс. пар оснований) и инфицировать делящиеся и неделящиеся клетки, эти вирусы являются перспективным вектором для доставки генов в условиях in vivo [17]. Геном ВИЧ включает три гена структурных белков (gag, pol и env) и 6 генов регуляторных белков (tat, rev, vpr, vpu, nef и vif) [18, 19]. Некоторые гены могут быть исключены из генома вируса и при этом не вызвать снижения вирусной способности к размножению и инфицированию клеток. Лентивирусный жизненный цикл похож на жизненные циклы других ретровирусов, за исключением способности заражать и неделящиеся клетки. Эта способность обеспечивается взаимодействием вирусного преинтеграционного комплекса с ядерной оболочкой и транспортом через нее [20]. Сборка лентивирусного вектора происходит в упаковывающих клетках — это перевиваемые клетки, осуществляющие синтез вирусоспецифических белков [21]. Упаковывающие клетки включают в себя пакующую, векторную и оболочечную кассеты, которые вместе позволяют собрать функциональную вирусную частицу [22, 23]. При этом их одновременная экспрессия не вызывает образования ретровирусных частиц, способных инициировать инфекционный процесс у человека [24].
Лентивирусы имеют сравнительно небольшой объем вставки целевого гена (до 8 тыс. пар оснований), могут обеспечить длительную экспрессию трансгена и индуцируют минимальный иммунный ответ организма-хозяина [25]. Некоторые авторы считают эти векторы малоподходящими для доставки in vivo, так как они увеличивают риск инсерционного мутагенеза. Однако при использовании лентивирусов для трансдукции дифференцированных клеток риск инсерционного мутагенеза меньше, чем при использовании других ретровирусов [26].
Векторные системы на основе аденовирусов
Аденовирусы — семейство ДНК-вирусов, несущих в своем составе одну двуцепочечную молекулу ДНК и лишенных липидной оболочки. Аденовирусы разделены на основе связывания со специфическими сыворотками на 51 серотип [27], а по способности агглютинировать эритроциты у людей, кроликов и мышей и по онкогенности для грызунов — еще на 6 субгрупп (от A до F) [28]. Аденовирусы разных субгрупп поражают различные органы и ткани человека [29].
Создание рекомбинантных аденовирусных векторов во многом схоже с созданием лентивирусных векторов. Аденовирусы, дефектные по репликации, получали за счет замены гена Е1, необходимого для репликации, на ген интереса, промотор и энхансер [30]. При этом такие рекомбинантные векторы эффективно размножаются в пакующих клетках, экспрессирующих продукт гена Е1. Неспособные к репликации вне пакующих клеток аденовирусные векторы используют для введения трансгенов в условиях in vivo [31]. Рекомбинантные аденовирусные векторы обеспечивают очень высокую экспрессию клонированных генов, но на короткое время (5–10 сут) — из-за иммунного ответа организма-реципиента [32]. Для решения этой проблемы было создано второе поколение аденовирусных векторов, у которых дополнительно к гену Е1 удалили гены, отвечающие за репликацию вируса, и оставили только элементы, определяющие начало и конец генома и вирусную пакующую последовательность [33]. Такие векторы способны к более длительной экспрессии генов [14].
Аденовирусы способны инфицировать большинство типов клеток (делящиеся и неделящиеся). Объем для вставки целевого гена составляет 20 тыс. пар оснований, что является достаточно большой емкостью рекомбинантных аденовирусных векторов. Аденовирусы реплицируются в ядре инфицированной клетки как эписомные элементы и обладают высокой эффективностью трансдукции. Так, например, после прямой внутричерепной инъекции рекомбинантного аденовируса была обнаружена его способность заражать нейроны, астроциты, олигодендроглию, эпендимоциты, хориоидальный эпителий и микроглию [34, 35].
Векторные системы на основе вируса простого герпеса
Вирусные векторы на основе вируса простого герпеса (herpes simplex virus, HSV) имеют конструкцию проще, чем векторы на основе аденовирусов. Сам вирус включает около 80 генов, один из которых (IE3) чаще всего замещается при создании вектора [36]. Исключены могут быть и другие гены, что позволяет увеличить объем вектора или заклонировать несколько генов интереса. Недостатками векторов на основе вирусов простого герпеса являются кратковременная экспрессия клонированных генов, токсичность для клеток-мишеней, низкая эффективность трансдукции и способность заражать только неделящиеся клетки [37].
Для создания вирусных векторов сейчас активно используются ампликоны — многократно повторяющиеся последовательности вируса простого герпеса, которые включают мономерные последовательности, организованные как конкатемеры [38]. Каждый мономер включает хотя бы один участок начала репликации вирусной ДНК (oriS или oriL) и последовательность для упаковки ДНК в вирусную частицу (pac) [39]. Молекулярное клонирование этих последовательностей в бактериальную плазмиду позволяет получить вектор, который упаковывает ДНК в вирион HSV. Такие векторные системы способны включать до 150 тыс. пар оснований чужеродной ДНК, что предоставляет возможность одним вектором доставлять в клетку-мишень несколько транскрипционных единиц, не вызывая при этом иммунного ответа и цитопатического эффекта [40]. Особенность жизненного цикла вирусов простого герпеса — наличие двух стадий инфицирования: активной (проникновение и репродукция в клетках эпителия, приводящие к лизису клеток) и латентной (внедрение и репродукция в нервных клетках) [41]. Лизис клеток происходит за счет того, что при репликации вируса сформировавшиеся вирионы покидают клетку путем почкования ядерной мембраны.
Вирусы простого герпеса — нейротрофичны и высокоэффективны при изучении ретроградного и антероградного транспорта в ЦНС, могут быть введены в неопасном латентном состоянии. HSV-векторы имеют большую генетическую емкость и могут обеспечить долгосрочную экспрессию трансгена, однако, как отмечалось выше, основным недостатком их являются токсичность для клеток и низкая эффективность трансдукции [24].
Векторные системы на основе поксвирусов
Поксвирусы — крупные вирусы, которые содержат двунитевую ДНК. Поксвирусный вектор позволяет включить до 25 тыс. пар нуклеотидов ДНК интереса без исключения генов самого вируса. Векторные системы на основе поксвирусов не имеют такого широкого применения, поскольку эукариотические промоторы неэффективно распознаются транскрипционными механизмами поксвирусов и для эффективной экспрессии рекомбинантных генов в клетке-реципиенте нужно использовать поксвирусные промоторы [42]. Поксвирусные транскрипты не подвергаются сплайсингу, из-за чего ДНК интереса обязательно должна быть представлена в форме комплементарной ДНК. Особенностью жизненного цикла поксвирусов является наличие собственной ДНК-зависимой РНК-полимеразы, которая обеспечивает считывание более половины вирусного генома в течение начальной и ранней стадий репродуктивного цикла. Вследствие большого размера и неинфекционной природы поксвирусной ДНК чужеродные гены клонируются в поксвирусах путем рекомбинации в условиях in vivo. Поксвирусы обладают природным тропизмом к опухолевой ткани [43].
Главными недостатками поксвирусов являются высокая иммуногенность, кратковременная продолжительность экспрессии и сложности репликации в эукариотических клетках. При этом положительными сторонами поксвирусов служат высокая эффективность трансдукции, емкость и способность заражать большинство типов клеток [44].
Аденоассоциированные вирусные системы и их преимущества
Рекомбинантные аденоассоциированные вирусы (ААВ) являются одними из наиболее перспективных векторов доставки для генной терапии и нейробиологии благодаря своим непатогенным свойствам, низкой иммуногенности со стороны хозяина, тропности к большинству клеток и тканей, высокой эффективности трансдукции и продолжительной экспрессии [45]. Основной их недостаток — небольшая емкость вектора (до 5 тыс. пар оснований).
Всего у ААВ известно 8 различных серотипов. ААВ2, ААВ3 и ААВ5 первоначально были выделены у обезьян [46]. Полагают, что ААВ6 был сформирован путем рекомбинирования ААВ2 и ААВ1, а именно 5’-конец ААВ2 слился с 3’-концом ААВ1 (включая две открытые рамки считывания) [47, 48]. ААВ7 и ААВ8 были выделены от макаки-резуса при ПЦР-амплификации с использованием праймеров, полученных из консервативных областей серотипов ААВ1–ААВ6 [49]. Среди всех восьми серотипов ААВ2 является наиболее изученным и широко используется в качестве вектора доставки генов [5].
ААВ принадлежит к семейству Parvoviridae. Это группа безоболочечных вирусов, содержащих одноцепочечную ДНК. Вирусная частица имеет икосаэдрическую (20 граней) симметрию с диаметром 18–26 нм и молекулярную массу 5,5–6,2 МДа [50].
Наиболее характерными особенностями ядерной структуры являются группы тройных выступов и выступающих петель, расположенных между двумя смежными субъединицами [51]. Положительно заряженные группы, расположенные вдоль одной стороны каждого выступа, отвечают за связывание с клеточным рецептором — гепарансульфат протеогликаном (HSPG) [52]. В связи с высокой экспрессией данного рецептора в разных тканях связывание с HSPG определяет широкий спектр клеточной специфичности к ААВ2-инфекции [53]. Петля, расположенная в соседней субъединице, также участвует в связывании с клеточным рецептором HSPG и содержит эпитоп для нейтрализующих антител [5]. Нейтрализация иммунного ответа определяет возможность будущего успешного повторного попадания вирусных векторов. Анализ структуры вирусных частиц ААВ2 с помощью рентгеновского излучения открыл возможность изменения эпитопа, содержащегося в вирусных белках капсида, и нейтрализующего антитела и осуществления других модификаций, применимых для реадмиссии в генной терапии [54].
ААВ2 имеет одноцепочечную геномную ДНК, состоящую из 4680 оснований. Инвертированные концевые повторы, состоящие из 145 оснований с высоким содержанием пар GC и способные образовывать палиндромную структуру, расположены как на 5’-конце, так и на 3’-конце [55]. ААВ2-геном синтезируется с мРНК, начиная от промотора p19 [56]. Ген cap кодирует три вирусных белка: VP1 (735 аминокислот, 90 кДа), VP2 (598 аминокислот, 72 кДа) и VP3 (533 аминокислот, 60 кДа), образующих вирусный капсид в соотношении 1:1:20 соответственно, при этом капсид имеет икосаэдрическую симметрию и состоит из 60 субъединиц [57]. Три капсидных белка синтезируются с одной мРНК, начиная с промотора р40, и отличаются друг от друга благодаря альтернативному сплайсингу и разным кодонам инициации [58]. Все три белка капсида имеют одни и те же карбоксиконцевые последовательности и стоп-кодоны (рис. 1).
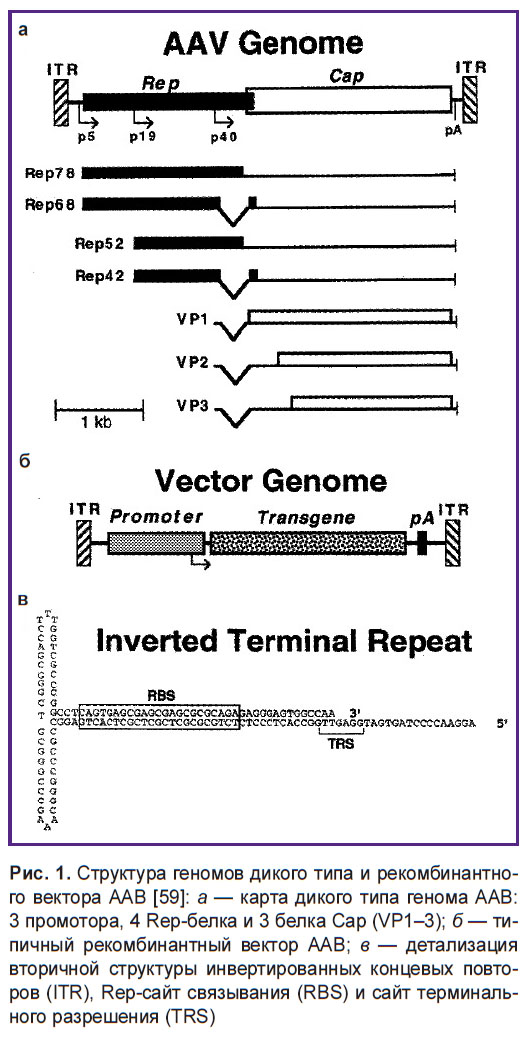 | Рис. 1. Структура геномов дикого типа и рекомбинантного вектора ААВ [59]: а — карта дикого типа генома ААВ: 3 промотора, 4 Rep-белка и 3 белка Cap (VP1–3); б — типичный рекомбинантный вектор ААВ; в — детализация вторичной структуры инвертированных концевых повторов (ITR), Rep-сайт связывания (RBS) и сайт терминального разрешения (ТRS) |
Инвертированные концевые повторы на обоих концах выполняют по крайней мере следующие три функции. Во-первых, 3’-конец служит в качестве праймера для синтеза новой нити ДНК. Во-вторых, он имеет Rep-связывающий сайт (RBS) для Rep78 и Rep68, которые обладают хеликазной, цепьспецифической и сайтспецифической эндонуклеазной активностью [60]. В-третьих, там находится концевой сайт прикрепления (ТRS), который идентичен последовательности в 19-й хромосоме и выступает в качестве последовательности для интеграции с вирусным геномом [61].
Инфекционный путь ААВ2 включает несколько стадий (рис. 2). Вирусные частицы сначала прикрепляются к клеточной поверхности путем связывания с рецептором и корецептором, что в свою очередь ведет к интернализации и включению во внутриклеточный транспорт [62]. Затем эти частицы проникают через ядерную мембрану и либо интегрируют свои геномы с геномом клетки-хозяина, либо реплицируют свой геном внутри ядра [63]. Общепризнано, что основным клеточным рецептором для связывания ААВ2 является HSPG [64].
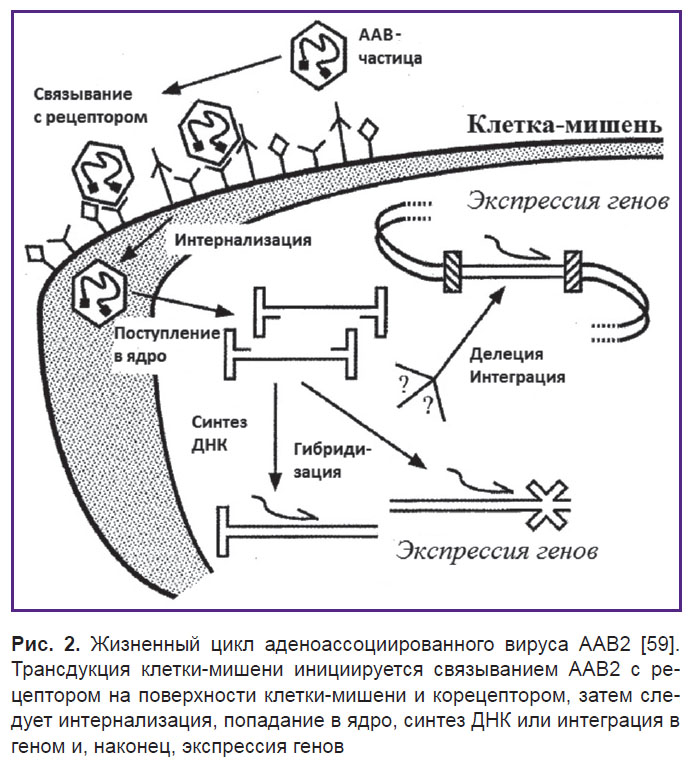 | Рис. 2. Жизненный цикл аденоассоциированного вируса ААВ2 [59]. Трансдукция клетки-мишени инициируется связыванием ААВ2 с рецептором на поверхности клетки-мишени и корецептором, затем следует интернализация, попадание в ядро, синтез ДНК или интеграция в геном и, наконец, экспрессия генов |
Благодаря анализу сайтнаправленного мутагенеза была обнаружена важность основных аминокислот в связывании капсида ААВ2 с гепарином [65].
HSPG выступает в качестве основного рецептора, но ААВ2 также связывается с корецепторами — рецептором 1 фактора роста фибробластов (FGFR1) и αvβ5-интегрином [66, 67]. FGFR1, как полагают, увеличивает степень прикрепления вируса к клетке, в то время как αvβ5-интегрин необходим при эндоцитозе [68, 69]. После эндоцитоза вирусные частицы ААВ2 освобождаются от эндосомы при низком рН [70, 71]. Низкий рН, вероятно, вызывает конформационные изменения вирусных белков, которые играют решающую роль в успешном выходе из эндосомы и проникновении в ядро [72, 73].
В одних ядрах геном ААВ интегрирует в 19-ю хромосому в качестве провируса, а в других вирусные геномы могут оставаться как эписомы [5].
Хелперные плазмиды. Трансфекция
Трансфекция — процесс введения нуклеиновой кислоты в клетки человека и животных невирусным методом. Плазмидную ДНК наиболее часто трансфецируют в клетки, но и другие макромолекулы могут также быть введены внутрь клетки, такие как малые интерферирующие РНК, олигонуклеотиды, РНК и белки [74]. Трансфекция может состоять из нескольких стадий, но обязательным этапом является образование в плазматической мембране пор, через которые внутрь клетки способен проникать внеклеточный материал [75, 76].
Стандартный метод получения рААВ включает в себя котрансфекцию клеток рААВ плазмидой и упаковочной плазмидой, кодирующей гены Rep и белки VP (variol protein) [77]. Кроме того, клетки могут быть коинфицированы вирусом-помощником, например аденовирусом, который обеспечивает выполнение различных функций в производстве ААВ [78, 79]. Как следствие, клетки производят не только ААВ, но и аденовирусы, которые загрязняют конечный продукт (рис. 3, а).
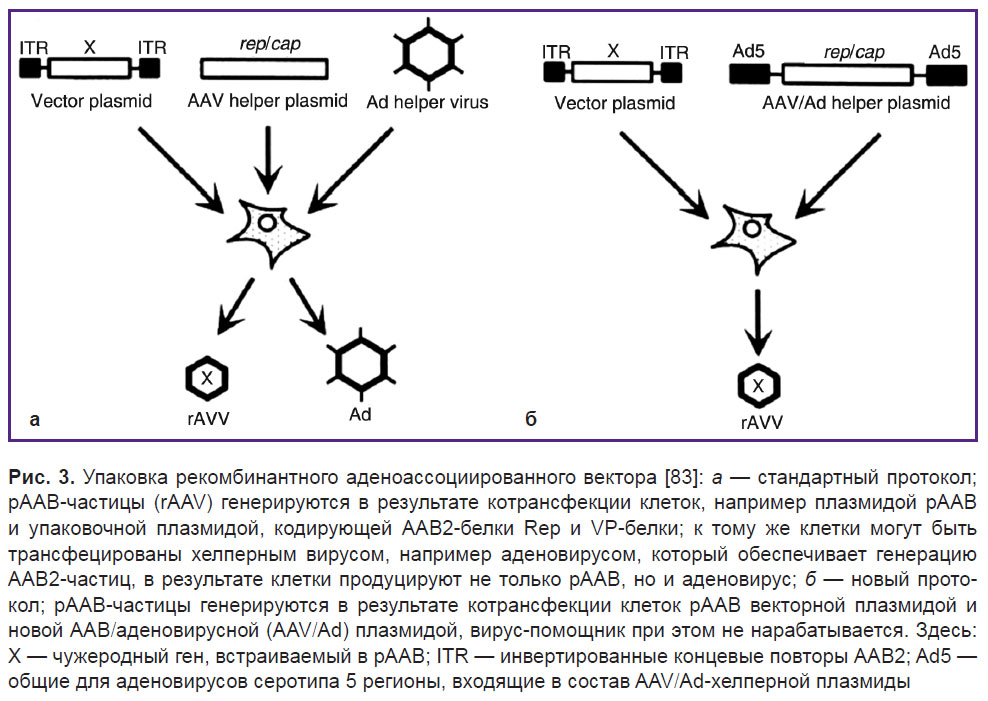 | Рис. 3. Упаковка рекомбинантного аденоассоциированного вектора [83]: а — стандартный протокол; рААВ-частицы (rAAV) генерируются в результате котрансфекции клеток, например плазмидой рААВ и упаковочной плазмидой, кодирующей ААВ2-белки Rep и VP-белки; к тому же клетки могут быть трансфецированы хелперным вирусом, например аденовирусом, который обеспечивает генерацию ААВ2-частиц, в результате клетки продуцируют не только рААВ, но и аденовирус; б — новый протокол; рААВ-частицы генерируются в результате котрансфекции клеток рААВ векторной плазмидой и новой ААВ/аденовирусной (AAV/Ad) плазмидой, вирус-помощник при этом не нарабатывается. Здесь: Х — чужеродный ген, встраиваемый в рААВ; ITR — инвертированные концевые повторы ААВ2; Ad5 — общие для аденовирусов серотипа 5 регионы, входящие в состав AAV/Ad-хелперной плазмиды |
Передовой же метод получения рААВ основан на котрансфекции клеток рААВ и хелперной плазмидой, полученной из ААВ и аденовируса [80–82]. При этом производства аденовируса не происходит (рис. 3, б) [83].
Хелперная плазмида, или плазмида-помощник, обладает одной или несколькими функциями, которые отсутствуют у дефектной плазмиды [84]. Трансформированные клетки должны быть инфицированы также вспомогательным вирусом, например аденовирусом, что обеспечивает множество необходимых преимуществ для производства рААВ [85–87].
Типичные методы производства рААВ-векторов требуют котрансфекции клеток плазмидным вектором ААВ2 и вспомогательной плазмидой (хелперной) [88].
Хелперная плазмида экспрессирует гены rep и cap, а также гены, кодирующие продукты, важные для производства ААВ2 (рис. 4). Простой котрансфекции клеток с хелперной плазмидой и ААВ2-вектором достаточно для успешной наработки рААВ [83, 89].
Рекомбинантные векторы, полученные из ААВ2, — перспективный инструмент для генной терапии и нейробиологии, так как они являются непатогенными и могут долгосрочно экспрессироваться в различных тканях [91–93].
Основные свойства отдельных вирусных векторов, такие как размер вирусных частиц, размер генома и емкость вставки, иммуногенность, а также эффективность трансдукции и экспрессии вируса в клетках-мишенях, отражены в таблице (см. таблицу).
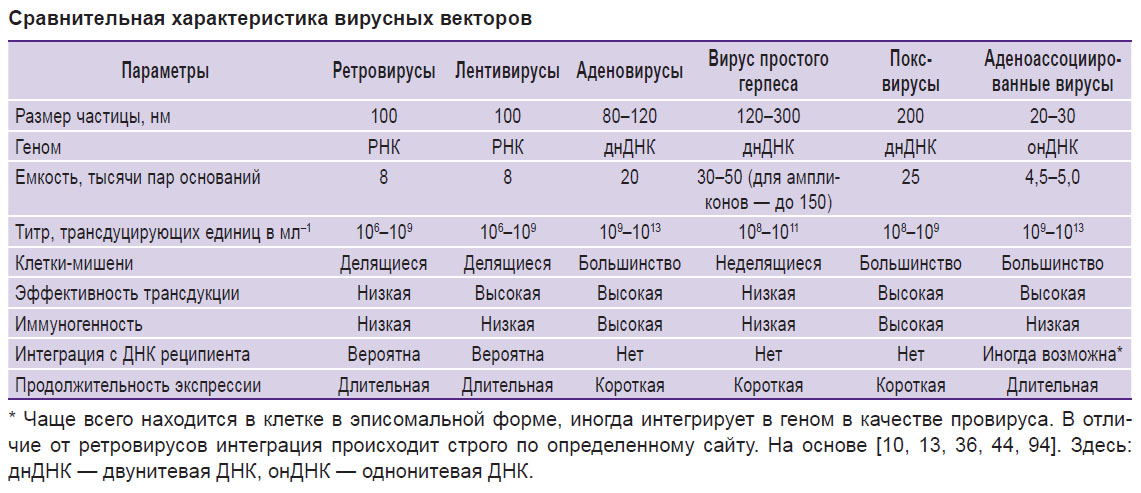 Сравнительная характеристика вирусных векторов Сравнительная характеристика вирусных векторов |
Применение генетических вирусных векторов в нейробиологии
Перенос генетической информации в клетки центральной нервной системы имеет большие перспективы для фундаментальных и клинических исследований в нейробиологии [95, 96]. Так как мозг представляет собой сложноорганизованную структуру, это делает затруднительным генетические манипуляции в нем [97]. В настоящее время вирусные векторы являются наиболее широко используемыми транспортными средствами для переноса генов в зрелые клетки мозга млекопитающих [98, 99]. Тем не менее не существует «универсального идеального вектора» и любые исследования мозга требуют использования определенных векторных систем [100–102]. Кроме того, все вирусные векторные системы имеют свои недостатки и преимущества в зависимости от выбранных клеток-мишеней и специфики каждого исследования [103]. Причем количество векторов, подходящих для фундаментальных исследований, превосходит количество тех, которые могут применяться в клинических испытаниях [104, 105]. Сейчас наиболее широко используемыми векторами в нейробиологии являются аденоассоциированные, аденовирусные, герпесвирусные и лентивирусные векторы [106].
Рекомбинантные ААВ представляют собой векторы нескольких серотипов со специфическим клеточным тропизмом. Например, AАВ2 заражает преимущественно нейроны, но не все виды нейронов заражаются одинаково хорошо. Другие серотипы, такие как ААВ4 и ААВ5, демонстрируют различный тропизм и способность к диффузии [25].
Cre-рекомбиназа в сочетании с Cre-зависимой экспрессией белков и рААВ используется для достижения экспрессии трансгена в различных нейронных популяциях. При этом Cre-зависимые векторы показали надежную способность к ретроградной инфекции и экспрессии трансгена [82].
Генетически кодируемые нейрональные сайленсеры используются для проверки участия нейрональных популяций в поведенческих функциях [107]. Например, соматостатин-экспрессирующие клетки (SST) в комплексе Бетцингера (структура заднего мозга, которая регулирует дыхание) селективно выключались, чтобы проверить их участие в дыхании [108]. В этом исследовании использовались G-белки, связывающиеся с рецепторами аллатостатина (AlstR) дрозофилы, которые могут заставить «замолчать» нейроны млекопитающих при образовании комплекса рецептор–лиганд путем активации проводимости калия. Отдельно лиганд и его рецептор, как полагают, не могут повлиять на нейроны млекопитающих. Использование рААВ2 позволило добиться специфической экспрессии AlstR в субпопуляции клеток SST.
Оценка электрической активности нейронов широко исследовалась для изучения функции нейронов, а в последнее время — для характеристики нейронных сетей. Мониторинг активности нейрона — сложная задача, которая чаще всего сводится к крупномасштабной, мультинейронной визуализации уровня кальция, отражающего электрическую активность [109]. Рекомбинантные ААВ могут доставить генетически закодированные индикаторы кальция для визуализации динамики кальция в естественных условиях с достаточными уровнями экспрессии для эффективного обнаружения. Так, рААВ-экпрессия кальциевого индикатора camgaroo-2 под контролем тетрациклинового трансактиваторного промоторного элемента была осуществлена в обонятельной луковице трансгенных мышей CaMKIIa-tTA [110]. Это вызвало экспрессию в обонятельных сенсорных аксонах и позволило контролировать динамику кальция в ответ на различные ароматические вещества.
Другой кальциевый индикатор GCaMP3 был экспрессирован в клетках моторной коры мыши с использованием рAAВ2/рААВ5, что позволило контролировать динамическую активность нескольких нейронов у бегущей мыши [111].
Также ААВ применяются для выявления и оценки синаптических связей между нейронами разных типов. К примеру, такой подход был использован, чтобы отличить проекционные модели двух смешанных нейронных популяций, которые различаются по экспрессии типов дофаминовых рецепторов 1 или 2 (D1R или D2R) [109]. Применение Cre-зависимого вирусного вектора и сильного промотора EF-1α позволило управлять GFP-трансгеном в сочетании с наличием генетической модификации в мышах, что раскрыло поразительно четкие различия стрионигральной и стриопаллидарной субстанций [111].
Ампликоны вируса простого герпеса применяются для экспрессии генов большого размера или генов с повышенной копийностью. Так, например, мутантная форма торсина А, который ответственен за развитие торсионной дистрофии и формирует витые включения в цитоплазматической мембране [112], может быть экспрессирована в культуре клеток глиомы с помощью ампликона вируса простого герпеса [113].
В лабораторной практике применяются лентивирусные векторы с синапсиновым промотором, способствующим экспрессии заданного гена в любых типах нейрональных клеток [114–117]. Также используются лентивирусные векторы с промотором, содержащим кальций-кальмодулин-зависимую протеинкиназу II альфа, с экспрессией в возбуждающих глутаматергических нейронах [118, 119]. Применяются данные векторы для оптического и электрофизиологического мониторинга их активности, а также для оптогенетической стимуляции, например при остром паркинсонизме, индуцированном посредством медикаментов.
Аденовирусные векторы также применяются в нейробиологии, например в оптогенетике. Так, лабораторией S. Kasparova был создан оптогенетический активатор, который управляет астроцитарной активностью через экспрессию разных вариантов чанелродопсина-2 (ChR2), с целью запуска внеклеточных событий, таких как внеклеточное увеличение концентрации ионов Ca 2+ [120].
Заключение
На данный момент не существует универсального «идеального» вектора, и разные исследования требуют использования определенных векторных систем. У всех вирусных векторных систем будут свои недостатки и преимущества в зависимости от выбираемых клеток-мишеней и специфики каждого исследования. В частности, преимущества рекомбинантных аденоассоциированных векторов (способность интегрировать целевой ген в геном хозяина в нужное место, что предотвращает нежелательные мутации; встраивание как в делящиеся, так и в покоящиеся клетки; широкий профиль трансдукции; низкий иммунный ответ; сильная и устойчивая экспрессия трансгена) выделяют их среди остальных вирусных векторов и делают эти векторы популярным и универсальным инструментом для доставки гена in vitro и in vivo.
Финансирование исследования. Работа выполнена при поддержке Федеральной целевой программы «Исследования и разработки по приоритетным направлениям развития научно-технологического комплекса России на 2014–2020 годы» Министерства образования и науки России, соглашение 14.581.21.0016 («Разработка методов, технологий и платформ для исследований функционирования нервных систем на основе создания высокоразрешающей информационной модели кортикальных структур мозга», уникальный идентификатор ПНИЭР RFMEFI58115X0016).
Конфликт интересов. Авторы декларируют отсутствие конфликта интересов.



