Зоопсихология: конспект лекций.
6.3. Перцептивная психика. Проблема интеллекта у животных.
Низший уровень развития перцептивной психики. Перцептивная психика является высшей стадией развития психического отражения. Эта стадия психического развития уже характеризуется наличием подлинных навыков и восприятий. Компоненты среды отражаются организмом как целостные единицы, тогда как на предыдущем уровне развития отражались лишь отдельные свойства или сумма предметных компонентов среды. Именно на этой стадии развития психики появляются чувственные представления. Сама перцептивная психика, которая наблюдается у множества живых организмов, обнаруживает большие различия. Поэтому возникла необходимость провести более дробную классификацию, согласно которой первый уровень развития перцептивной психики назван низшим.
Низший уровень развития перцептивной психики характерен в первую очередь для высших беспозвоночных – головоногих моллюсков и членистоногих. Среди членистоногих характеристику этого уровня психического развития лучше всего рассмотреть на примере насекомых – самого многочисленного класса членистоногих.
Специфический образ жизни, различные формы двигательной активности и разнообразие качественно различных агентов среды, управляющих поведением, обусловили развитие у насекомых многочисленных и своеобразно устроенных органов чувств. Среди них наиболее важен зрительный аппарат, так как именно хорошо развитое зрение способствовало оптическому восприятию форм как необходимому компоненту перцептивной психики. Следует помнить, что на уровне элементарной сенсорной психики различение форм животным еще невозможно.
До недавнего времени считалось, что насекомые способны к восприятию формы, но лишь в специфических рамках. В первых экспериментах было показано, что пчелы могут воспринимать только те объекты, которые отдаленно напоминают по своему строению цветок (круги, звездочки). Но позднее, в экспериментах советского зоолога Мазохина-Поршнякова, было доказано, что пчел изначально можно дрессировать и на восприятие непривычных для них фигур, например треугольника или круга, в результате чего был сделан вывод, что пчелы способны распознавать фигуры непосредственно по их графическим признакам.
Подобные опыты на одиночных осах проводил и один из основателей современной этологии Н. Тинберген. Он обучал самок ос распознавать круг из сосновых шишек, выложенных вокруг входа в норку. После того как оса улетала за добычей, круг передвигался на 30 см в сторону. Возвратившись, оса сперва искала норку в центре круга. В следующих опытах (помимо передвижения круга) шишки заменяли черными камешками, а вокруг норки выстраивали треугольник или даже эллипс из этих камешков, но оса тем не менее прилетала в круг, хотя из предыдущих опытов было известно, что она вполне способна отличить камешки от шишек. Таким образом, пространственная ориентация осуществлялась здесь только по форме (кругу).
Способность к предметному восприятию у высших насекомых заметно ниже, чему позвоночных, что можно объяснить специфическим строением органов зрения. Кроме того, насекомые в большей степени ориентируются не по предметным компонентам среды, а по их отдельным признакам, что в большей степени характерно для стадии элементарной сенсорной психики.
Едва ли не более важную роль, чем у насекомых, зрение играет и у головоногих моллюсков. Для них зрение является ведущей рецепцией, на что указывают сложное строение и крупные размеры глаз. Относительные размеры глаз кальмаров превышают относительные размеры глаз большинства водных млекопитающих (китов, дельфинов) в десятки раз. Поразительна и огромная разрешающая способность (зоркость) глаза головоногих: на 1 мм2у разных представителей головоногих приходится от 40 до 162 тысяч палочек, у человека – 120–400 тысяч, у совы, обладающей самым зорким глазом в мире, – 680 тысяч.
Головоногие моллюски способны к подлинному предметному восприятию, что выражается прежде всего в различении ими формы объектов. Это было доказано в опытах Б.Б. Бойкотта и Дж. З. Янга. Оказалось, что осьминоги могут не только воспринимать форму объектов, но и различать их относительный размер, а также положение в пространстве (например, отличали вертикальный прямоугольник от горизонтального). Всего эти головоногие моллюски различали более 46 различных форм.
У высших беспозвоночных появляется уже зачатки общения, которое особенно развито у животных, ведущих групповой образ жизни (пчелы, муравьи). Именно у этих насекомых появилась возможность передавать информацию с помощью специальных сигнальных действий. Весьма выражено у беспозвоночных и территориальное поведение. Его зачатки можно обнаружить уже у дождевых червей. У высших беспозвоночных хорошо выражены маркировка индивидуального участка, своеобразное совмещение территориального поведения и передачи информации.
Уже на низшем уровне развития перцептивной психики представлены все те прогрессивные признаки, которые характеризуют перцептивную психику вообще, но во многих отношениях поведение относящихся сюда животных носит и примитивные черты, сближающие его с поведением нижестоящих животных. Поведение по-прежнему ориентировано на отдельные свойства предметов, предметное восприятие выражено слабо. В поведении преобладают жестко запрограммированные элементы, в нем очень мало гибкости. Одновременно с этим на данном уровне развития психики появляется четко выраженный активный поиск положительных раздражителей, таксисное поведение получает мощное развитие. Налицо все виды высших таксисов, включая мнемотаксисы. Именно мнемотаксисы играют важную роль в пространственной ориентации, а в заучивании ориентиров уже проявляется способность к изменению поведения, т. е. к научению.
Хотя у беспозвоночных, в частности насекомых, накопление индивидуального опыта, научение играют существенную роль, наблюдаются и определенная противоречивость в процессах научения, сочетание прогрессивных и примитивных черт. Хорошо заметен переходный этап между инстинктивным поведением и истинным научением, что и помещает данный уровень развития психики между элементарной сенсорной и развитой перцептивной психикой.
Само инстинктивное поведение представлено уже развитыми новыми категориями, такими как групповое поведение, общение. На современном этапе развития науки лучше всего изучен язык пчел, доказано, что у этих насекомых хорошо развиты сложные формы общения. Самые сложные формы инстинктивного поведения закономерно сочетаются у них с наиболее разнообразными и сложными проявлениями научения, что обеспечивает не только исключительную согласованность действий всех членов пчелиной семьи, но и максимальную пластичность поведения особи. Психические способности пчел (как и некоторых других высших насекомых) в некоторых отношениях, очевидно, уже выходят за рамки низшего уровня перцептивной психики.
На низшем уровне перцептивной психики находится и ряд представителей низших позвоночных. Основная причина этого – их относительно малые размеры. Все беспозвоночные живут в условиях (температура, освещение), в корне отличных от условий обитания крупных позвоночных. Уже поэтому психическое отражение действительности у насекомых, как и у большинства других беспозвоночных, не может не быть принципиально иным, чем у позвоночных. По общим признакам психического отражения, свойственного данному уровню, можно сделать вывод, что у насекомых наблюдается типичное проявление низшего уровня перцептивной психики, но в формах, отвечающих тем особым условиям жизни этих животных, о которых говорилось выше.
Высший уровень развития перцептивной психики. Доказано, что в ходе эволюционного процесса в животном мире образовались как бы три отдельно стоящие вершины: позвоночные, насекомые и головоногие моллюски. Все эти группы довольно рано отмежевались от общего эволюционного ствола и независимо друг от друга достигли вершин развития. Именно у этих животных наблюдаются наиболее сложные формы поведения и психического отражения, обусловленные высоким развитием уровня строения и жизнедеятельности. Представители всех названных групп способны к предметному восприятию, но лишь у позвоночных оно получило полное развитие. Неудивительно, что высшего уровня развития перцептивной психики в ходе эволюции достигли только позвоночные, да и то далеко не все представители этого типа. Лишь у высших позвоночных обнаруживаются все наиболее сложные проявления психической деятельности, которые вообще встречаются в мире животных.
Высокое развитие психической деятельности позвоночных напрямую связано с усложнением их организации, разнообразием движений, усложнением строения нервной системы и органов чувств. Все основные проявления психической деятельности, свойственной животным, описанные в других разделах книги, характерны именно для позвоночных. Рассмотрим наиболее важные из этих проявлений.
В первую очередь это манипулирование. Конечности животных, изначально выполнявшие только опорную и локомоторную функции, по мере развития получили ряд дополнительных функций, одной из которых и является манипулирование. Для зоопсихолога особый интерес представляет манипулирование передними конечностями, которое в итоге привело к возникновению орудийной деятельности приматов и послужило биологической предпосылкой зарождения трудовых действий у древнейших людей. Манипулирование свойственно в основном приматам, заметно реже оно наблюдается у представителей других отрядов млекопитающих. При манипулировании животное всесторонне знакомится с предметом, больше узнает о его свойствах. При соответствующих условиях животные получают наиболее всеобъемлющую и разнообразную информацию, необходимую для развития высших форм психической активности. Оказалось, что медведи владеют тремя способами фиксации объекта на весу, еноты – шестью, низшие обезьяны и полуобезьяны – тремя десятками таких способов! К тому же только обезьяны обладают различными двигательными возможностями, достаточными для того, чтобы произвести подлинный деструктивный анализ (расчленение) объекта на весу. Разновидностью манипулирования является и комфортное поведение, хорошо развитое у многих высших позвоночных.
На данном этапе развития перцептивной психики получили свое развитие также зрительные обобщения и формирование представления. Известно, что подлинное восприятие предметных компонентов среды возможно только на основе способности к анализу и обобщению, поскольку только таким способом можно узнавать постоянно меняющиеся компоненты среды. Все позвоночные начиная с рыб способны к предметному восприятию, в частности к восприятию форм. Высшие позвоночные способны к обобщению, т. е. в опытах они узнают объект, если он не только поменял свое место, но и изменил положение в пространстве. Например, млекопитающие могут быстро узнавать треугольники разного размера и разным образом ориентированные в плоскости. При соответствующем научении высшие позвоночные в состоянии даже в очень сложных ситуациях вычленять существенные детали в воспринимаемых объектах и узнавать эти объекты в сильно измененном виде. Отсюда напрашивается вывод о наличии у позвоночных достаточно сложных общих представлений.
Наличие у позвоночных представлений, выражающихся в отсроченных реакциях, и способности к нахождению обходных путей (включая явления экстраполяции), придает их поведению исключительную гибкость и намного повышает эффективность их действий на поисковых этапах поведенческих актов. Однако способность к обобщению не указывает на высокий уровень психического развития организма. Эта способность служит в первую очередь предпосылкой к развитию сложных навыков, которые составляют основное содержание накопления индивидуального опыта не только в сенсорной, но и в эффекторной сфере деятельности организма.
У высших позвоночных заметно усложняются и процессы общения. У них очень разнообразны средства коммуникации, которые включают в себя элементы различной модальности, например ольфакторные, тактильные. Ольфакторная коммуникация досталась им в наследство от территориального поведения, когда животные активно маркировали границы собственных участков.
Компоненты инстинктивного поведения позвоночных, служащие для общения, в той или иной степени ритуализованы. Оптическое общение осуществляется с помощью характерных поз, телодвижений, которые заметно упрощены и имеют четкую последовательность действий. В первую очередь они служат для биологического разграничения видов и в большей степени выражены у близкородственных видов. Конкретные формы оптического общения у высших позвоночных отличаются большим разнообразием и дифференцированностью. У млекопитающих оптическое общение нередко сочетается с ольфакторным, выделение систем общения по отдельным модальностям у этих животных в большой степени условно. В той или иной степени это касается и акустических сигналов, которые у млекопитающих зачастую сопровождаются характерными позами. Наиболее развита звуковая сигнализация у птиц, она охватывает практически все сферы их жизнедеятельности. Большое значение имеют не только четкие межвидовые различия акустического общения, но и различия индивидуальные, по которым особи узнают друг друга.
Таким образом, можно сказать, что на высшем уровне развития перцептивной психики формируются все основные формы поведения животных, а более древние из этих форм, возникшие на ранних стадиях эволюции психики, достигают своего высшего развития.
Сложные навыки представляют собой исключительно динамические моторно-рецепторные системы, обеспечивающие на основе высокоразвитой ориентировочной деятельности выработку весьма пластичных двигательных программ. У высших животных наблюдается слияние ориентировочного процесса с двигательной активностью, и принятие верных решений в меняющихся условиях среды производится на основе высокоразвитого чувственного обобщения. Подобные сложные навыки, свойственные высшим позвоночным, стали предпосылками для развития высших форм психической деятельности животных – интеллектуальных действий.
Проблема интеллекта животных. Общепризнано, что интеллектуальное поведение является вершиной психического развития животных. В ходе многочисленных экспериментов доказано, что интеллектуальная деятельность характерна только для высших позвоночных, но, в свою очередь, не ограничивается одними приматами. Следует помнить, что интеллектуальное поведение животных является не чем-то обособленным, из ряда вон выходящим, это лишь одно из проявлений единой психической деятельности с ее врожденными и благоприобретаемыми аспектами. По мнению К. Фабри, «. интеллектуальное поведение не только теснейшим образом связано с разными формами инстинктивного поведения и научения, но и само складывается (на врожденной основе) из индивидуально-изменчивых компонентов поведения. Оно является высшим итогом и проявлением индивидуального накопления опыта, особой категорией научения с присущими ей качественными особенностями. Поэтому интеллектуальное поведение дает наибольший приспособительный эффект. при резких, быстро протекающих изменениях в среде обитания».[29]
Основной предпосылкой развития интеллекта является манипулирование. В первую очередь это относится к обезьянам, для которых данный процесс служит источником наиболее полных сведений о свойствах и структуре предметных компонентов среды. В ходе манипулирования, особенно при выполнении сложных манипуляций, обобщается опыт деятельности животного, формируются обобщенные знания о предметных компонентах окружающей среды, и именно этот обобщенный двигательно-сенсорный опыт составляет главнейшую основу интеллекта обезьян. При манипулировании животное получает информацию одновременно по ряду сенсорных каналов, но преобладающее значение у обезьян имеет сочетание кожно-мышечной чувствительности рук со зрительными ощущениями. Кроме того, в обследовании объекта манипулирования участвуют обоняние, вкус, тактильная чувствительность околоротовых вибрисс, иногда слух. Животные получают комплексную информацию об объекте как о едином целом, обладающем разнокачественными свойствами. Именно в этом и заключается значение манипулирования как основы интеллектуального поведения.
Первостепенное значение для интеллектуального поведения имеют зрительные обобщения, также хорошо представленные у высших позвоночных. По экспериментальным данным, кроме приматов, зрительное обобщение хорошо развито у крыс, некоторых хищных млекопитающих, из птиц – у врановых. У этихживотных зрительное обобщение зачастую близко к абстрагированию, свойственному мыслительным процессам.
Другой элемент интеллектуального поведения, направленный в двигательную сферу, подробно изучается у позвоночных с помощью метода проблемного ящика. Животные вынуждены решать сложные предметные задачи, находить последовательность отпирания различных запоров и задвижек, чтобы выбраться из клетки или добраться до лакомства. Доказано, что высшие позвоночные решают предметные задачи гораздо хуже, чем задачи, основанные на применении локомоторных функций. Это можно объяснить тем, что в психической деятельности животных преобладает познавание пространственных отношений, постигаемых ими с помощью локомоторных действий. Только у обезьян и некоторых других млекопитающих за счет развития манипуляционной деятельности прекращают доминировать локомоторные действия, животные легче абстрагируются и соответственно лучше решают предметные задачи.
Важной предпосылкой интеллектуального поведения, по мнению К. Фабри, является способность к широкому переносу навыков в новые ситуации. Эта способность вполне развита у высших позвоночных, хотя и проявляется у разных животных в разной степени. Основные лабораторные эксперименты в данном направлении проводились на обезьянах, собаках и крысах. По словам К. Фабри, «способности высших позвоночных к разнообразному манипулированию, кширокому чувственному (зрительному) обобщению, к решению сложных задач и переносу сложных навыков в новые ситуации, к полноценной ориентации и адекватному реагированию в новой обстановке на основе прежнего опыта являются важнейшими элементами интеллекта животных. И все же сами по себе эти качества еще недостаточны, чтобы служить критериями интеллекта, мышления животных».[30]
Какими же основными критериями обладает интеллектуальное поведение животных? Одной из основных особенностей интеллекта является то, что при этой деятельности в дополнение к обычному отражению предметов возникает также отражение их отношений и связей. В зачаточных формах это было представлено еще при формировании сложных навыков. Любое интеллектуальное действие состоит как минимум из двух фаз: фазы подготовки действия и фазы осуществления действия. Именно наличие фазы подготовки является характерной чертой интеллектуального действия. По мнению А.Н. Леонтьева, интеллект впервые возникает там, где возникает процесс подготовки возможности осуществить ту или иную операцию или навык.
В ходе эксперимента можно четко разграничить основные фазы интеллектуального действия. Например, обезьяна берет палку и в следующее мгновение с ее помощью пододвигает к себе банан, или же она предварительно строит пирамиду из пустых ящиков, чтобы сорвать с веревки подвешенную под потолком приманку. Н.Н. Ладыгина-Котс детально изучала у шимпанзе процесс подготовки и даже изготовления орудия, необходимого для решения технически несложной задачи – выталкивания приманки из узкой трубки. На глазах у шимпанзе в трубку закладывалась приманка таким образом, что ее нельзя было достать просто пальцами. Одновременно с трубкой животному давали различные предметы, пригодные для выталкивания корма. После того как проводилось некоторое усовершенствование предмета, используемого для доставания корма, подопытная обезьяна вполне (хотя и не всегда немедленно) справлялась со всеми поставленными задачами.
Во всех этих опытах хорошо видны две фазы интеллектуального действия: первая, подготовительная фаза – подготовка орудия, вторая фаза – доставание приманки с помощью этого орудия. Первая фаза вне связи со следующей фазой лишена какого бы то ни было биологического смысла. Вторая фаза – фаза осуществления деятельности – в целом направлена на удовлетворение определенной биологической потребности животного (в описанных опытах – пищевой).
Еще одним важным критерием интеллектуального поведения является то обстоятельство, что при решении задачи животное пользуется не одним стереотипно выполняемым способом, а пробует разные способы, которые являются результатом ранее накопленного опыта. Животные пытаются произвести не различные действия, а различные операции и в конечном счете могут решить задачу разными способами. Например, из ящиков можно построить пирамиду, чтобы сорвать висящий банан, а можно разобрать ящик на части и попытаться сбить лакомство отдельными дощечками. Операция перестает быть неподвижно связанной с деятельностью, отвечающей определенной задаче. Именно этим интеллект заметно отличается от любых, даже самых сложных, навыков. Так как интеллектуальное поведение животных характеризуется отражением не просто предметных компонентов среды, а отражает отношения между ними, здесь осуществляется и перенос операции не только по принципу сходства вещей (например, преград), с которыми она была связана, но и по принципу сходства отношений, связей вещей, которым она отвечает.
Несмотря на высокий уровень развития, интеллект млекопитающих, в частности обезьян, имеет четкую биологическую ограниченность. Наравне с другими формами поведения он всецело определяется образом жизни и биологическими закономерностями, за рамки которых животное перешагнуть не может. Это показывают многочисленные наблюдения за человекообразными обезьянами в природе. Так, шимпанзе сооружают довольно сложные плетеные гнезда, в которых проводят ночь, но никогда не строят даже простейших навесов от дождя и во время тропических ливней нещадно мокнут. В природных условиях обезьяны редко пользуются орудиями, предпочитая при необходимости добывать более доступные корма, чем тратить время и силы на добычу труднодоступных.
Ограниченность интеллектуального поведения была показана и в многочисленных экспериментах, проводимых Ладыгиной-Котс над человекообразными обезьянами. Например, самец шимпанзе допускал порой глупые ошибки при употреблении предметов, предоставленных ему для выталкивания приманки из трубы. Он пытался втолкнуть в трубу кусок фанеры вопреки явному несоответствию ее ширины диаметру трубы и принимался обгрызать его лишь после ряда таких неудачных попыток. По мнению Ладыгиной-Котс, шимпанзе «не в состоянии схватить сразу существенные особенности в новой ситуации».[31]
Даже самые сложные проявления интеллекта обезьян представляют собой в конечном итоге не что иное, как применение в новых условиях филогенетически выработанного способа действия. Обезьяны способны притягивать к себе плод с помощью палки только потому, что в природных условиях им часто приходится пригибать ветку с висящим на ней плодом. Именно биологическая обусловленность всей психической деятельности обезьян, включая антропоидов, является причиной ограниченности их интеллектуальных способностей, неспособности к установлению мысленной связи между одними лишь представлениями и их комбинированием в образы. Неспособность мысленно оперировать представлениями приводит обезьян и к неспособности понимать истинные причинно-следственные связи, поскольку это возможно лишь с помощью понятий, которые у обезьян, как и у всех других животных, полностью отсутствуют.
Между тем на данном этапе развития науки проблема интеллекта животных изучена недостаточно. По существу до сих пор проведены обстоятельные экспериментальные исследования только над обезьянами, преимущественно высшими, в то время как возможность интеллектуальных действий у других позвоночных практически не подтверждена доказательными экспериментальными данными. Вместе с тем ошибочно считать, что интеллект присущ только приматам. Скорее всего, объективные исследования будущих зоопсихологов помогут пролить свет на этот непростой, но очень интересный вопрос.
Что такое перцептивная психика




Глава 2. Перцептивная психика
Низший уровень развития перцептивной психики
Разумеется, перцептивная психика, свойственная огромному числу животных, стоящих на разных ступенях эволюционного развития, обнаруживает в конкретных своих проявлениях большие различия. Поэтому необходимо и на этой стадии развития психики выделить отдельные уровни, из которых здесь будет рассмотрен низший.
Огромное разнообразие «экологических ниш», занимаемых насекомыми и другими членистоногими, обусловило, естественно, развитие весьма различных форм приспособления в области строения и поведения этих животных.
Соответственно и крылевая мускулатура насекомых отличается исключительно сложным устройством: она состоит из множества различно располагающихся мышечных пучков, частично действующих как антагонисты. Большую сложность обнаруживают и другие отделы мышечной системы.
Вообще у насекомых (не считая крылевой мускулатуры насекомых) различают следующие отделы мышечной системы: мышцы туловища, заведующие его движениями; мышцы конечностей, с помощью которых движется вся конечность по отношению к туловищу; мышцы отдельных члеников конечностей; мышцы внутренних органов.
Центральная нервная система
В соответствии со сложной и высокодифференцированной организацией двигательного аппарата находится и сложное строение центральной нервной системы насекомых, которую, однако, мы можем здесь охарактеризовать лишь в самых общих чертах.
Как и у кольчатых червей, основу всей нервной системы составляют у насекомых метамерно расположенные ганглии, связанные друг с другом межсегментными нервными стволами. Эти стволы сближаются у большинства насекомых настолько, что образуют (хотя бы внешне) одну непарную нервную цепочку. Одновременно имеет место и слияние последовательных метамерных ганглиев.
Этим обеспечивается значительно большая скорость проведения нервных импульсов, чем у низших моллюсков. Соответственно огромной величине глаза исключительно развит и зрительный нерв. Выдающийся советский гистолог А. А. Заварзин показал большое сходство тонкого гистологического строения оптических центров головоногих и позвоночных.
Таким образом, мы видим, что глубокие различия между элементарной сенсорной и перцептивной психикой находят свое воплощение и в структуре нервной системы уже на низшем уровне перцептивной психики.
Сенсорные способности, таксисы
Специфические условия, в которых живут насекомые, далеко идущее приспособление к ним двигательной активности при обилии и разнообразии качественно весьма различных агентов среды, управляющих их поведением,- все это обусловило появление у данных животных многих, и притом своеобразна устроенных, органов чувств.
Благодаря такому устройству сложных глаз в мозг насекомого передаются «мозаичные» изображения воспринимаемых предметных компонентов среды. Конечно, это не означает, что насекомое действительно видит окружающий мир в виде мозаики. В мозгу поступающие из отдельных омматидиев зрительные раздражения синтезируются в целостную картину, как это имеет место и у позвоночных. Все же нам очень трудно представить себе, как на самом деле видит насекомое.
Еще в прошлом веке делались попытки установить путем фотографирования через фасеточные глаза четкость и дальность зрения насекомых и раков. Позже, в частности в 60-х годах нашего века, такие опыты повторялись, но едва ли возможно таким образом решить проблему, ибо, как известно, видит не глаз, а мозг. Насекомые, очевидно, весьма близоруки: ясное видение, например, бабочки крапивницы возможно лишь на расстоянии приблизительно 2-3 сантиметров. От объектов, расположенных дальше (уже начиная с 5-7 сантиметров), насекомые получают лишь расплывчатые изображения, но, очевидно, этого достаточно для их общей ориентировки в пространстве.
На расстоянии пчела способна увидеть лишь объекты в сто раз большей величины, чем это необходимо для зрения человека. Отдельный цветок она увидит лишь тогда, когда она приблизится к нему вплотную. Все эти особенности зрения насекомых вполне соответствуют тому микроландшафту, в котором живут эти животные.
Острота зрения (разрешающая способность, т. е. способность видеть две сближенные точки) зависит у насекомых при прочих равных условиях, в частности, от количества омматидиев, которое, однако, сильно колеблется даже в пределах отрядов. Так, в отряде жуков это количество колеблется от трех-четырех десятков до нескольких десятков тысяч. У пчелы каждый глаз состоит приблизительно из четырех тысяч фасеток. Чем мельче фасетки, тем слабее в них освещение, но тем детальнее зрение. Особенно большое значение это имеет для предметного зрения. Кроме того, при более выпуклом глазе оси омматидиев сильнее расходятся, и в результате расширяется поле зрения. Одновременно суживается точка, видимая одной фасеткой, чем также увеличивается острота зрения.
Наилучшим образом насекомые видят подвижные объекты или неподвижные предметы во время собственного движения. Пролетающая над лугом пчела, например, видит покачивающиеся на ветру цветы или же изображение неподвижного цветка скользит по ее фасеточному глазу. В любом случае это подвижное (расплывчатое) изображение заставляет ее спуститься вниз и обследовать растение с минимального, т. е. оптимального для зрения пчелы, расстояния.
Что касается цветового зрения, то видимый ими спектр шире, чем у человека, так как его граница проходит в ультрафиолетовом секторе. Спорный вопрос о способности насекомых к восприятию красного цвета в настоящее время, очевидно, решен в положительном смысле (за исключением медоносной пчелы и некоторых других насекомых). Советский энтомолог Г. А. Мазохин-Поршняков считает, что вообще цветовое зрение у взрослых насекомых представляет универсальное явление. По его данным, цвета различаются даже видами, активная жизнь которых протекает ночью.
До сих пор мы касались только реакции насекомых на свет, который играет в их жизни очень большую роль. Не меньшее, если не большее, значение имеют для них запахи. В естественном поведении насекомых стимулы различной модальности всегда выступают в комплексе. Так, уже упомянутые осы рода Philanthus охотятся за пчелами, руководствуясь зрением с расстояния 30 сантиметров, но опознают их по запаху лишь с расстояния 15-5 сантиметров. Во время зрительного поиска оса совершенно не реагирует на запах пчелы.
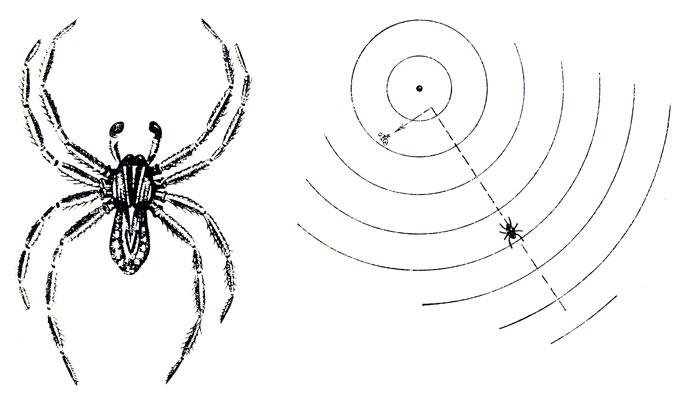
Рис. 36. Водяной паук Pirata бежит по поверхности воды к источнику вибрации (центральная точка). Только непосредственно около неподвижной мухи вид последней приобретает преобладающее влияние и паук сворачивает к ней (опыт Берестынской-Вильчек)
Для ознакомления с таксисами у насекомых рассмотрим в качестве примера фототаксисы. Эти таксисы (положительные или отрицательные) встречаются у насекомых прежде всего в виде тропо-, тело- и менотаксисов, хотя не утратили своего значения и примитивные таксисы. Так, например, положительные и отрицательные клинотаксисы с четко выраженными маятникообразными поисковыми движениями играют ведущую роль в оптической ориентации у многих личинок мух. Тропо- и телотаксисы связаны у них Друг с другом многими переходными формами. У взрослых насекомых чаще всего встречаются, однако, четко выраженные телотаксисы. Как правило, тропотаксисное движение очень скоро переходит у них в телотаксисное. У комаров и вшей преобладает тропотаксисная ориентация движения.
Менотаксисы, относящиеся к более сложным формам ориентации, встречаются у высших насекомых (муравьев, пчел, бабочек, жуков). Немецкий исследователь поведение насекомых М. Линдауер выделяет здесь следующие четыре случая: 1) угол фиксации источника света случаен (муравьи, жук-навозник); 2) угол фиксации является врожденным (некоторые бабочки); 3) угол фиксации сообщается особями одного вида друг другу («танцы» пчел, информирующие об угле ориентации по положению солнца); 4) угол фиксации выучивается в индивидуальном порядке. Последний случай

Рис. 37. Мнемотаксис у роющей осы Philanthus. После отлета осы кольцо из шишек вокруг норки было перемещено в сторону. После возвращения оса ищет норку внутри кольца и находит ее лишь после повторных облетов (опыт Тинбергена)
Необходимо, однако, подчеркнуть, что и в тех случаях, когда ориентация осуществляется на основе других фототаксисов, поведение насекомых одновременно определяется рядом дополнительных факторов. Например, доказано, что у многих насекомых характер их фототаксисного поведения сильно видоизменяется в зависимости от температуры воздуха, времени дня, физиологического состояния и т. д. Хорошо известно, что муха не реагирует на свет, пока спокойно летает по комнате. Но стоит начать ее преследовать, как у нее сейчас же «просыпается» положительный фототелотаксис: она быстро направляется к источнику света, к окну.
Аналогичным образом насекомые реагируют на разнообразные механические, температурные, химические и оптические раздражения, на силу тяжести, и соответственно у. них встречаются во многих различных формах тигмо-, рео-, анемо-, гидро-, гео-, термо-, хемо-, фото- и другие таксисы.
Особый интерес представляет способность насекомых (и других членистоногих) к оптическому восприятию форм как необходимого компонента перцептивной психики. На уровне элементарной сенсорной психики различение форм еще невозможно.
До недавнего времени считалось, что насекомые способны к восприятию формы, но лишь в специфических рамках. Эта специфика усматривалась в том, что в экспериментах насекомые, особенно пчелы, оказывались способными различать лишь фигуры,
напоминающие по своим очертаниям цветы (например, кресты или звездчатые фигуры), простые же геометрические фигуры они в этих опытах не различали. Однако Мазохин-Поршняков (в работах, проведенных совместно с Г. М. Вишневской) показал, что пчел можно с полным успехом дрессировать и на такие фигуры, как треугольник, круг, квадрат, из чего он заключил, что пчелы способны распознавать фигуры непосредственно по их графическим признакам.
О способности насекомых к распознаванию форм свидетельствуют и полевые опыты Тинбергена, в которых самки роющих ос обучались распознавать круг из сосновых шишек, выложенных вокруг входа в норку. После того как оса улетала за добычей, круг передвигался на 30 сантиметров в сторону. После возвращения оса искала норку сперва в центре круга (рис. 37). В следующих опытах к тому же шишки заменялись черными камешками, а вокруг норки выстраивался треугольник или даже эллипс из шишек, но оса тем не менее прилетала в круг, хотя из предыдущих опытов было известно, что оса вполне способна отличать камешки от шишек. Таким образом, пространственная ориентация осуществлялась здесь только по форме (кругу).
Хотя у высших насекомых способность к предметному восприятию и не вызывает сомнения, в этом отношении они явно уступают позвоночным, Очевидно, это связано со специфическим строением органов зрения: разрешающая способность глаза зависит у насекомых от количества омматидиев. Чем их больше, тем выше эта способность, так как картина воспринимаемого объекта расчленяется на большее число точек. К тому же насекомые, как и другие членистоногие, очевидно, ориентируются преимущественно все же не по предметным компонентам среды как таковым, а по отдельным их признакам, как это имеет место на уровне элементарной сенсорной психики.
Общение и групповое поведение
У высших беспозвоночных уже в полном объеме представлена категория поведения, которая, как было показано, только в зачаточной
форме обнаруживается у некоторых представителей низших многоклеточных беспозвоночных. Речь идет об общении, которое у высших беспозвоночных проявляется иногда и в очень сложных формах подлинного группового поведения (в отличие от колониальной жизни, свойственной многим низшим беспозвоночным). Напомним, что общение, в точном значении этого термина, появляется только тогда, когда контакты между животными начинают осуществляться посредством специальных сигнальных действий.
У головоногих моллюсков и членистоногих, особенно у насекомых, мы находим четкие системы коммуникации с передачей информации по разным сенсорным каналам. Это относится прежде всего к пчелам и муравьям, что, конечно, связано со сложной, высокодифференцированной структурой их сообществ и разделением функций между членами последних (распределение пищедобывательной, защитной и воспроизводительной функций между отдельными особями).
У муравьев разные формы химической чувствительности играют, очевидно, большую роль, чем другие виды рецепции, и общение между особями осуществляется прежде всего по этому каналу: личинки муравьев выделяют специфические вещества, побуждающие взрослых рабочих особей кормить их, по запаху обитатели одного муравейника узнают «своих» и чужих». Даже о том, жив или мертв муравей, его сородичи узнают по специфическим выделениям.
Согласование поведения обитателей муравейника происходит путем подачи и приема химических сигналов. Достаточно указать на химические средства муравьев (а также пчел и ос), выделение которых служит сигналом тревоги. Эти вещества выделяются муравьями в случае опасности и быстро распространяются по воздуху. Их восприятие вызывает у других муравьев («солдат») защитную реакцию (готовность к нападению на врага), хотя источник опасности им еще не известен. Поскольку, однако, радиус действия такого сигнала, подаваемого одним муравьем (например, в случае вторжения в муравейник какого-либо насекомого), невелик, то он способен указать и место, откуда угрожает опасность. Чем больше эта опасность, тем больше муравьев выделяют химические сигналы тревоги и тем ‘больше, следовательно, особей призывается на помощь.
По другим выделениям, оставляемым на субстрате в виде следа, муравьи узнают, куда, в какую сторону ушел их сородич. К тому же интенсивность такого пахучего следа указывает на количество пищи, обнаруженной в том месте, куда ведет след, тем более что, направляясь к муравейнику, муравьи наносили по пути пахучие метки лишь в тех случаях, когда они находили корм. При этом тем больше оставляется меток, чем больше обнаружено корма. Поскольку эти метки довольно быстро испаряются, вновь идущие по этому следу муравьи будут воспринимать лишь свежие метки, а таких будет все меньше по мере приближения кормового объекта. Таким образом, муравьи в конце концов узнают о том, что пищевые ресурсы в данном месте исчерпаны, и перестанут к нему направляться.
У некоторых пчел также существует химический способ передачи информации о нахождении пищи. Это относится, прежде всего, к пчелам из подсемейства Meliponinae, у которых нет «языка танца». Эти широко распространенные в тропиках пчелы не имеют жала, но яростно кусаются. У мелких форм (самая маленькая имеет длину всего 1,5 миллиметра) нашедшая пищу сборщица только приводит других пчел в возбужденное состояние, беспокойно бегая по сотам, толкая их и время от времени подбегая к летку. Вылетающие же пчелы ориентируются только по запаху цветов. У других видов возбуждение пчел первоначально достигается подобным же образом, но дополнительно сборщица, принесшая взяток, приводит соты толчками в вибрирующее состояние. После этого она возвращается к обнаруженным цветам, а на обратном пути оставляет на траве, ветках или камнях через каждые два-три метра пахучие метки, по которым затем приводит группами других пчел к источнику пищи. Это достигается тем, что пчела разведчица летает взад и вперед между двумя соседними метками, пока другие пчелы не присоединяются к ней. Следовательно,, здесь мы имеем дело с (комплексной передачей информации по оптическому, тактильно-вибрационному и, возможно, акустическому каналам, но при этом ведущую роль играют оставляемые сборщицей запахи.
У нашей медоносной пчелы также существует химический способ передачи информации, но только как вспомогательный, когда она имеет дело с кормом, лишенным собственного заметного запаха. В этих случаях место нахождения кормового объекта метится секретом из так называемой железы Насонова.
Кроме того, как тоже показал Фриш, во второй форме танца угол между линией пробежки и вертикалью (на вертикально расположенных сотах) соответствует углу между линией полета пчелы от улья к кормовому объекту и положением солнца. Таким образом, горизонтальное направление истинного полета переносится на вертикальную плоскость, ориентация по положению солнца подменяется ориентацией по земному притяжению. При этом, если следует лететь навстречу солнцу, пчела танцует снизу вверх, если же от солнца, то сверху вниз (рис. 38, в). Таким образом, медоносная пчела способна к транспозиции с фотоменотаксисов на геоменотаксисы: фиксированный угол к видимому положению солнца переносится в темном улье к силе тяжести.
Эта способность медоносной пчелы к транспозиции, коренящаяся в широко распространенной среди насекомых способности к переносу менотаксисных движений на вертикальную плоскость (рис. 39), обеспечивает, как мы видим, исключительную полноту и точность передачи информации. Такая транспозиция является частным случаем трансформации, лежащей в основе всякого общения между животными: свойства действительных предметов и ситуации трансформируются на языке животных с помощью видоспецифического «кода» в оптические, акустические и прочие условные сигналы, информирующие других особей об этих свойствах. Несмотря на всю сложность и точность, система коммуникации медоносной пчелы в принципе не отличается от любой другой системы общения у животных. Переводятся ли истинные пространственные параметры в сигналы, воспринимаемые с помощью зрения, осязания и чувства силы тяжести, как это имеет место у медоносной пчелы, или же в сигналы, воспринимаемые хеморецепцией (муравьи),- информация в любом случае передается в трансформированном, условном виде. Здесь как нельзя нагляднее выступает характер и функциональное значение психики у животных.
Вершиной развития инстинктивных компонентов общения является ритуализация поведения, которая особенно четко выступает в воспроизводительном поведении насекомых, в частности в «ухаживании» самцов за самками. Вот типичный брачный «ритуал» самца бабочки бархатницы, описанный Тинбергеном: полетев вслед за самкой, самец садится рядом с ней, потом становится перед ней, начинает «дрожать» крыльями (передне-задние движения сомкнутых крыльев), затем повторно раскрывает и закрывает их, что сопровождается постукиванием усиками по крыльям самки. После этого самец обхватывает самку крыльями и зажимает между ними ее усики, которые при этом прикасаются к пахучему участку на крыльях самца. Только после этого, если самка не отвергает «ухаживания» самца, начинается спаривание.
В этом примере обращает на себя внимание строгая фиксированность последовательности сигнальных поз и телодвижений, что и характерно для ритуализованного поведения. Как мы видим, при ритуализации двигательные акты всегда носят условный характер: выполняемые животным движения несут лишь сигнальную, но не непосредственно «механическую» функцию. Именно на этой основе зародился язык поз, телодвижений и звуков у животных во всем своем удивительном многообразии. При этом в условности сигналов, находит непосредственное воплощение психический компонент общения.
Наиболее сложные системы общения присущи, конечно, насекомым, живущим большими семьями. Это относится опять-таки в первую очередь к медоносной пчеле и муравьям. Дифференцированные и многообразные взаимоотношения между членами пчелиной семьи, опирающиеся на сложные формы коммуникации, являются уникальными в животном мире. По поводу этого сообщества высказывались самые различные мнения начиная от антропоморфичного толкования ее как копии человеческого общества или государства (сохранилось неудачное, вводящее в заблуждение название «общественные насекомые») до представления о некоей сумме независимо действующих «рефлекторных автоматов». Вместе с тем оказалось (исследования К. П. Трубецкой), что пчелиная семья представляет собой объединение особей, поведение которых носит самостоятельный, но не независимый характер. Самостоятельность поведения отдельных пчел проявляется прежде всего в резко выраженных индивидуальных различиях: предпочтение отдельных видов ульевых работ, различия в сроках и характере перехода от ульевой деятельности к летной и др. Большую сложность в количественном и качественном аспектах достигают взаимоотношения между обитателями улья, в результате чего функциональная настроенность одних пчел может оказать влияние на других. Рабочие особи с их повышенным уровнем нервной деятельности часто сообщают возбуждение другим, менее активным рабочим пчелам.
Весьма существенным признаком территориального поведения является активная защита участка хозяином. Тинберген придает этому критерию наряду с привязанностью к местности особое значение. Враждебное отношение («агрессивность») проявляется хозяином по отношению к любому представителю того же вида, особенно того же пола.
Часто такое поведение приурочено лишь к периоду размножения. Так, например, древесный таракан (Cryptocereus punctulatus), устраивающий гнездовую камеру в гнилой древесине, весьма интенсивно (и обычно успешно) защищает место, где находится эта камера, от вторжения других самцов. В случае поражения гнездовую камеру, где находится и самка, занимает победитель.
В высокоразвитых формах территориальное поведение, как показал немецкий этолог А. Хеймер, встречается у стрекоз. Это интересно уже потому, что стрекозы относятся к самым древним насекомым и, появившись в палеозое, дожили до наших дней, не претерпев за эти 250 миллионов лет существенных морфологических изменений. Вместе с тем, как показывает Хеймер, архаичность строения сочетается у них с высокоразвитым поведением, причем не только в сфере территориальности. Этот, казалось бы, парадоксальный факт сохранения древнейших признаков строения при наличии весьма прогрессивных форм поведения вновь подтверждает общее правило несоответствия морфологических и поведенческих признаков, если иметь в виду морфофункциональные отношения на уровне целого организма.
Таким образом, у данных видов стрекоз существует вполне развитое территориальное поведение: занимаются индивидуальные участки, в которых выделяются функциональные зоны (отдыха, размножения), на этих участках выполняются все жизненные функции (за исключением сна: вечером самцы собираются в особых местах ночевок), хозяин дает знать о своем присутствии, маркирует и активно защищает участок.
Интересные особенности территориального поведения муравьев, были выявлены советским энтомологом А. А. Захаровым. Оказалось, что у муравьев существуют два основных типа использования кормовых участков: совместное использование угодий несколькими семьями и использование кормового участка населением лишь, одного гнезда. При этом выявилась прямая связь между плотностью муравьев на участке и «агрессивностью»: у видов с низкой плотностью участки не охраняются (кроме пригнездовой зоны), при высокой же плотности на кормовых участках появляются охраняемые территории, а между ними «нейтральные зоны» (рис. 40). Как показал другой советский исследователь, Г. М. Длусский, муравьи того же вида, принадлежащие к другим семьям, равно как представители других видов, в пределы этих территорий не допускаются.
Захаров справедливо отмечает сходство территориального поведения муравьев с таковым у птиц и хищных млекопитающих.
Инстинкт и научение в поведении насекомых
Долгие годы господствовало мнение, будто насекомые и другие членистоногие являются существами, поведением которых руководит жесткий «слепой инстинкт». Это представление укоренилось преимущественно под влиянием работ выдающегося французского энтомолога Ж. А. Фабра, который сумел своими блестящими исследованиями убедительно показать, что даже сложнейшие действия насекомых не являются проявлением «разума», а выполняются на врожденной, инстинктивной основе. Одностороннее развитие положений Фабра и привело к указанной неверной оценке поведения насекомых, к отрицанию не только разумности их поведения, но и к отрицанию, или хотя бы умалению, роли накопления индивидуального опыта, научения в их жизни.
Как мы уже видели, формирование любой формы видотипичного, наследственно «закодированного», т. е. инстинктивного, поведения в онтогенезе всегда сопряжено в той или иной степени с какими-либо элементами индивидуально приобретаемого поведения, научения. О строго фиксированном инстинктивном поведении в «чистом виде» не приходится говорить даже относительно низших животных.
В полной мере это относится и к насекомым, инстинктивное поведение которых также совершенствуется научением. В этом состоит основная роль научения в жизни насекомых. Можно, очевидно, считать, что научение стоит у насекомых и других членистоногих «на службе» у инстинктивного поведения. Как и у других животных, инстинктивные движения (врожденные двигательные координации) у них генетически строго фиксированы. Инстинктивные же действия, инстинктивное поведение являются и у насекомых в той или иной степени пластичными благодаря включению в них благоприобретаемых компонентов.
Более того, как установили названные исследователи, биологическая значимость сигнальной деятельности медоносных пчел определяется стереотипом условных рефлексов, приобретаемых в онтогенезе по мере освоения пространства и при общении в семье. Оказалось, что интерпретация передаваемой в танце информации о расстоянии и направлении полета к источнику пищи возможна лишь в том случае, если пчела до этого научилась соотносить местонахождение корма с характером информации, содержащейся в танце сборщиц. Кроме того, тактильный компонент танца (вибрации брюшка) не имеет врожденного сигнального значения. Последнее приобретается в онтогенезе также условно рефлекторным путем: пчелы, не имевшие в онтогенезе контактов (пищевых) с танцовщицей, не в состоянии интерпретировать этот существеннейший элемент танца. Следовательно, каждая пчела должна в основном научиться «понимать» язык танца. С другой стороны, образование временных связей оказалось важным и для формирования самой способности к выполнению танцев.
Таким образом, нет неизменных форм поведения даже там, где прежде всего требуется стереотипность,- в сигнальных позах и телодвижениях. Даже такое врожденное коммуникативное поведение, как «танцы» пчел, не только дополняется и обогащается процессами научения, не только переплетается с ними, но и формируется в комплексе с индивидуально приобретаемыми элементами поведения.
Конечно, медоносная пчела занимает среди насекомых исключительное положение, и далеко не у всех представителей этого огромного класса психическое развитие достигает такой высоты. Об исключительных психических качествах медоносной пчелы свидетельствуют, в частности, экспериментальные данные, говорящие о наличии у нее аналогов некоторых высших психических функций позвоночных. Речь идет об установленной Мазохиным-Поршняковым высокоразвитой способности пчелы к зрительным обобщениям, например, типа «треугольник» и «четырехугольник» (независимо от конкретной формы, соотношения размеров и взаимной ориентации фигур) (рис. 41, а), «двуцветность» и др. В одной из серий опытов пчелам предлагалось выбрать из попарно предъявляемых фигур те, у которых один локальный признак (зачерченный кружочек) находился на конце цепочки из кружочков независимо от длины и формы этих цепей (рис. 41, б). Со всеми предложенными им задачами, даже в наиболее сложных вариантах, пчелы вполне справлялись. При этом отмечалась большая пластичность, нестандартность поведения, что экспериментатор справедливо связывает с непрерывной изменчивостью условий среды (непостоянство освещения, взаимного расположения, формы, окраски и многих других признаков компонентов среды), при которых этим насекомым приходится добывать пищу. Мазохин-Поршняков приходит к выводу, что выбор незнакомого объекта на основе обобщенных зрительных образов (иногда неправильно обозначаемых им как «понятия») есть свидетельство нестандартного использования пчелами личного опыта, его применения в новой ситуации, отличной от обстановки первоначальной выработки соответствующего навыка.
Таким образом, здесь справедливо подчеркивается наличие и значение факта переноса определенного навыка в новую ситуацию и решение сложной задачи на основе индивидуального опыта, зафиксированного в виде обобщенного зрительного представления. В этом отношении мы действительно находим уже у пчел психические способности, аналогичные тем, которые относятся к предпосылкам интеллектуальных действий высших позвоночных животных. Однако одних этих предпосылок недостаточно для интеллектуального поведения, мышления животных, особенно если смотреть на эти высшие психические функции животных как на ступень по направлению к зарождению человеческого сознания. Поэтому описанные способности пчел не могут служить и критерием для признания у них мышления и уж во всяком случае не приходится говорить о наличии у пчелы рассудочной деятельности, хотя бы и в элементарной форме, как толкует результаты своих исследований Мазохин-Поршняков. Признавая у высших животных наличие своеобразных мыслительных способностей, интеллекта, нужно со всей определенностью отдавать себе отчет в том, что рассудок, т. е. разум, сознание как качественно иная категория психического отражения не присущи ни одному из животных, а только человеку.
Общая характеристика низшего уровня перцептивной психики
На низшем уровне перцептивной психики уже представлены все те прогрессивные признаки, которые характеризуют перцептивную психи-ку вообще, но во многих отношениях поведение относящихся сюда животных носит и примитивные черты, сближающие его с поведением нижестоящих животных. Так, основную роль играет ориентация поведения по-прежнему по отдельным свойствам предметов, но не по предметам как таковым: предметное восприятие явно играет еще подчиненную роль в общем поведении. Равным образом в последнем преобладают ригидные, «жестко запрограммированные» элементы поведения над гибкими, благоприобретаемыми и т. д.
С другой стороны, мы констатируем на этом уровне четко выраженный активный поиск положительных раздражителей, т. е. положительное таксисное поведение получает мощное развитие. Налицо все виды высших таксисов, включая мнемотаксисы. Последние играют в пространственной ориентации особенно существенную роль, и именно в индивидуальном заучивании ориентиров проявляется в наибольшей степени способность к благоприобретаемому изменению видового поведения, к научению.
Вместе с тем важно подчеркнуть, что хотя у рассматриваемых здесь животных, в частности насекомых, накопление индивидуального опыта, научение играют существенную роль, наблюдается и определенная противоречивость в процессах научения, сочетание прогрессивных и примитивных черт. Специфическая направленность, приуроченность этих процессов к определенным функциональным сферам, как и само подчиненное положение, которое занимает научение по отношению к инстинктивному поведению, несомненно, указывают на переходное положение данного уровня психического развития между элементарной сенсорной и развитой перцептивной психикой.
Но, как мы видели, это никоим образом не означает, что насекомым, как и другим представителям рассматриваемой группы животных, недостает пластичности поведения. Наоборот, и здесь в полной мере проявляется общая закономерность, что усложнение инстинктивного поведения неизбежно сочетается с усложнением процессов научения (и наоборот). Только такое сочетание обеспечивает подлинный прогресс психической деятельности.
Инстинктивное поведение представлено на рассматриваемом уровне психического развития уже весьма развитыми новыми категориями: групповое поведение, общение, ритуализация. Особую сложность приобретают формы общения у видов, живущих огромными семьями, из которых лучше всего изучены пчелы. Язык пчел, этих высших представителей членистоногих, относится к наиболее сложным формам общения, которые вообще существуют в животном мире. Возможно, что новые исследования познакомят нас с поразительными психическими способностями и других насекомых, но на сегодняшний день пчелы представляются нам наиболее развитыми в этом отношении. Наиболее сложные формы инстинктивного поведения закономерно сочетаются у них с наиболее разнообразными и сложными проявлениями научения, что обеспечивает не только исключительную согласованность действий всех членов пчелиной семьи, но и максимальную пластичность поведения особи. Психические способности пчел (как и некоторых других высших насекомых) в некоторых отношениях, очевидно, уже выходят за рамки низшего уровня перцептивной психики.
В ином направлении, чем у членистоногих, шло развитие психической активности у головоногих моллюсков. По некоторым признакам они приблизились к ветви, ведущей к позвоночным, о чем уже свидетельствуют их крупные размеры и отмеченные выше особенности строения нервной системы и особенно зрительного рецептора, что непосредственно связано с резким увеличением скорости движения по сравнению с другими моллюсками.
Поведение головоногих еще совершенно недостаточно изучено, но уже известны многие примечательные их способности. Прежде всего они выделяются существенным усложнением инстинктивного поведения. У головоногих уже встречаются территориальное поведение (занятие и защита индивидуальных участков), «агрессивность», которая только намечается у высших червей, групповое поведение (стайная жизнь кальмаров и каракатиц), в сфере размножения появляются ритуализованные формы поведения, что находит свое воплощение в видоспецифическом «ухаживании» самцов за самками. Все это присуще только высшим животным, помимо головоногих, членистоногим и позвоночным.
Рядом исследователей обращалось особое внимание на весьма развитое у осьминогов «любопытство», что выражается в обследовании ими биологически «бесполезных» предметов, а также на их высокоразвитые манипуляционные и конструктивные способности. Эти способности проявляются в строительстве валов и убежищ из камней, панцирей крабов, раковин устриц и т. п. Этот строительный материал осьминог подбирает, переносит и укрепляет «руками». Иногда такие гнезда представляют собой закрытые со всех сторон строения. По некоторым, правда, еще не подтвержденным наблюдениям осьминоги способны и к орудийным действиям, пользуясь камнями для защиты.
Очень важным представляется и то обстоятельство, что впервые у головоногих появляется способность к установлению контактов с человеком, к общению с ним, результатом чего является возможность подлинного приручения этих животных (в отличие от насекомых!).
Таким образом, головоногие, далеко оставив позади других моллюсков, как и вообще всех низших беспозвоночных, достигли, несомненно, высокого уровня психического развития, во многом сблизившись с позвоночными животными.
Вместе с тем и у головоногих наблюдается та же противоречивость в способности к научению, что и у насекомых. Так, например, у осьминога вообще хорошо развита способность к научению на зрительные и тактильные стимулы, но в ряде случаев он оказывается не в состоянии решить, казалось бы, несложные задачи. Особенно это относится к преодолению преград: осьминог неспособен находить обходной путь, если приманка (краб) располагается за прозрачной преградой (в стеклянном цилиндре или за проволочной сеткой). Тщетно пытаясь овладеть приманкой в прямом направлении, осьминог не хватает ее сверху, через край. Правда, по Бойтендийку, некоторые осьминоги все же способны решить несложные задачи обходного пути. При этом, очевидно, большое значение имеет прежний опыт особи. Другие головоногие уступают осьминогу по своим психическим способностям.
На низшем уровне перцептивной психики находится, очевидно, хотя бы отчасти, и ряд представителей низших позвоночных. Однако в корне различные строение и образ жизни членистоногих и позвоночных являются причиной того, что и их поведение и психика, в сущности, несопоставимы. Так, одной из отличительных особенностей насекомых являются их малые по сравнению с позвоночными размеры. В связи с этим окружающий насекомое мир представляет собой нечто совершенно особое: это не микромир простейших, но и не макромир позвоночных. Человеку трудно себе представить этот мир насекомых с его (с нашей точки зрения) микроландшафтами, микроклиматами и т. д. Хотя насекомые живут рядом и вместе с нами, они живут в совершенно иных условиях температуры, освещения и т. п. Уже поэтому психическое отражение действительности не может у насекомых не быть принципиально иным, чем у позвоночных, да и у большинства других беспозвоночных.
Но поскольку наиболее общие признаки психического отражения, характерные для данного его уровня, присущи всем упомянутым животным, можно, очевидно, по поводу рассмотренных нами в качестве примера насекомых сказать, что мы имеем здесь дело с типичными проявлениями низшего уровня перцептивной психики, но в формах, отвечающих тем особым условиям жизни этих животных, которые отмечались выше.



