Что такое пищеварительная вакуоль

«Вакуом» – это старинный термин, который был возрожден Де Дювом. В это понятие он включил все участки клетки, окруженные мембраной, кроме митохондрий. Составной часть вакуома является лизосомальная система в состав которой входит вакуолярная система, представляющая собой совокупность одномембранных органелл клетки. Вакуолярная система играет важную роль в жизнедеятельности клеток всех царств живой природы. Она связана с процессами распада и транспорта органических веществ и представляет собой динамичную систему, состоящую из канальцев, цистерн и пузырьков, которые сообщаются между собой путем установления постоянных или временных связей в результате слияния их мембран. Благодаря наличию вакуома вещества способны поступать в клетку, перемещаться внутри нее, выводиться наружу и, не смешиваясь с ее материалом, обмениваться продуктами путем диффузии или активного транспорта. Кроме того. вакуом, как показывают исследования, играет значительную роль в формировании приобретенной локальной устойчивости растительной клетки, инфицированной вирусом.
Научная проблема и ее обоснование
Роль вакуома в функционировании растительной клетки изучена до настоящего времени недостаточно и продолжает интенсивно исследоваться. Известно, что внутри вакуолярной системы, являющейся составной частью вакуома, выделяется два основных метаболических потока – анаболический и катаболический. Анаболический связан с синтезом, транспортом, секрецией белков и с формированием первичных лизосом. Катаболический связан с процессами лизосомального переваривания.
При этом поток вещества может проходить через весь вакуом, начинаясь от эндоплазматического ретикулума, где они синтезируются, до внеклеточного пространства. В противоположном направлении движения соединений имеется блокирующий механизм, который препятствует попаданию веществ экстрацеллюлярного пространства во внутреннюю часть системы. По некоторым данным это обусловлено невозможностью слияния цитомембран эндоплазматического и экзоплазматического типа из-за ключевых отличий в их строении и химическом составе. Например, мембраны, принадлежащие митохондриям, ШЭР и аппарату Гольджи, являются более тонкими, по сравнению с везикулами Гольджи и некоторыми видами лизосомальных мембран. Основной органеллой, осуществляющей дифференциацию и трансформацию мембран в клетках, как растений, так и животных организмов, считается аппарат Гольджи. Было выяснено, что постепенное увеличение толщины клеточных мембран происходит в аппарате Гольджи в направлении от его проксимального полюса (наружная сторона) к дистальному (внутренняя сторона). В связи с этим Де Дювом было предложено подразделить вакуом на две части:
– эндоплазматическое пространство, которое приблизительно соответствует пространству, занятому эндоплазматическим ретикулумом (ЭР) и ядерной оболочкой;
– эктоплазматическое пространство, в которое входят все остальные окруженные мембраной клеточные компоненты.
Функции эндоплазматического пространства, по мнению Де Дюва, не ограничиваются чисто катаболическими процессами. Эта система является также местом хранения эндогенных биосинтетических продуктов, запасаемых в различных секреторных гранулах и первичных лизосомах, а также экзогенных веществ, поступающих в клетку в процессе эндоцитоза. Другая функция системы связана с экзоцитозом (процессом, обратным эндоцитозу), при котором из клетки выводятся секреторные продукты в результате слияния секреторных гранул с плазмаллемой, а также трансклеточный транспорт, или диацитоз, при котором эти вещества, проходя через определенный слой клеток, попадают во внеклеточную среду. В эндоплазматическом пространстве происходит также взаимодействие между веществами, изолированными внутри различных везикул в случае их слияния. Эта функция включает в себя не только переваривание эндоцитозного материала лизосомальными ферментами, но и переваривание вещества секреторных гранул при их слиянии с фагоцитозными вакуолями и с лизосомами. Впервые взаимодействие лизосом с секреторными гранулами было обнаружено Смитом и Даркуа. Этот процесс существенно отличается от обычной клеточной автофагии тем, что вместо предварительной изоляции предназначенных для переваривания структур мембранными элементами или инкорпорации (внедрения) их в первоначально интактном виде внутрь лизосом здесь имеет место истинное слияние мембран.
Автофагией называется способность клеток переваривать собственные структурные компоненты или участки цитоплазмы, окруженные изолирующими их клеточными мембранами, с образованием впоследствии автофаговой вакуоли. Автофагия, как показывают исследования, наблюдается в клетках всех эукариот. Она играет важную роль в поддержании внутриклеточного гомеостаза, а также в осуществлении процессов нормального онтогенетического развития (рост, дифференцировка, метаморфоз). В клетках, подвергающихся стрессу, автофагия является наиболее общей субклеточной реакцией.
Единого мнения на происхождение изолирующих клеточных мембран в настоящее время нет. Существует мнение, что их происхождение связано с эндоплазматическим ретикулумом (ЭР). Две мембраны, ранее принадлежащие ЭР и разделенные просветом, после завершения изоляции участка цитоплазмы сливаются в одну более толстую, образуя таким образом цитосегресому. Согласно другой точке зрения, внутренняя мембрана ЭР растворяется, и содержимое образовавшейся цитосегресомы становится доступным для гидролитических ферментов, локализованных ранее в изолирующей цистерне. Механизм описанных процессов в настоящее время до конца не выяснен. Кроме ЭР источником изолирующих мембран может быть и аппарат Гольджи. Предполагается также возможность формирования цитосегресом в результате слияния вакуолей. Изолирующая мембрана в этом случае происходит от плазмалеммы. Автофаговые вакуоли могут быть образованы и в результате слияния мембран цитосегресом с мембранами лизосом. Имеются данные о существовании регуляторных механизмов, управляющих процессами автофагии и способных изменить скорость и частоту этих процессов в различных ситуациях (суточный ритм, стрессовые воздействия, дифференцировка). Мощными индукторами автофагии могут быть некоторые алкалоиды, антибиотики.
В клетках растений и животных имеется еще один механизм деструкции – кринофагия. Она заключается в прямом слиянии первичных лизосом с мембранной оболочкой органелл, что ведет к лизису последней. Первичные лизосомы – это клеточные структуры, содержащие в своем матриксе набор гидролитических ферментов и еще не успевшие принять участие в процессах переваривания.
Автофагия и кренофагия – это один из механизмов образования вакуолей в растительных и животных клетках. Происхождение и статус клеточных вакуолей в настоящее время является дискуссионным. Особенно большой интерес вызывает большая центральная вакуоль растительных клеток. Вопрос о ее происхождении возник еще в ХIХ веке, но до сих пор не имеет однозначного решения. До сих пор ведется дискуссия по поводу того, является ли ее содержимое просто жидкой фазой, окруженной мембраной, или она является самостоятельной органеллой, способной к активному росту и развитию. По мнению некоторых авторов, возникновение центральной вакуоли можно связать со слиянием уже существующих более мелких вакуолей или формированием ее из каких-либо иных вакуолярных элементов, либо с образованием из структур, не имеющих непосредственного отношения к вакуолярной системе. Наиболее распространено мнение о формировании предшественников вакуолей из локальных участков канальцев ЭР, которые постепенно расширяются и затем трансформируются в тонопласт. Ряд исследователей придерживаются мнения, что вакуоли образуются из элементов аппарата Гольджи. Существует также представление о возможности появления в цитоплазме растительных клеток вакуолей эндоцитозного происхождения, окружающая мембрана которых первоначально принадлежала плазмалемме.
Исследования показали, что вакуоли в живых клетках содержат довольно широкий набор ферментов – это кислая фосфатаза, пероксидаза, цитохро-С-редуктаза, вещества фенольной природы, запасные белки, а также различные низко- и высокомолекулярные метаболиты и токсичные соединения. Большинство вакуолярных ферментов связано с вакуолярной мембраной, хотя такие кислые гидролазы, как фосфатаза и карбоксипептитаза, найдены в клеточном соке. Изучение распределения глобул белков в тонопласте позволяет отнести его к числу функционально активных мембран.
Вакуоли одной и той же клетки могут отличаться друг от друга по своему химическому содержимому. Это, видимо, с одной, стороны свидетельствует о различном их происхождении, а с другой, о возможном отличии в выполняемой ими функции. Низкомолекулярные соединения могут поступать из вакуолей обратно в цитоплазму, что, по-видимому, объясняет уменьшение объема этих структур на определенных стадиях клеточного цикла. Эти же свойства вакуолей придают им характер буфера между наружной и внутренней средой, контролирующего гомеостаз растительной клетки и ответственного за изменение вязкости цитоплазмы. Клеточные вакуоли выполняют также ряд других важных функций. Наиболее известной из них является осморегуляция и поддержание тургора путем создания в клетках высокого гидростатического давления. В настоящее время наибольший интерес вызывает лизосомальная функция растительных вакуолей, что согласуется с представлением некоторых исследователей о происхождении вакуолей из производных диктиосом аппарата Гольджи.
Впервые растительные лизосомы, как структуры, сходные с лизосомами животных, были обнаружены при цитохимическом изучении меристематических клеток. Лизосомы принято подразделять на первичные и вторичные. Первичные лизосомы – это микротельца, ферменты которых еще не принимали участие в процессе переваривания. Микротельца, являющиеся местом активного литического процесса, называются вторичными лизосомами. Среди вторичных лизосом, в зависимости от того, какой материал подвергается лизису, выделяют гетеролизосомы (переваривают внеклеточные вещества) и автолизосомы (переваривают вещества клетки). На поздних стадиях те и другие получили название телолизосомы (остаточные тельца). Деградирующие телолизосомы, утратившие свою активность, названы постлизосомы. В животных клетках обнаружены образования, названные прелизосомами. Они не имеют собственной гидролитической активности и предназначены для последующего переваривания лизосомами. В зависимости от их происхождения выделяют гетерофагосомы и автофагосомы (цитосегресомы). Французскими исследователями была предложена классификация, позволяющая с помощью ультраструктурных и цитохимических критериев идентифицировать среди различных производных диктиосом органеллы лизосомальной природы. Согласно этой классификации существует 3 вида структур:
– электроннопрозрачные относительно крупные пузырьки, содержащие полисахариды;
– визикулы более мелких размеров электронноплотные, содержащие кислые гидролазы;
– пузырьки, морфологически сходные с первыми, но предположительно выполняющие функцию транспорта полисахаридов от аппарата Гольджи к формирующейся либо растущей клеточной оболочке. В некоторых случаях они участвуют в образовании специализированных вакуолей, богатых полисахаридами.
Функцию транспорта полисахаридов приписывают также и первым структурам, вторые предположительно представляют собой протофитолизосомы, созревающие впоследствии в фитолизосомы.
Существует мнение, что в клетках, как растений, так и животных, помимо формирования первичных лизосом посредством аппарата Гольджи существует и другой значительно более упрощенный путь отпочковывания их непосредственно от канальцев и цистерн ЭР. Предполагают, что этот путь связан со стимуляцией лизосомального аппарата при различных патологических состояниях как естественной природы, так и вызванных экспериментально.
Исходя из исследований (А.В. Реунов 1999.; Нагорская 2000; С.Н. Лега 2002; и др.) можно выделить, как нам представляется, наряду с другими многочисленными функциями, также и защитную функцию вакуолярной системы, которая заключается в способности вакуолей, в результате активации лизосомального компартмента при вирусном патогенезе, участвовать в разрушении вирусных частиц. Подтверждением активации литических процессов в клетках растений при вирусном патогенезе является обнаружение в принекрозных зонах (1–2 см от края ВТМ-индуцированного локально некроза) листьев Datura stramonium L. (С.Н. Лега 2002) процессов автофагии. Об этом свидетельствует наличие электроннопрозрачных участков цитоплазмы, изолированных замкнутыми мембранами ретикулума, которые нередко содержали включения электронноплотного и, вероятно, лизируемого материала. Кроме того, в принекрозных зонах было зафиксированно повышеное содержание, по сравнению со здоровыми клетками, пузырьков Гольджи, набухание элементов ЭПР и превращение их в вакуоли. Свидетельством развития процессов автолиза в клетках листьев растений, инфицированных вирусом, является формирование в их цитоплазме также цитосегресом, цитосом, и образований, в которых нередко обнаруживались подвергшиеся лизису мембранные структуры.
Стимуляция процессов автолиза имеет место и в зараженных ВТМ клетках листьев растений Gomphrena qlobosa L. расположенных вокруг ВТМ-индуцированных хлоротичных зон (С.Н. Лега, 2002). При инфицировании листьев гомфрены ВТМ на них образуются хлоротичные поражения, напоминающие локальные некрозы (что, однако, не сопровождается локализацией инфекции), в которых выявлялись значительные скопления вирусных частиц. За пределами хлоротичных зон вирусные частицы обнаруживались в значительно меньшем количестве и имели, как показали исследования, необычно высокую осмиофильность и способность к слипанию. Иногда вирусные частицы утрачивали присущую им упорядоченность и имели хлопьеобразный вид, что свидетельствует о деградации вируса. Кроме того, в этих участках листа наблюдались существенные морфологические изменения клеточных структур, о чем говорит повышенная осмиофильность тонопласта, посветление матрикса митохондрий и нарушение целостности их крист, наличие электроннопрозрачных зон в участках цитоплазмы на границе с центральной вакуолью, содержащих деградируемые вирусные частицы (С.Н. Лега, 2002).
Активация литического компартмента, о чем свидетельствует активация процесса автофагии, увеличение количества и активности аппарата Гольджи а также деградация вирусных частиц в клетках листьев растений, зараженных вирусом, по-видимому, связана с механизмом формирования в них приобретенной локальной устойчивости.
Выводы и перспективы
Таким образом, вакуом, а именно лизосомальная и в частности вакуолярная система, как показывают исследования, играет весьма важную роль в реализации защитных механизмов при формировании приобретенной локальной устойчивости в инфицированной вирусом растительной клетке.
Рецензенты:
Галкин М.А., д.б.н., профессор, заведующий кафедрой ботаники, Пятигорский медико-фармацевтический институт, филиал, Волгоградский государственный медицинский университет, г. Пятигорск;
Коновалов Д.А., д.фарм.н., профессор, заведующий кафедрой, заместитель директора ПМФИ по научной работе, Пятигорский медико-фармацевтический институт, филиал Волгоградский государственный медицинский университет, г. Пятигорск.
Что такое пищеварительная вакуоль
Принято различать несколько типов и подтипов пищеварения.
Гидролиз пищевых веществ за счет ферментов, вырабатываемых пищеварительными железами самого организма, характерен для собственного типа пищеварения. В результате осуществления собственного типа пищеварения образуется основное количество олигомеров, поступающих в кровь и лимфу. Расщепление компонентов пищи ферментами, синтезируемыми микроорганизмами, которые обитают в пищеварительном тракте, называют симбионтным типом пищеварения (поскольку он является следствием симбиоза организмов хозяина и микробов). Так переваривается клетчатка в толстом кишечнике человека.
Гидролиз пищевых веществ ферментами, поступающими в пищеварительный тракт вместе с пищей, относят к аутолитическому типу пищеварения, так как происходит самопереваривание. Аутолитическое пищеварение играет важную роль у новорожденного, потому что компоненты грудного молока перевариваются ферментами, входящими в его состав.
 Рис. 11.1. Схема внеклеточного, внутриклеточного и мембранного гидролиза пищевых веществ.
Рис. 11.1. Схема внеклеточного, внутриклеточного и мембранного гидролиза пищевых веществ.
1 — внеклеточная среда; 2 — перевариваемый субстрат и продукты его гидролиза; 3 — ферменты; 4 — внутриклеточная среда; 5 — мембрана энтероцита; 6 — ядро; 7 — внутриклеточная пищеварительная вакуоль, 8 — мезосома.
А — внеклеточное (дистантное) пищеварение. Полимеры и олигомеры пищевых веществ под влиянием ферментов пищеварительных соков в полости кишки гидролизуются до мономеров, которые через мембрану энтероцита транспортируются в его цитоплазму. Б — внутриклеточное цитоплазматическое пищеварение. Олигомеры пищевых веществ проникают через мембрану энтероцита в его цитоплазму и под влиянием ферментов, находящихся в цитоплазме, превращаются в мономеры. В — внутриклеточное вакуольное (внеплазматическое) пищеварение, связанное с эндоцитозом. В мембране энтероцита образуется выпячивание, которое заполняется перевариваемым субстратом и превращается в вакуоль. Вакуоль соединяется с мезо-сомой, заполненной ферментами, которые расщепляют субстрат до конечных продуктов гидролиза, поступающих через мембрану вакуоли в цитоплазму энтероцита. Г — мембранное пищеварение. Адсорбированные на внешней поверхности мембраны энтероцита ферменты расщепляют олигомеры пищевых веществ до мономеров, которые затем поступают в цитоплазму клетки.
В зависимости от локализации процесса гидролиза пищевых веществ различают два типа пищеварения — внутриклеточное и внеклеточное.
Внутриклеточное пищеварение — расщепление мельчайших частичек пищевых веществ, поступивших в энтероцит путем эндоцитоза, за счет клеточных ферментов. Этот тип пищеварения играет важную роль в кишечном пищеварении в раннем постнатальном периоде развития. По мере формирования функций пищеварительного тракта у ребенка значение внутриклеточного пищеварения уменьшается.
Внеклеточное пищеварение А. М. Уголев предложил делить на 2 подтипа — дистанционное и пристеночное.
Дистанционное (полостное) пищеварение осуществляется в полостях пищеварительного тракта, удаленных от мест выработки ферментов. В процессе полостного пищеварения деполимеризация молекул пищевых веществ совершается в основном до олигомеров. Пристеночное пищеварение (контактное, мембранное) совершается в тонком кишечнике — в пристеночном слое слизи, на поверхности ворсинок и микроворсинок, в гликокаликсе (мукополисахаридных нитях, связанных с мембраной микроворсинок). В слизи и гликокаликсе содержится много адсорбированных ферментов пищеварительных соков, выделенных в полость кишки и расположенных на огромной площади соприкосновения с перевариваемым субстратом. Поэтому в процессе пристеночного пищеварения значительно увеличивается скорость гидролиза пищевых веществ, что приводит к возрастанию объема всасывания продуктов гидролиза.
Схема внеклеточного и мембранного пищеварения представлена на рис. 11.1. Из этой схемы следует, что при внеклеточном пищеварении (А) ферменты расщепляют субстрат в полости пищеварительного тракта до конечных продуктов гидролиза, которые затем проникают в цитоплазму энтероцита.
В процессе внутриклеточного цитоплазматического пищеварения (Б) крупные осколки молекул пищевых веществ проникают через мембрану энтероцита в его цитоплазму и расщепляются ее ферментами до мономеров. При внутриклеточном вакуольном пищеварении (В) мельчайшие частички субстрата захватываются из полости кишки мембраной энтероцита, образующей выпячивание, которое превращается в вакуоль. Вакуоль, содержащая субстрат, объединяется с вакуолью (мезосомой), наполненной ферментом, который и осуществляет гидролиз субстрата. Мембранное пищеварение (Г) характеризуется тем, что интенсивный процесс гидролиза крупных осколков молекулы питательных веществ до мономеров происходит на поверхности энтероцита за счет ферментов, адсорбированных на его мембране.
 Рис. 11.2. Схема деполимеризации частичек пищевых веществ на поверхности микроворсинок энтероцита, апикальном и латеральном гликокаликсе.
Рис. 11.2. Схема деполимеризации частичек пищевых веществ на поверхности микроворсинок энтероцита, апикальном и латеральном гликокаликсе.
1—3 — перевариваемые субстраты в полости кишки; 4 — апикальный гликока-ликс; 5 — латеральный гликокаликс; 6 — мембрана микроворсинки; 7 — микроворсинки энтероцита.
Крупные осколки пищевых веществ в полости кишки под влиянием ферментов пищеварительных соков расщепляются до олигометров (1—3). Их гидролиз на поверхности микроворсинок и нитей гликокаликса (4—6) завершается образованием монометров, молекулы которых проникают в микроворсинку, а затем — в цитоплазму энтероцита (8).
На рис. 11.2 схематически показано, что на поверхности микроворсинок энтероцитов и нитей апикального и латерального гликокаликса происходит гидролиз частичек пищевых веществ за счет адсорбированных ферментов. Поскольку суммарная площадь, на которой происходит гидролиз пищевых веществ, очень большая, то это обусловливает высокую эффективность мембранного пищеварения
Заключительные стадии гидролиза пищевых веществ осуществляют ферменты, синтезируемые энтероцитами и встроенные в структуры их мембран. Образующиеся на поверхности мембраны энтероцита мономеры всасываются за счет активности ее ионных каналов.
Научная электронная библиотека

Глущенко Л. Ф., Глущенко Н. А.,
1.1.1. Строение и химический состав дрожжевой клетки
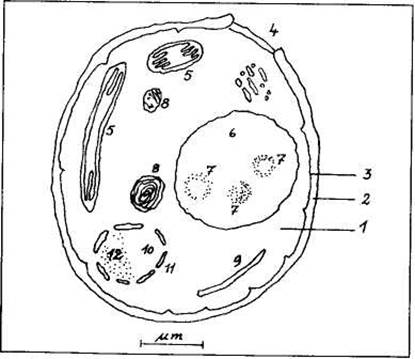
Дрожжевая клетка содержит большое количество митохондрий (5). Митохондрии получают пируват, образующийся в цитоплазме, и разлагают его в процессе дыхания на диоксид углерода и воду с образованием аденозинтрифосфата (АТФ) и аденозиндифосфата (АДФ), представляющих собой носители энергии. Поэтому митохондрии называют иногда «энергетическими станциями клетки».
Шероховатая эндоплазматическая сеть (ЭС) (9) служит для синтеза протеина, а гладкая эндоплазматическая сеть синтезирует липиды и отвечает за процессы освобождения от ядовитых веществ. Образующийся протеин блокируется и перемещается в предусмотренное место в везикулах, снабженных оболочкой. Эту задачу берет на себя комплекс Гольджи, представляющий собой своего рода «сортировочную станцию». Секреторная везикула с ядовитым веществом (например, со спиртом) транспортируется к клеточной мембране и выносится наружу.
За переработку отходов клетки отвечают лизосомы, обеспечивающие внутриклеточное пищеварение и разлагающие высокомолекулярные структуры в низкомолекулярные. Рибосомы синтезируют протеин и распределяют его в клетке. Тем самым они отвечают за соединение аминокислот с образованием продуктов генного синтеза в соответствии с информацией, получаемой из ядра клетки.
Энергоемкое образование липидов, представляющих собой главные составные части мембран, зависит от наличия кислорода. При этом часть имеющихся жирных кислот превращается в ненасыщенные жирные кислоты, имеющие более низкую температуру плавления и соответственно обладающие более высокой текучестью. При недостатке кислорода построение клеток преждевременно прекращается.
Стенка клетки полупроницаема. Поступление растворенных веществ (сахаров, аминокислот, жирных кислот, минеральных веществ) происходит избирательно через нерастворимые транспортные протеины, находящиеся в мембране и пропускающие совершенно определенные группы веществ. Выделение наружу продуктов распада, образованного спирта, происходит через мембрану при помощи так называемой везикулы Гольджи.
Совокупность оболочки, состоящей из клеточной мембраны, прикрепленных слоев и гликокаликса, называется стенкой клетки [13]. Клеточная стенка представляет собой жесткую структуру толщиной 25 нм, составляет около 25% сухой массы клетки и состоит в основном из глюкана, маннана, хитина и белка. Организация клеточной стенки недостаточно изучена, однако современные теории отдают предпочтение модели трехслойной структуры, согласно которой внутренний глюкановый слой отделен от внешнего маннанового промежуточным слоем с повышенным содержанием белка.
Морфологическое строение дрожжевой клетки часто оказывает влияние на физиологическое состояние. У молодых клеток оболочка тонкая, протоплазма мелкозернистая. По мере старения клетки протоплазма становится крупнозернистой и количество включений и вакуолей в ней увеличивается. Старые, голодающие клетки обычно находятся в осадке и характеризуются зернистой протоплазмой; мертвые клетки имеют неправильную форму и окрашиваются метиленовым синим [13].
При избытке питательных веществ, например, после начала брожения, дрожжевая клетка запасает резервные вещества. По данным Кунце [13] содержание гликогена может возрасти на 30 % и более от содержания СВ дрожжей; он помещается в аккумулирующих гранулах, расположенных в цитоплазме. Считается, что важна не абсолютная концентрация гликогена, а изменение относительного количества в течение той или иной стадии процесса. Существенное снижение содержания гликогена говорит о неблагополучном состоянии дрожжей. Точно так же, как фосфаты и липиды, которые требуются дрожжам для построения новых веществ клетки, откладывается трегалоза (дисахарид) [13].



