Что такое включения в бактериальной клетке
Один из основных признаков прокариотической клетки — отсутствие внутреннего разделения, обеспечиваемого элементарными мембранами. По сути, единственная мембранная система прокариотической клетки — ЦПМ, входящая в состав клеточной оболочки, часто сложно устроенная и проникающая поперечными складками глубоко в цитоплазму. В составе любой стабильной бактериальной клетки выделяют условные компартменты — поверхностные структуры, клеточную оболочку и цитоплазму.
Поверхностные структуры бактерий
Основные поверхностные структуры бактериальной клетки — капсула, жгутики и микроворсинки. Их наличие — относительно стабильный признак, используемый для идентификации бактерий.
Капсула бактерий
Клеточную оболочку многих бактерий окружает слой аморфного, сильно обводнённого вещества. Этот покров выполняет важные функции: делает оболочку клетки (состоящей из клеточной стенки и ЦПМ) более плотной и прочной, предохраняет бактерии от воздействия бактерицидных факторов, обеспечивает адгезию на различных субстратах, может содержать запасы питательных веществ.
Организация капсул бактерий. Основную роль в организации капсул бактерий играет ЦПМ. Выделяют микрокапсулы (выявляемые только при электронной микроскопии в виде слоя мукополисахаридных микрофибрилл) и макрокапсулы (обнаруживают при световой микроскопии). У некоторых бактерий полимеры клеточной оболочки, выделяемые наружу, свободно располагаются вокруг неё, образуя слизистый слой. Капсулированные бактерии могут превращаться в бескапсульные варианты и, поскольку первые образуют мукоидные или гладкие (S), а бескапсульные — шероховатые (R) колонии, это явление известно как S- и R-диссоциация. Капсула и слизистый слой не препятствуют поступлению и выходу различных веществ из бактериальной клетки, а также плохо удерживают красители.
Окраска капсул бактерий. Для облегчения микроскопирования капсулы можно сделать видимыми, проведя негативную окраску мазка по Бурри-Гинсу или Хиссу либо с помощью реакции набухания по Нойфельду (см. ниже).
Состав капсул бактерий. В состав большинства бактериальных капсул входят сложные полисахарид ды. Капсулы также могут содержать соединения азота (например, у пневмококков капсула состоит из полисахаридов, глюкозамина и глюкуроновой кислоты), но могут и не содержать азот (например, капсулы лейконостоков состоят из декстрана, левулана, фруктозана и других полимеризованных моносахаров). Капсулы некоторых болезнетворных бактерий (например, Bacillus anthracis) формируют полисахариды и полипептиды, образованные мономерами D-глутаминовой кислоты. Поскольку D-аминокислоты устойчивы к воздействию протеаз, такая капсула лучше защищает бактерию от ферментативных воздействий фагоцитов.
Антигенные свойства капсул бактерий. Капсульные Аг (К-Аг) многих патогенных бактерий проявляют выраженные иммуногенные свойства (например, вакцины против пневмококковых и менингококковых инфекций готовят из материала капсулы). Связывание AT с капсулой изменяет её светопреломляющие характеристики, вызывая кажущееся набухание капсулы, видимое при соответствующих условиях освещения под микроскопом и известное как реакция Нойфельда.
Микробиология: конспект лекций

Данная книга предназначена студентам медицинских образовательных учреждений. Это краткое пособие поможет при подготовке и сдаче экзамена по микробиологии. Материал изложен в очень удобной и запоминающейся форме и поможет студентам за сжатый срок детально освоить основные концепции и понятия курса, а также конкретизировать и систематизировать знания.
Оглавление
Приведённый ознакомительный фрагмент книги Микробиология: конспект лекций предоставлен нашим книжным партнёром — компанией ЛитРес.
ЛЕКЦИЯ № 2. Морфология и ультраструктура бактерий
1. Особенности строения бактериальной клетки. Основные органеллы и их функции
Отличия бактерий от других клеток
1. Бактерии относятся к прокариотам, т. е. не имеют обособленного ядра.
2. В клеточной стенке бактерий содержится особый пептидогликан — муреин.
3. В бактериальной клетке отсутствуют аппарат Гольджи, эндоплазматическая сеть, митохондрии.
4. Роль митохондрий выполняют мезосомы — инвагинации цитоплазматической мембраны.
5. В бактериальной клетке много рибосом.
6. У бактерий могут быть специальные органеллы движения — жгутики.
7. Размеры бактерий колеблются от 0,3–0,5 до 5—10 мкм.
По форме клеток бактерии подразделяются на кокки, палочки и извитые.
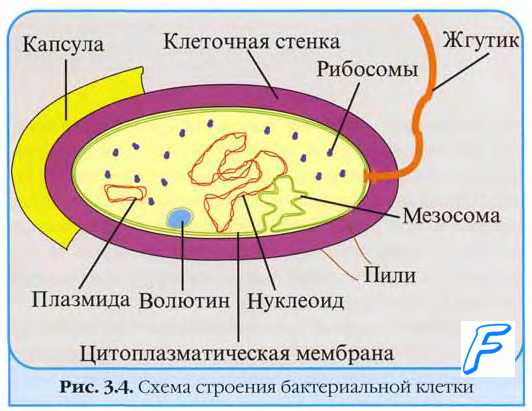
В бактериальной клетке различают:
1) основные органеллы:
г) цитоплазматическую мембрану;
д) клеточную стенку;
2) дополнительные органеллы:
Цитоплазма представляет собой сложную коллоидную систему, состоящую из воды (75 %), минеральных соединений, белков, РНК и ДНК, которые входят в состав органелл нуклеоида, рибосом, мезосом, включений.
Наряду с нуклеоидом в цитоплазме могут находиться автономные кольцевые молекулы ДНК с меньшей молекулярной массой — плазмиды. В них также закодирована наследственная информация, но она не является жизненно необходимой для бактериальной клетки.
Рибосомы представляют собой рибонуклеопротеиновые частицы размером 20 нм, состоящие из двух субъединиц — 30 S и 50 S. Рибосомы отвечают за синтез белка. Перед началом синтеза белка происходит объединение этих субъединиц в одну — 70 S. В отличие от клеток эукариотов рибосомы бактерий не объединены в эндоплазматическую сеть.
Мезосомы являются производными цитоплазматической мембраны. Мезосомы могут быть в виде концентрических мембран, пузырьков, трубочек, в форме петли. Мезосомы связаны с нуклеоидом. Они участвуют в делении клетки и спорообразовании.
Включения являются продуктами метаболизма микроорганизмов, которые располагаются в их цитоплазме и используются в качестве запасных питательных веществ. К ним относятся включения гликогена, крахмала, серы, полифосфата (волютина) и др.
2. Строение клеточной стенки и цитоплазматической мембраны
Клеточная стенка — упругое ригидное образование толщиной 150–200 ангстрем. Выполняет следующие функции:
1) защитную, осуществление фагоцитоза;
2) регуляцию осмотического давления;
4) принимает участие в процессах питания деления клетки;
5) антигенную (определяется продукцией эндотоксина — основного соматического антигена бактерий);
6) стабилизирует форму и размер бактерий;
7) обеспечивает систему коммуникаций с внешней средой;
8) косвенно участвует в регуляции роста и деления клетки.
Клеточная стенка при обычных способах окраски не видна, но если клетку поместить в гипертонический раствор (при опыте плазмолиза), то она становится видимой.
Клеточная стенка вплотную примыкает к цитоплазматической мембране у грамположительных бактерий, у грамотрицательных бактерий клеточная стенка отделена от цитоплазматической мембраны периплазматическим пространством.
Клеточная стенка имеет два слоя:
1) наружный — пластичный;
2) внутренний — ригидный, состоящий из муреина.
В зависимости от содержания муреина в клеточной стенке различают грамположительные и грамотрицательные бактерии (по отношению к окраске по Грамму).
У грамположительных бактерий муреиновый слой составляет 80 % от массы клеточной стенки. По Грамму, они окрашиваются в синий цвет. У грамположительных бактерий муреиновый слой составляет 20 % от массы клеточной стенки, по Грамму, они окрашиваются в красный цвет.
У грамположительных бактерий наружный слой клеточной стенки содержит липопротеиды, гликопротеиды, тейхоевые кислоты, у них отсутствует липополисахаридный слой. Клеточная стенка выглядит аморфной, она не структурирована. Поэтому при разрушении муреинового каркаса бактерии полностью теряют клеточную стенку (становятся протопластами), не способны к размножению.
У грамотрицательных бактерий наружный пластический слой четко выражен, содержит липопротеиды, липополисахаридный слой, состоящий из липида А (эндотоксина) и полисахарида (О-антигена). При разрушении грамотрицательных бактерий образуются сферопласты — бактерии с частично сохраненной клеточной стенкой, не способные к размножению.
К клеточной стенке прилегает цитоплазматическая мембрана. Она обладает избирательной проницаемостью, принимает участие в транспорте питательных веществ, выведении экзотоксинов, энергетическом обмене клетки, является осмотическим барьером, участвует в регуляции роста и деления, репликации ДНК, является стабилизатором рибосом.
Имеет обычное строение: два слоя фосфолипидов (25–40 %) и белки.
По функции мембранные белки разделяют на:
2) пермиазы — белки транспортных систем;
3) энзимы — ферменты.
Липидный состав мембран непостоянен. Он может меняться в зависимости от условий культивирования и возраста культуры. Разные виды бактерий отличаются друг от друга по липидному составу своих мембран.
3. Дополнительные органеллы бактерий
Жгутики — органеллы движения. Есть у подвижных бактерий. Это особые белковые выросты на поверхности бактериальной клетки, содержащие белок — флагелин. Количество и расположение жгутиков может быть различным.
1) монотрихи (имеют один жгутик);
2) лофотрихи (имеют пучок жгутиков на одном конце клетки);
3) амфитрихи (имеют по одному жгутику на каждом конце);
4) перитрихи (имеют несколько жгутиков, расположенных по периметру).
О подвижности бактерий судят, рассматривая живые микроорганизмы, либо косвенно — по характеру роста в среде Пешкова (полужидком агаре). Неподвижные бактерии растут строго по уколу, а подвижные дают диффузный рост.
Капсулы представляют собой дополнительную поверхностную оболочку. Они образуются при попадании микроорганизма в макроорганизм. Функция капсулы — защита от фагоцитоза и антител.
Различают макро — и микрокапсулы. Макрокапсулу можно выявить, используя специальные методы окраски, сочетая позитивные и негативные методы окраски. Микрокапсула — утолщение верхних слоев клеточной стенки. Обнаружить ее можно только при электронной микроскопии. Микрокапсулы характерны для вирулентных бактерий.
Среди бактерий различают:
1) истиннокапсульные бактерии (род Klebsiella) — сохраняют капсулообразование и при росте на питательных средах, а не только в макроорганизме;
2) ложнокапсульные — образуют капсулу только при попадании в макроорганизм.
Капсулы могут быть полисахаридными и белковыми. Они играют роль антигена, могут быть фактором вирулентности.
Споры — это особые формы существования некоторых бактерий при неблагоприятных условиях внешней среды. Спорообразование присуще грамположительным бактериям. В отличие от вегетативных форм споры более устойчивы к действию химических, термических факторов.
Чаще всего споры образуют бактерии рода Bacillus и Clostridium.
Процесс спорообразования заключается в утолщении всех оболочек клетки. Они пропитываются солями дипикалината кальция, становятся плотными, клетка теряет воду, замедляются все ее пластические процессы. При попадании споры в благоприятные условия она прорастает в вегетативную форму.
У грамотрицательных бактерий также обнаружена способность сохраняться в неблагоприятных условиях в виде некультивируемых форм. При этом нет типичного спорообразования, но в таких клетках замедлены метаболические процессы, невозможно сразу получить рост на питательной среде. Но при попадании в макроорганизм они превращаются в исходные формы.

ГК «Униконс»
Продвижение и реализация комплексных пищевых добавок, антисептиков и др. продукции.

«Антисептики Септоцил»
Септоцил. Бытовая химия, антисептики.

«Петритест»
Микробиологические экспресс-тесты. Первые результаты уже через 4 часа.

«АльтерСтарт»
Закваски, стартовые культуры. Изготовление любых заквасок для любых целей.
1.1.4. Морфология и физиология микроорганизмов
Микроорганизмы в зависимости от молекулярно-биологической организации подразделяют на прокариотов и эукариотов.
Одной из основных таксономических категорий является вид совокупность особей, объединенных по близким свойствам, но отличающихся от других представителей рода. Совокупность однородных микроорганизмов, выделенных на питательной среде, характеризующаяся сходными морфологическими, тинкториальными (отношение к красителям), культуральными, биохимическими и антигенными свойствами, называется чистой культурой.
Морфология бактерий в некоторой степени зависит от условий их культивирования, состава питательной среды, факторов окружающей среды и других. В оптимальных условиях культивирования молодые, активно растущие клетки являются наиболее типичными морфологически, в то время как старые могут иметь атипичную морфологию. Бактериальные клетки одного штамма могут отличаться между собой.
Одноклеточные бактерии по внешним признакам могут быть шарообразные (кокки), палочковидные и извитые. На рисунке 1.8 изображены основные формы микробов.
Кокки не всегда имеют правильную круглую форму, для некоторых видов они принимают закругленную, овальную, продолговатую форму.
Например, менингококк (возбудитель эпидемического менингита) имеет форму кофейных зерен, обращенных вогнутой поверхностью друг к другу.
Большинство палочковидных бактерий располагается беспорядочно, так как после деления клетки расходятся.
Многоклеточные бактерии делят на два типа нитчатые и слизистые. Отличаются от одноклеточных более сложным строением и определенным циклом развития.
Бактериальная клетки имеет сложное строение (рис. 1.9). В состав входят клеточная стенка, цитоплазматическая мембрана, цитоплазма с включениями и ядро, называемое нуклеоидом. Бактерии могут иметь и дополнительные структуры: капсулу, микрокапсулу, слизь, жгутики, пили; некоторые бактерии способны образовывать споры.
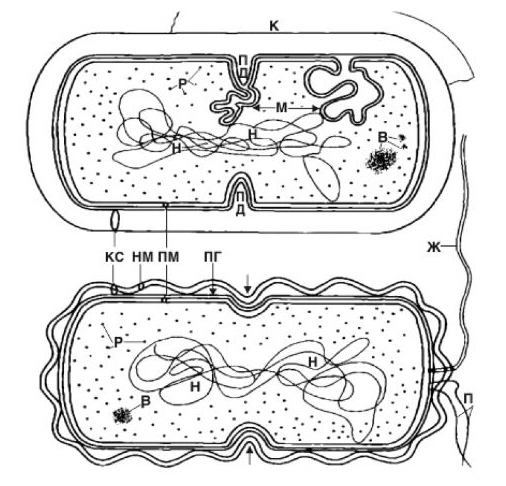
Стрелкой указано деление грамотрицательной бактерии
путем образования перетяжки.
Цитоплазматическая мембрана является трехслойной структурой и окружает наружную часть цитоплазмы бактерий. По структуре она состоит из двойного слоя липидов, главным образом фосфолипидов со встроенными поверхностными и интегральными белками. Цитоплазматическая мембрана является динамической структурой с подвижными компонентами, поэтому ее представляют как мобильную, текучую структуру. Она участвует в регуляции осмотического давления, транспорте веществ и энергетическом метаболизме клетки.
Цитоплазма бактерий коллоидная система, занимает основной объем клетки и состоит из растворимых белков. В молодых клетках она оптически однородна, в более старых имеет зернистость. Основная функция обмен веществ. В цитоплазме имеются различные включения митохондрии, рибосомы, полисахариды, полимасляная кислота и полифосфаты (волютин). Митохондрии выполняют дыхательные функции и анаэробный бродильный распад веществ; в рибосомах происходит биосинтез белков. Волютин выполняет роль запасных питательных веществ. При механическом или ином повреждении цитоплазмы клетка погибает.
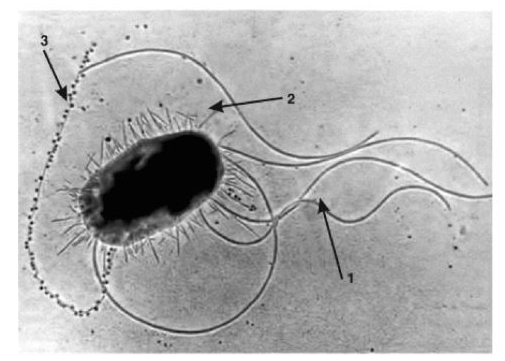
У многих бактерий имеются жгутики, благодаря которым бактерии подвижны. Жгутики представляют собой тонкие нити, отходящие от цитоплазматической мембраны. Жгутики выявляют с помощью электронной микроскопии препаратов, напыленных тяжелыми металлами (рис. 1.10), или в световом микроскопе после обработки препаратов специальными методами (например, после серебрения). Число жгутиков специфично для каждого вида бактерий от одного (холерный вибрион) до десятков, сотен (кишечная палочка, протей). В зависимости от расположения и числа жгутиков различают основные типы жгутикования (рис. 1.11).
Также у бактерий существуют такие образования, как ворсинки и пили, нитевидные тонкие образования. В функции входит прикрепление к поражаемой клетке, или конъюгация.
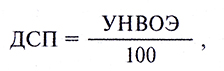
Термин «пили» чаще означает особые ворсинки, способствующие выполнению половых функций.
Из семейства сахаромицетов наибольшее промышленное значение имеет большой род сахаромицес, в который входит множество видов, в том числе дрожжи, применяемые в виноделии, пивоварении, хлебопечении, производстве спирта. Из семейства несахаромицетов наиболее часто встречаются роды торула и кандида, причиняющие значительный ушерб на пищевых предприятиях.
Дрожжи часто обнаруживают на мясе и мясопродуктах; они вызывают ослизнение мяса при хранении, образование пигментных пятен, а осмофильные дрожжи развиваются в рассолах.
В клетках плесени различают клеточную стенку, цитоплазму с включениями, одно или несколько ядер, в отличие от бактерий. Размножаются половым, бесполым путем и вегетативно. Основной способ распространения спорообразование.
Физиология микроорганизмов изучает жизнедеятельность микробных клеток, процессы их питания, дыхания, роста, размножения, закономерности взаимодействия с окружающей средой.
Микроорганизмы не имеют специальных органов питания, питание осуществляется всей поверхностью клетки. Интенсивность проникновения питательных веществ в клетку зависит от степени проницаемости клеточной оболочки, растворимости питательных веществ в воде, разности концентраций питательных веществ в клетке и во внешней среде. Чем сложнее химическое строение вещества, тем хуже оно проникает в клетку.
По способности усвоения углерода микроорганизмы делятся на автотрофные и гетеротрофные. Автотрофы способны самостоятельно продуцировать органические соединения из углекислого газа и солей азотной кислоты, способны существовать только в неорганической среде. Гетеротрофы используют готовые органические соединения (сахар, многоатомные спирты, органические кислоты) для получения углерода, минеральные вещества неспособны использовать в качестве энергетического материала. Нерастворимые источники углерода, такие как жиры, крахмал, также могут быть использованы после расщепления микроорганизмами.
Гетеротрофы делят на метатрофы и наратрофы. К метатрофам в первую очередь относятся сапрофиты, т.е. гнилостные микробы и большинство возбудителей брожения, а также значительное количество патогенных микроорганизмов. Для синтеза белков необходим азот, поэтому источником азота могут быть белковые вещества организмов, а в искусственных питательных средах используются пептоны и аминокислоты. Паратрофы организмы-паразиты, пользующиеся готовыми веществами клетки-хозяина (вирусы, риккетсии).
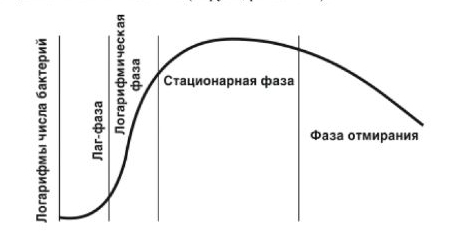
Рис. 1.12. Фазы роста и размножения бактерий
Быстрота размножения микробов изменяется во времени. На жидкой искусственной питательной среде наиболее четко видна динамика развития колонии, которое проходит четыре основные фазы (рис. 1.12). Лаг-фаза, фаза задержки роста, продолжается 3-5 ч, когда бактерии приспосабливаются к новым условиям среды, клетки увеличиваются в объеме, но размножение не происходит. Логарифмическая фаза роста характеризуется усиленным размножением бактерий, скорость появления клеток во много раз превышает скорость отмирания, фаза продолжается 5-6 ч. Во время этой фазы бактерии наиболее уязвимы и наилучшие возможности дня применения уничтожающих их агентов. В стационарной фазе число образующихся клеток постепенно становится равным числу отмирающих, фаза может быть очень продолжительной. Фаза отмирания наступает по мере истощения питательной среды и накопления продуктов обмена. Скорость отмирания варьируется. Часть оставшихся в живых клеток переходят в споры.
К числу основных факторов, оказывающих влияние на рост и размножение микроорганизмов, относятся факторы внешней среды.



