Клеточное дыхание
Клеточное или тканевое дыхание — совокупность биохимических реакций, протекающих в клетках живых организмов, в ходе которых происходит окисление углеводов, липидов и аминокислот до углекислого газа и воды. Высвобожденная энергия запасается в химических связях макроэргических соединений (АТФ и др.) и может быть использована по мере необходимости. Входит в группу процессов катаболизма. О физиологических процессах транспортировки к клеткам многоклеточных организмов кислорода и удалению от них углекислого газа см. статью Дыхание.

Содержание
Использование различных начальных субстратов
В качестве исходных субстратов дыхания могут выступать различные вещества, преобразуемые в ходе специфических метаболических процессов в Ацетил-КоА с высвобождением ряда побочных продуктов. Восстановление НАД (НАДФ) и образование АТФ может происходить уже на этом этапе, однако большая их часть образуется в цикле трикарбоновых кислот при переработке Ацетил-КоА.
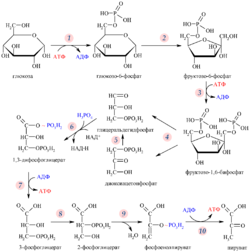
Гликолиз
Гликолиз — путь ферментативного расщепления глюкозы — является общим практически для всех живых организмов процессом. У аэробов он предшествует собственно клеточному дыханию, у анаэробов завершается брожением. Сам по себе гликолиз является полностью анаэробным процессом и для осуществления не требует присутствия кислорода.
Первый его этап протекает с расходом энергии 2 молекул АТФ и включает в себя расщепление молекулы глюкозы на 2 молекулы глицеральдегид-3-фосфата. На втором этапе происходит НАД-зависимое окисление глицеральдегид-3-фосфата, сопровождающееся субстратным фосфорилированием, то есть присоединением к молекуле остатка фосфорной кислоты и формированием в ней макроэргической связи, после которого остаток переносится на АДФ с образованием АТФ.
Таким образом, уравнение гликолиза имеет следующий вид:
Сократив АТФ и АДФ из левой и правой частей уравнения реакции, получим:
Окислительное декарбоксилирование пирувата
Образовавшаяся в ходе гликолиза пировиноградная кислота (пируват) под действием пируватдегидрогеназного комплекса (сложная структура из 3 различных ферментов и более 60 субъединиц) распадается на углекислый газ и ацетальдегид, который вместе с Кофермент А образует Ацетил-КоА. Реакция сопровождается восстановлением НАД до НАД∙Н.
У эукариот процесс протекает в матриксе митохондрий.
β-окисление жирных кислот
Деградация жирных кислот (у некоторых организмов также алканов) происходит у эукариот в матриксе митохондрий. Суть этого процесса заключается в следующем. На первой стадии к жирной кислоте присоединяется кофермент А с образованием ацил-KoA. Он дегидрируется с последовательным переносом восстановительных эквивалентов на убихинон дыхательной ЭТЦ. На второй стадии происходит гидратирование по двойной связи С=С, после чего на третьей стадии происходит окисление полученной гидроксильной группы. В ходе этой реакции восстанавливается НАД.
Наконец, на четвёртой стадии образовавшаяся β-кетокислота расщепляется β-кетотиолазой в присутствии кофермента А на ацетил-КоА и новый ацил-КоА, в которой углеродная цепь на 2 атома короче. Цикл β-окисления повторяется до тех пор, пока вся жирная кислота не будет переработана в ацетил-КоА.
Цикл трикарбоновых кислот
Ацетил-КоА под действием цитратсинтазы передаёт ацетильную группу оксалоацетату с образованием лимонной кислоты, которая поcтупает в цикл трикарбоновых кислот (цикл Кребса). В ходе одного оборота цикла лимонная кислота несколько раз дегидрируется и дважды декарбоксилируется с регенерацией оксалоацетата и образованием одной молекулы ГТФ (способом субстратного фосфорилирования), трёх НАДН и ФАДН2.
Суммарное уравнение реакций:
Ацетил-КоА + 3НАД + + ФАД + ГДФ + Фн + 2H2O + КоА-SH = 2КоА-SH + 3НАДH + 3H + + ФАДН2 + ГТФ + 2CO2
У эукариот ферменты цикла находятся в свободном состоянии в матриксе митохондрий, только сукцинатдегидрогеназа встроена во внутреннюю митохондриальную мембрану.
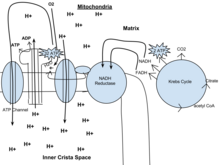
Окислительное фосфорилирование
Основное количество молекул АТФ вырабатывается по способу окислительного фосфорилирования на последней стадии клеточного дыхания: в электронтранспортной цепи. Здесь происходит окисление НАД∙Н и ФАДН2, восстановленных в процессах гликолиза, β-окисления, цикла Кребса и т. д.. Энергия, выделяющаяся в ходе этих реакций, благодаря цепи переносчиков электронов, локализованной во внутренней мембране митохондрий (у прокариот — в цитоплазматической мембране), трансформируется в трансмембранный протонный потенциал. Фермент АТФ-синтаза использует этот градиент для синтеза АТФ, преобразуя его энергию в энергию химических связей. Подсчитано, что молекула НАД∙Н может дать в ходе этого процесса 2.5 молекулы АТФ, ФАДН2 — 1.5 молекулы.
Конечным акцептором электрона в дыхательной цепи аэробов является кислород.
Анаэробное дыхание
Общее уравнение дыхания, баланс АТФ
| Стадия | Выход кофермента | Выход АТФ (ГТФ) | Способ получения АТФ |
|---|---|---|---|
| Первая фаза гликолиза | −2 | Фосфорилирование глюкозы и фруктозо-6-фосфата с использованием 2 АТФ из цитоплазмы. | |
| Вторая фаза гликолиза | 4 | Субстратное фосфорилирование | |
| 2 НАДН | 3 (5) | Окислительное фосфорилирование. Только 2 АТФ образуется из НАДН в электронтранспортной цепи, поскольку кофермент образуется в цитоплазме и должен быть транспортирован в митохондрии. При использовании малат-аспартатного челнока для транспорта в митохондрии из НАДН образуется 3 моль АТФ. При использовании же глицерофосфатного челнока образуется 2 моль АТФ. | |
| Декарбоксилирование пирувата | 2 НАДН | 5 | Окислительное фосфорилирование |
| Цикл Кребса | 2 | Субстратное фосфорилирование | |
| 6 НАДН | 15 | Окислительное фосфорилирование | |
| 2 ФАДН2 | 3 | Окислительное фосфорилирование | |
| Общий выход | 30 (32) АТФ [2] | При полном окислении глюкозы до углекислого газа и окислении всех образующихся коферментов. | |
Примечания
См. также
Полезное
Смотреть что такое «Клеточное дыхание» в других словарях:
клеточное дыхание — — [http://www.dunwoodypress.com/148/PDF/Biotech Eng Rus.pdf] Тематики биотехнологии EN cell respiration … Справочник технического переводчика
Дыхание — Диафрагмальный (брюшной) тип дыхания у человека У этого термина существуют и другие значения, см. Клеточное дыхание … Википедия
ДЫХАНИЕ — Обычно дыхание ассоциируется с вдохом и выдохом, т.е. дыхательными движениями, необходимыми для вентиляции легких у наземных позвоночных. Однако у большинства организмов ни этих движений, ни самих легких нет, поэтому более общее определение… … Энциклопедия Кольера
ДЫХАНИЕ — ДЫХАНИЕ, совокупность процессов, обеспечивающих поступление в организм кислорода и удаление диоксида углерода (внешнее дыхание), а также использование кислорода клетками и тканями для окисления органических веществ с освобождением энергии,… … Современная энциклопедия
ДЫХАНИЕ — совокупность процессов, обеспечивающих поступление в организм кислорода и удаление углекислого газа (внешнее дыхание), а также использование кислорода клетками и тканями для окисления органических веществ с освобождением энергии, необходимой для… … Большой Энциклопедический словарь
Дыхание — ДЫХАНИЕ, совокупность процессов, обеспечивающих поступление в организм кислорода и удаление диоксида углерода (внешнее дыхание), а также использование кислорода клетками и тканями для окисления органических веществ с освобождением энергии,… … Иллюстрированный энциклопедический словарь
ДЫХАНИЕ — ДЫХАНИЕ, я, ср. 1. Процесс поглощения кислорода и выделения углекислого газа живыми организмами. Органы дыхания. Клеточное д. (спец.). 2. Втягивание и выпускание воздуха лёгкими. Ровное д. Сдерживать д. Д. весны (перен.). • Второе дыхание прилив… … Толковый словарь Ожегова
дыхание — ДЫХАНИЕ, ДЫХАНЬЕ, я; ср. 1. Вбирание и выпускание воздуха лёгкими или (у некоторых животных) иными соответствующими органами как процесс поглощения кислорода и выделения углекислого газа живыми организмами. Органы дыхания. Шумное, тяжёлое,… … Энциклопедический словарь
Дыхание — в общеупотребительном смысле обозначает ряд беспрерывно чередующихся во время жизни движений грудной клетки в форме вдоха и выдоха и обусловливающих, с одной стороны, прилив свежого воздуха в легкие, а с другой выведение из них уже испорченного… … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
Дыхание — I Дыхание (respiratio) совокупность процессов, обеспечивающих поступление из атмосферного воздуха в организм кислорода, использование его в биологическом окислении органических веществ и удаление из организма углекислого газа. В результате… … Медицинская энциклопедия


Питательные вещества, которые обычно используются клетками животных и растений при дыхании, включают: сахар, аминокислоты и жирные кислоты, а наиболее распространенные окислитель обеспечивает большую часть химической энергии молекулярной кислород (O2). [1] Химическая энергия, запасенная в АТФ (связь его третьей фосфатной группы с остальной частью молекулы может быть разорвана, позволяя формировать более стабильные продукты, тем самым высвобождая энергию для использования клеткой), затем может использоваться для управления процессами, требующими энергии, включая биосинтез, движение или перенос молекул через клеточные мембраны.
Содержание
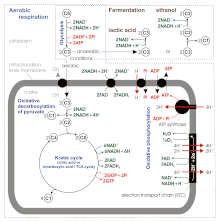
Аэробного дыхания
Аэробного дыхания требует кислород (O2) для создания АТФ. Несмотря на то что углеводы, жиры, и белки расходуются в качестве реагентов, аэробное дыхание является предпочтительным методом пируват распад в гликолиз, и требует пирувата для митохондрии чтобы полностью окислиться цикл лимонной кислоты. Продуктами этого процесса являются углекислый газ и вода, а передаваемая энергия используется для разрыва связей в АДФ, чтобы добавить третью фосфатную группу с образованием АТФ (аденозинтрифосфат), от фосфорилирование на уровне субстрата, НАДН и FADH2
| Упрощенная реакция: | C6ЧАС12О6 (s) + 6 O2 (г) → 6 СО2 (г) + 6 часов2O (л) + тепло |
| ΔG = −2880 кДж на моль C6ЧАС12О6 |
Отрицательное значение ΔG указывает на то, что реакция может происходить самопроизвольно.
Потенциал НАДН и ФАДН2 преобразуется в большее количество АТФ через электронная транспортная цепь с кислородом и протонами (водородом) в качестве «концевых акцепторов электронов». [1] Большая часть АТФ, производимого аэробным клеточным дыханием, производится окислительного фосфорилирования. Энергия O2 [1] Release используется для создания хемиосмотический потенциал накачивая протоны через мембрану. Затем этот потенциал используется для управления АТФ-синтазой и производства АТФ из ADP и фосфатная группа. В учебниках биологии часто говорится, что 38 молекул АТФ могут образовываться на одну окисленную молекулу глюкозы во время клеточного дыхания (2 из гликолиза, 2 из цикла Кребса и около 34 из системы транспорта электронов). [4] Однако этот максимальный выход никогда не достигается полностью из-за потерь из-за протекающих мембран, а также из-за затрат на перемещение пирувата и АДФ в митохондриальный матрикс, и текущие оценки колеблются от 29 до 30 АТФ на глюкозу. [4]
Аэробный метаболизм до 15 раз более эффективен, чем анаэробный метаболизм (который дает 2 молекулы АТФ на 1 молекулу глюкозы), потому что двойная связь в O2 имеет более высокую энергию, чем другие двойные связи или пары одинарных связей в других распространенных молекулах биосферы. [3] Однако некоторые анаэробные организмы, такие как метаногены могут продолжить анаэробное дыхание, производя больше АТФ за счет использования других неорганических молекул (не кислорода) с высокой энергией в качестве конечных акцепторов электронов в цепи переноса электронов. Они разделяют первоначальный путь гликолиз но аэробный метаболизм продолжается с циклом Кребса и окислительным фосфорилированием. Постгликолитические реакции происходят в митохондриях в эукариотические клетки, а в цитоплазма в прокариотические клетки.
Гликолиз
Глюкоза + 2 НАД + + 2 Пя + 2 ADP → 2 пируват + 2 часа + + 2 НАДН + 2 АТФ + 2 Н + + 2 часа2O + энергия
Начиная с глюкозы, 1 АТФ используется для передачи фосфата глюкозе для производства глюкозо-6-фосфата. Гликоген может быть преобразован в глюкозо-6-фосфат также с помощью гликогенфосфорилазы. Во время энергетического обмена глюкозо-6-фосфат становится фруктозо-6-фосфатом. Дополнительный АТФ используется для фосфорилирования фруктозо-6-фосфата в фруктозо-1,6-бисфосфат с помощью фосфофруктокиназы. Затем 1,6-бифосфат фруктозы расщепляется на две фосфорилированные молекулы с тремя углеродными цепями, которые позже распадаются на пируват.
Окислительное декарбоксилирование пирувата
Пируват окисляется до ацетил-КоА и СО.2 посредством пируватдегидрогеназный комплекс (PDC). PDC содержит несколько копий трех ферментов и расположен в митохондрии эукариотических клеток и в цитозоле прокариот. При превращении пирувата в ацетил-КоА одна молекула НАДН и одна молекула СО2 сформирован.
Цикл лимонной кислоты
Это также называется Цикл Кребса или цикл трикарбоновых кислот. Когда присутствует кислород, ацетил-КоА производится из молекул пирувата, созданных в результате гликолиза. однажды ацетил-КоА может возникнуть аэробное или анаэробное дыхание. [6] Когда присутствует кислород, митохондрии подвергаются аэробному дыханию, которое приводит к циклу Кребса. Однако, если кислорода нет, происходит ферментация молекулы пирувата. В присутствии кислорода, когда образуется ацетил-КоА, молекула затем попадает в цикл лимонной кислоты (Цикл Кребса) внутри митохондриального матрикса и окисляется до CO2 в то же время уменьшая НАД к НАДН. НАДН может использоваться электронная транспортная цепь создавать дальше АТФ как часть окислительного фосфорилирования. Чтобы полностью окислить эквивалент одной молекулы глюкозы, два ацетил-КоА должны метаболизироваться в цикле Кребса. Два низкоэнергетических отходы, H2O и CO2, создаются во время этого цикла.
Чистая прибыль от одного цикла составляет 3 НАДН и 1 ФАДН.2 в виде соединений, переносящих водород (протоны плюс электрон), и 1 высокоэнергетический GTP, который впоследствии может быть использован для производства АТФ. Таким образом, общий выход из 1 молекулы глюкозы (2 молекул пирувата) составляет 6 НАДН, 2 ФАДН.2, и 2 АТФ.
Окислительного фосфорилирования
У эукариот окислительное фосфорилирование происходит в митохондриях. кристы. Он включает в себя цепь переноса электронов, которая устанавливает протонный градиент (хемиосмотический потенциал) через границу внутренней мембраны за счет окисления НАДН, образующегося в цикле Кребса. АТФ синтезируется ферментом АТФ-синтаза, когда хемиосмотический градиент используется для управления фосфорилированием АДФ. Перенос электронов осуществляется за счет химической энергии экзогенного кислорода. [1] а при добавлении двух протонов образуется вода.
Эффективность производства АТФ
В таблице ниже описаны реакции, происходящие при полном окислении одной молекулы глюкозы до диоксида углерода. Предполагается, что все уменьшенный коферменты окисляются цепью переноса электронов и используются для окислительного фосфорилирования.
| Шаг | выход кофермента | Выход АТФ | Источник АТФ |
|---|---|---|---|
| Подготовительный этап гликолиза | −2 | Фосфорилирование глюкозы и фруктозо-6-фосфата использует два АТФ из цитоплазмы. | |
| Окупаемость гликолиза | 4 | Фосфорилирование на уровне субстрата | |
| 2 НАДН | 3 или 5 | Окислительное фосфорилирование: каждый НАДН производит чистое 1,5 АТФ (вместо обычных 2,5) за счет транспорта НАДН через митохондриальную мембрану. | |
| Окислительное декарбоксилирование пирувата | 2 НАДН | 5 | Окислительного фосфорилирования |
| Цикл Кребса | 2 | Фосфорилирование на уровне субстрата | |
| 6 НАДН | 15 | Окислительного фосфорилирования | |
| 2 FADH2 | 3 | Окислительного фосфорилирования | |
| Общая доходность | 30 или 32 АТФ | От полного окисления одной молекулы глюкозы до двуокиси углерода и окисления всех восстановленных коферментов. | |
Результатом этих транспортных процессов с использованием протонного электрохимического градиента является то, что более 3 H + необходимы для производства 1 АТФ. Очевидно, это снижает теоретическую эффективность всего процесса, и вероятный максимум находится ближе к 28–30 молекулам АТФ. [4] На практике эффективность может быть даже ниже, потому что внутренняя мембрана митохондрий слегка пропускает протоны. [7] Другие факторы также могут рассеивать протонный градиент, создавая явно протекающие митохондрии. Несвязывающий белок, известный как термогенин экспрессируется в некоторых типах клеток и является каналом, по которому могут переноситься протоны. Когда этот белок активен во внутренней мембране, он замыкает связь между электронная транспортная цепь и Синтез АТФ. Потенциальная энергия протонного градиента не используется для производства АТФ, но генерирует тепло. Это особенно важно в бурый жир термогенез новорожденных и спящих млекопитающих.
Итак, наконец, на молекулу глюкозы
В сумме это дает 4 + 3 (или 5) + 20 + 3 = 30 (или 32) АТФ на молекулу глюкозы.
Эти цифры могут потребовать дальнейшей настройки по мере появления новых деталей конструкции. Приведенное выше значение 3 H + / ATP для синтазы предполагает, что синтаза перемещает 9 протонов и производит 3 ATP за один оборот. Число протонов зависит от количества субъединиц c в Fo c-кольцо, а теперь известно, что это 10 в дрожжах Fo [9] и 8 для позвоночных. [10] Включая один H + для транспортных реакций, это означает, что для синтеза одного АТФ требуется 1 + 10/3 = 4,33 протона у дрожжей и 1 + 8/3 = 3,67 у позвоночных. Это означало бы, что в митохондриях человека 10 протонов от окисления НАДН будут производить 2,72 АТФ (вместо 2,5), а 6 протонов от окисления сукцината или убихинола будут производить 1,64 АТФ (вместо 1,5). Это согласуется с экспериментальными результатами в пределах погрешности, описанными в недавнем обзоре. [11]
Общий выход АТФ в этаноле или молочной кислоте ферментация только 2 молекулы происходят из гликолиз, поскольку пируват не переносится в митохондрия и, наконец, окисляется до диоксида углерода (CO2), но сводится к этиловый спирт или молочная кислота в цитоплазма. [8]
Ферментация
Без кислорода пируват (пировиноградная кислота) не метаболизируется посредством клеточного дыхания, но подвергается процессу ферментации. Пируват не транспортируется в митохондрии, но остается в цитоплазме, где превращается в отходы который может быть удален из камеры. Это служит цели окисления переносчиков электронов, чтобы они могли снова выполнять гликолиз и удалять избыток пирувата. Ферментация окисляет НАДН до НАД + поэтому его можно повторно использовать при гликолизе. В отсутствие кислорода ферментация предотвращает накопление НАДН в цитоплазме и обеспечивает НАД + для гликолиза. Эти отходы различаются в зависимости от организма. В скелетных мышцах отходы молочная кислота. Этот вид брожения называется молочнокислое брожение. При напряженных упражнениях, когда потребность в энергии превышает запас энергии, дыхательная цепь не может обработать все атомы водорода, к которым присоединен NADH. Во время анаэробного гликолиза НАД + восстанавливается, когда пары водорода соединяются с пируватом с образованием лактата. Образование лактата катализируется лактатдегидрогеназой в обратимой реакции. Лактат также может использоваться в качестве косвенного предшественника гликогена печени. Во время восстановления, когда становится доступным кислород, НАД + присоединяется к водороду из лактата с образованием АТФ. В дрожжах отходы этиловый спирт и углекислый газ. Этот тип ферментации известен как алкогольный или ферментация этанола. АТФ, генерируемый в этом процессе, производится фосфорилирование на уровне субстрата, не требующий кислорода.
Ферментация менее эффективно использует энергию глюкозы: на глюкозу вырабатывается только 2 АТФ, по сравнению с 38 АТФ на глюкозу, которые номинально производятся аэробным дыханием. Это потому, что большая часть энергии аэробного дыхания происходит от O2 с его относительно слабой высокоэнергетической двойной связью. [3] [1] Однако гликолитический АТФ создается быстрее. Чтобы прокариоты продолжали быстро расти, когда они переходят из аэробной среды в анаэробную, они должны увеличивать скорость гликолитических реакций. Для многоклеточных организмов во время коротких всплесков напряженной активности мышечные клетки используют ферментацию, чтобы дополнить выработку АТФ из-за более медленного аэробного дыхания, поэтому ферментация может использоваться клеткой даже до того, как уровень кислорода истощится, как это имеет место в спорте. не требовать от спортсменов темпов, например спринт.
Анаэробное дыхание
Анаэробное дыхание используется некоторыми микроорганизмами, у которых ни кислород (аэробное дыхание), ни производные пирувата (ферментация) не являются конечными акцепторами электронов с высокой энергией. Скорее неорганический акцептор, такой как сульфат (SO42-), нитрат (NO3–), или сера (S) используется. [12] Такие организмы обычно встречаются в необычных местах, таких как подводные пещеры или поблизости от них. гидротермальные источники на дне океана.
В июле 2019 года было проведено научное исследование Кидд Майн в Канаде обнаружил серодышащие организмы которые живут на глубине 7900 футов под поверхностью и дышат серой, чтобы выжить. Эти организмы также примечательны тем, что потребляют такие минералы, как пирит в качестве источника пищи. [13] [14] [15]
Дыхание
Эволюция дыхательной системы
Всё живое на Земле существует за сёт солнечного тепла и энергии, достигающей поверхности нашей планеты. Все животные и человек приспособились добывать энергию из синтезированных растениями органических веществ. Чтобы использовать энергию Солнца, заключённую в молекулах органических веществ, её необходимо высвободить, окислив эти вещества. Чаще всего в качестве окислителя используют кислород воздуха, благо он составляет почти четверть объёма окружающей атмосферы.
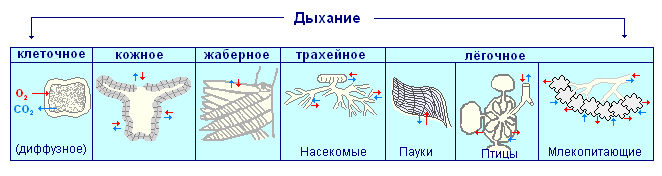
Одноклеточные простейшие животные, кишечнополостные, свободноживущие плоские и круглые черви дышат всей поверхностью тела. Специальные органы дыхания — перистые жабры появляются у морских кольчатых червей и у водных членистоногих. Органами дыхания членистоногих являются трахеи, жабры, листовидные лёгкие расположенные в углублениях покрова тела. Система органов дыхания ланцетника представлена жаберными щелями, пронизывающими стенку переднего отдела кишечника — глотку. У рыб под жаберными крышками располагаются жабры, обильно пронизанными мельчайшими кровеносными сосудами. У наземных позвоночных органами дыхания являются лёгкие. Эволюция дыхания у позвоночных шла по пути увеличения площади легочных перегородок, участвующих в газообмене, совершенствования транспортных систем доставки кислорода к клеткам, расположенным внутри организма, и развития систем, обеспечивающих вентиляцию органов дыхания.
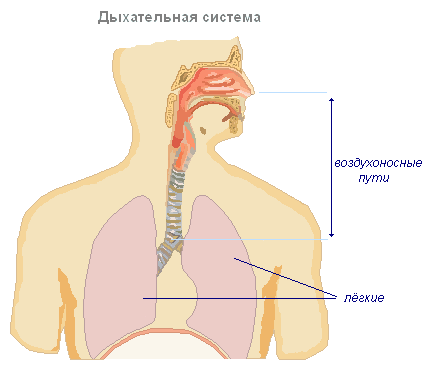
Строение и функции органов дыхания
Необходимым условием жизнедеятельности организма является постоянный газообмен между организмом и окружающей средой. Органы, по которым циркулируют вдыхаемый и выдыхаемый воздух, объединяются в дыхательный аппарат. Систему органов дыхания образуют носовая полость, глотка, гортань, трахея, бронхи и лёгкие. Большинство из них представляют собой воздухоносные пути и служат для проведения воздуха в лёгкие. В лёгких и происходят процессы газообмена. При дыхании организм получает из воздуха кислород, который разносится кровью по всему телу. Кислород участвует в сложных окислительных процессах органических веществ, при котором освобождается необходимая организму энергия. Конечные продукты распада — углекислота и частично вода — выводятся из организма в окружающую среду через органы дыхания.
Функции дыхательной системы
Носовая полость
Воздухоносные пути начинаются с носовой полости, которая через ноздри соединяется с окружающей средой. От ноздрей воздух проходит по носовым ходам, выстланным слизистым, реснитчатым и чувствительным эпителием. Наружный нос состоит из костных и хрящевых образований и имеет форму неправильной пирамиды, которая изменяется в зависимости от особенностей строения человека. В состав костного скелета наружного носа входят носовые косточки и носовая часть лобной кости. Хрящевой скелет является продолжением костного скелета и состоит из гиалиновых хрящей различной формы. Полость носа имеет нижнюю, верхнюю и две боковые стенки. Нижняя стенка образована твёрдым нёбом, верхняя — решётчатой пластинкой решётчатой кости, боковая — верхней челюстью, слёзной костью, глазничной пластинкой решётчатой кости, нёбной костью и клиновидной костью. Носовой перегородкой полость носа разделена на правую и левую части. Перегородка носа образована сошником, перпендикулярной пластинкой решётчатой кости и спереди дополняется четырёхугольным хрящом носовой перегородки.
На боковых стенках полости носа располагаются носовые раковины — по три с каждой стороны, что увеличивает внутреннюю поверхность носа, с которой соприкасается вдыхаемый воздух.
Носовая полость образована двумя узкими и извилистыми носовыми ходами. Здесь воздух согревается, увлажняется и освобождается от частичек пыли и микробов. Оболочка, выстилающая носовые ходы, состоит из клеток, которые выделяют слизь, и клеток реснитчатого эпителия. Движением ресничек слизь вместе с пылью и микробами направляется из носовых ходов наружу.
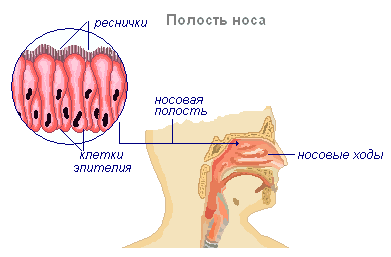
Внутренняя поверхность носовых ходов богато снабжена кровеносными сосудами. Вдыхаемый воздух, попадает в полость носа, обогревается, увлажняется, очищается от пыли и частично обезвреживается. Из носовой полости он попадает в носоглотку. Затем воздух из носовой полости попадает в глотку, а из неё — в гортань.
Гортань
Гортань — один из отделов воздухоносных путей. Сюда из носовых ходов через глотку поступает воздух. В стенке гортани есть несколько хрящей: щитовидный, черпаловидный и др. В момент глотания пищи мышцы шеи поднимают гортань, а надгортанный хрящ опускается и закрывается гортань. Поэтому пища поступает только в пищевод и не попадает в трахею.
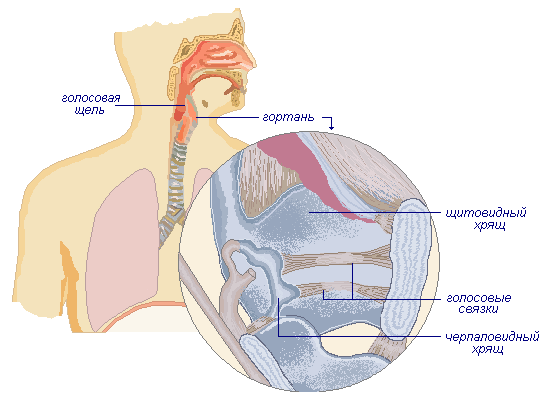
В узкой части гортани расположены голосовые связки, посредине между ними находится голосовая щель. При прохождении воздуха голосовые связки вибрируют, производя звук. Образование звука происходит на выдохе при управляемом человеком движении воздуха. В формировании речи участвуют: носовая полость, губы, язык, мягкое нёбо, мимические мышцы.
Трахея
Гортань переходит в трахею (дыхательное горло), которая имеет форму трубки длиной около 12 см, в стенках которого есть хрящевые полукольца, не позволяющие ей спадать. Задняя стенка её образована соединительнотканной перепонкой. Полость трахеи, как и полость других воздухоносных путей выстлана мерцательным эпителием, препятствующим проникновению в лёгкие пыли и других инородных тел. Трахея занимает серединное положение, сзади она прилежит к пищеводу, а по бокам от неё располагаются сосудисто-нервыне пучки. Спереди шейный отдел трахеи прикрывают мышцы, а вверху она охватывается ещё щитовидной железой. Грудной отдел трахеи прикрыт спереди рукояткой грудины, остатками вилочковой железы и сосудами. Изнутри трахея покрыта слизистой оболочкой, содержащей большое количество лимфоидной ткани и слизистых желёз. При дыхании мелкие частички пыли прилипают к увлажнённой слизистой оболочке трахеи, а реснички мерцательного эпителия продвигают их обратно к выходу из дыхательных путей.
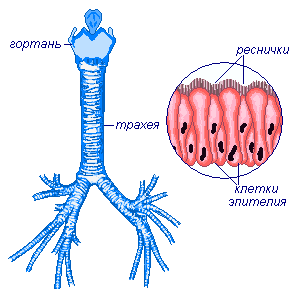
Нижний конец трахеи делится на два бронха, которые затем многократно ветвятся, входят в правое и левое лёгкие, образуя в лёгких «бронхиальное дерево».
Бронхи
В грудной полости трахея делится на два бронха — левый и правый. Каждый бронх входит в лёгкое и там делится на бронхи меньшего диаметра, которые разветвляются на мельчайшие воздухоносные трубочки — бронхиолы. Бронхиолы в результате дальнейшего ветвления переходят в расширения — альвеолярные ходы, на стенках которых находятся микроскопические выпячивания, называемые легочными пузырьками, или альвеолами.
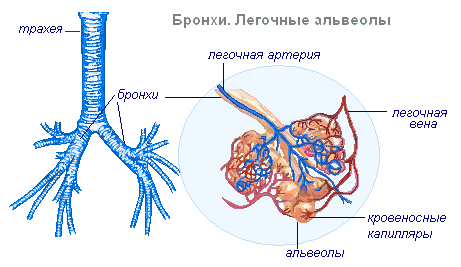
Лёгкие
Лёгкие занимают почти всю полость грудной полости и представляют собой упругие губчатые органы. В центральной части лёгкого располагаются ворота, куда входят бронх, легочная артерия, нервы, а выходят легочные вены. Правое лёгкое делится бороздами на три доли, левое на две. Снаружи лёгкие покрыты тонкой соединительнотканной плёнкой — легочной плеврой, которая переходит на внутреннею поверхность стенки грудной полости и образует пристенную плевру. Между этими двумя плёнками находится плевральная щель, заполненная жидкостью, уменьшающей трение при дыхании.
На лёгком различают три поверхности: наружную, или рёберную, медиальную, обращённую в сторону другого лёгкого, и нижнюю, или диафрагмальную. Кроме того, в каждом лёгком различают два края: передний и нижний, отделяющие диафрагмальную и медиальную поверхности от рёберной. Сзади рёберная поверхность без резкой границы переходит в медиальную. Передний край левого лёгкого имеет сердечную вырезку. На медиальной поверхности лёгкого располагаются его ворота. В ворота каждого лёгкого входит главный бронх, легочная артерия, которая несёт в лёгкое венозную кровь, и нервы, иннервирующие лёгкое. Из ворот каждого лёгкого выходят две легочные вены, которые несут к сердцу артериальную кровь, и лимфатические сосуды.
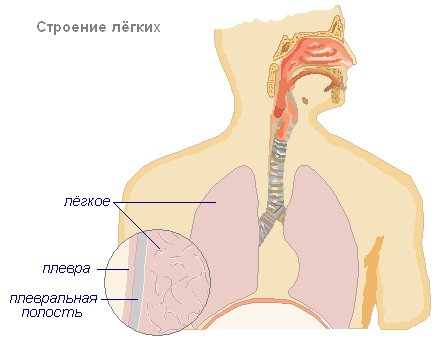
Лёгкие имеют глубокие борозды, разделяющие их на доли — верхнюю, среднюю и нижнюю, а в левом две — верхнюю и нижнюю. Размеры лёгкого не одинаковы. Правое лёгкое несколько больше левого, при этом оно короче его и шире, что соответствует более высокому стоянию правого купола диафрагмы в связи с правосторонним расположением печени. Цвет нормальных лёгких в детском возрасте бледно-розовый, а у взрослых они приобретают тёмно-серую окраску с синеватым оттенком — следствие отложения в них попадающих с воздухом пылевых частиц. Ткань лёгкого мягкая, нежная и пористая.
Газообмен лёгких
В сложном процессе газообмена выделяют три основные фазы: внешнее дыхание, перенос газа кровью и внутреннее, или тканевое, дыхание. Внешнее дыхание объединяет все процессы, происходящие в лёгком. Оно осуществляется дыхательным аппаратом, к которому относятся грудная клетка с мышцами, приводящими её в движение, диафрагма и лёгкие с воздухоносными путями.
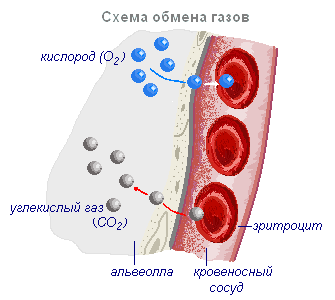
Воздух, поступивший в лёгкие при вдохе, изменяет свой состав. Воздух в лёгких отдаёт часть кислорода и обогащается углекислым газом. Содержание углекислого газа в венозной крови выше, чем в воздухе, находящемся в альвеолах. Поэтому углекислый газ выходит из крови в альвеолы и содержание его меньше, чем в воздухе. Сначала кислород растворяется в плазме крови, далее связывается с гемоглобином, а в плазму поступают новые порции кислорода.
Переход кислорода и углекислого газа из одной среды в другую проходит благодаря диффузии от большей концентрации к меньшей. Хотя диффузия протекает медленно, поверхность контакта крови с воздухом в лёгких настолько велика, что полностью обеспечивает нужный газообмен. Подсчитано, что полный газообмен между кровью и альвеолярным воздухом может происходить за время, которое втрое короче, чем время пребывания крови в капиллярах (т.е. в организме имеются значительные резервы обеспечения тканей кислородом).
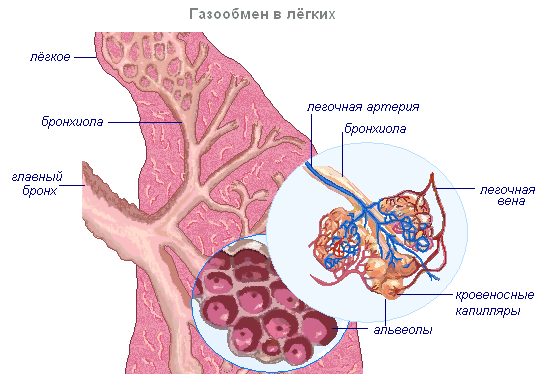
Венозная кровь, попав в лёгкие, отдаёт углекислый газ, обогащается кислородом и превращается в артериальную. В большом круге эта кровь расходится по капиллярам во все ткани и отдаёт кислород клеткам тела, которые постоянно потребляют его. Углекислого газа, выделяющегося клетками в результате их жизнедеятельности, здесь больше, чем в крови, и он диффундирует из тканей в кровь. Таким образом, артериальная кровь, пройдя через капилляры большого круга кровообращения, становится венозной и правой половиной сердца направляется в лёгкие, здесь опять насыщается кислородом и отдаёт углекислый газ.
В организме дыхание осуществляется с помощью дополнительных механизмов. Жидкие среды, входящие в состав крови (её плазмы), обладают низкой растворимостью в них газов. Поэтому, для того чтобы человек мог существовать, ему нужно было бы иметь сердце мощнее в 25 раз, лёгкие — в 20 раз и за одну минуту перекачивать более 100 литров жидкости (а не пять литров крови). Природа нашла способ преодоления этой трудности, приспособив для переноса кислорода особое вещество — гемоглобин. Благодаря гемоглобину кровь способна связывать кислород в 70 раз, а углекислый газ — в 20 раз больше, чем жидкая часть крови — её плазма.
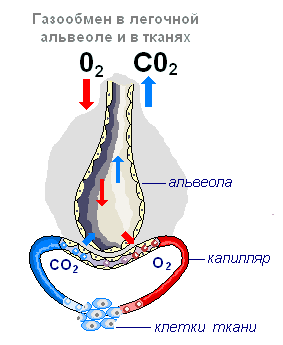
Альвеола — тонкостенный пузырёк диаметром 0,2 мм, заполненный воздухом. Стенка альвеолы образована одним слоем плоских клеток эпителия, по наружной поверхности которых разветвляется сетка капилляров. Таким образом, газообмен происходит через очень тонкую перегородку, образованную двумя слоями клеток: стенки капилляра и стенки альвеолы.
Обмен газов в тканях (тканевое дыхание)
Обмен газов в тканях осуществляется в капиллярах по тому же принципу, что и в лёгких. Кислород из тканевых капилляров, где его концентрация высока, переходит в тканевую жидкость с более низкой концентрацией кислорода. Из тканевой жидкости он проникает в клетки и сразу же вступает в реакции окисления, поэтому в клетках практически нет свободного кислорода.
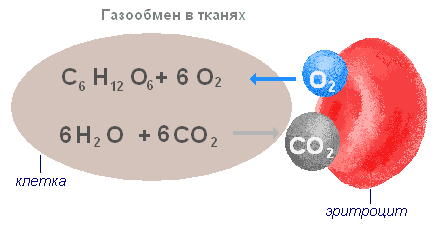
Диоксид углерода по тем же законам поступает из клеток, через тканевую жидкость, в капилляры. Выделяющийся углекислый газ способствует диссоциации оксигемоглобина и сам вступает в соединение с гемоглобином, образуя карбоксигемоглобин, транспортируется в лёгкие и выделяется в атмосферу. В оттекающей от органов венозной крови углекислый газ находится как в связанном, так и в растворённом состоянии в виде угольной кислоты, которая в капиллярах лёгких легко распадается на воду и углекислый газ. Угольная кислота может также вступать в соединения с солями плазмы, образуя бикарбонаты.
В лёгких, куда поступает венозная кровь, кислород снова насыщает кровь, а углекислый газ из зоны высокой концентрации (легочных капилляров) переходит в зону низкой концентрации (альвеол). Для нормального газообмена воздух в лёгких постоянно сменяться, что достигается ритмическими атаками вдоха и выдоха, за счёт движений межрёберных мышц и диафрагмы.
Транспорт кислорода в организме
| Путь кислорода | Функции |
| Верхние дыхательные пути | |
| Носовая полость | Увлажнение, согревание, обеззараживание воздуха, удаление частиц пыли |
| Глотка | Проведение согретого и очищенного воздуха в гортань |
| Гортань | Проведение воздуха из глотки в трахею. Защита дыхательных путей от попадания пищи надгортанным хрящом. Образование звуков путём колебания голосовых связок, движения языка, губ, челюсти |
| Трахея | Свободное продвижение воздуха |
| Бронхи | Свободное продвижение воздуха |
| Лёгкие | Органы дыхания. Дыхательные движения осуществляются под контролем центральной нервной системы и гуморального фактора, содержащегося в крови, — СО2 |
| Альвеолы | Увеличивают площадь дыхательной поверхности, осуществляют газообмен между кровью и лёгкими |
| Кровеносная система | |
| Капилляры лёгких | Транспортируют венозную кровь из легочной артерии в лёгкие. По законам диффузии О2 поступает из мест большей концентрации (альвеолы) в места меньшей концентрации (капилляры), в то же время СО2 диффундирует в противоположном направлении. |
| Легочная вена | Транспортирует О2 от лёгких к сердцу. Кислород, попав в кровь, сначала растворяется в плазме, затем соединяется с гемоглобином, и кровь становится артериальной |
| Сердце | Проталкивает артериальную кровь по большому кругу кровообращения |
| Артерии | Обогащают кислородом все органы и ткани. Легочные артерии несут венозную кровь к лёгким |
| Капилляры тела | Осуществляют газообмен между кровью и тканевой жидкостью. О2 переходит в тканевую жидкость, а СО2 диффундирует в кровь. Кровь становится венозной |
| Клетка | |
| Митохондрии | Клеточное дыхание — усвоение О2 воздуха. Органические вещества благодаря О2 и дыхательным ферментам окисляются (диссимиляция) конечные продукты — Н2О, СО2 и энергия которая идёт на синтез АТФ. Н2О и СО2 выделяются в тканевую жидкость, из которой диффундируют в кровь. |
Дыхание — это совокупность физиологических процессов, обеспечивающих газообмен между организмом и внешней средой (внешнее дыхание), и окислительных процессов в клетках, в результате которых выделяется энергия (внутреннее дыхание). Обмен газов между кровью и атмосферным воздухом (газообмен) — осуществляется органами дыхания.
Источником энергии в организме служат пищевые вещества. Основным процессом, освобождающим энергию этих веществ, является процесс окисления. Он сопровождается связыванием кислорода и образованием углекислого газа. Учитывая, что в организме человека нет запасов кислорода, непрерывное поступление его жизненно необходимо. Прекращение доступа кислорода в клетки организма ведёт к их гибели. С другой стороны, образованный в процессе окисления веществ углекислый газ должен быть удалён из организма, так как накопление значительного количества его опасно для жизни. Поглощение кислорода из воздуха и выделение углекислого газа осуществляется через систему органов дыхания.
Биологическое значение дыхания заключается в:



