Нонсенс-мутации
Нонсенс-мутация — точечная мутация в последовательности ДНК, которая приводит к появлению стоп-кодона (нонсенс-кодона) в транскрибованих мРНК. Результатом является неполный и обычно нефункциональный продукт этой мРНК. Миссенс-мутация — это точечная мутация, когда нуклеотидная замена приводит к трансляции мРНК другой аминокислоты. Некоторые генетические болезни, например талассемия и словесная диспраксия, является результатом нонсенс-мутаций.
Простой пример
Допустим, мы имеем нонсенс-мутацию в четвертом триплете в последовательности ДНК (ЦГА), которая заключается в замене цитозина в тимин. Результатом является триплет ТГА в последовательности ДНК. Поскольку ТГА транскрибируется как УГА, результирующим будут следующий мРНК-транскрипт и белок:
Остальные кодонов после стоп-кодона не транслируются. Результатом является сокращенный белковый продукт, который часто не способен выполнять присущую ему функцию.
Нонсенс-обусловленный расписание мРНК
Несмотря на ожидаемую тенденцию ранних кодонов терминации вызывать трансляцию укороченных белков, на самом деле это не часто происходит in vivo. Многие организмы, например человек и дрожжи, используют механизм под названием «нонсенс-обусловленный расписание мРНК», который приводит к деградации мРНК, которые содержат нонсенс-мутации, перед тем как они будут транслироваться в нефункциональные белки.
Патологии, ассоциированные с нонсенс-мутациями
Нонсенс-мутации могут привести к генетическим болезням, повреждая гены, кодирующие специфические протеины. Примеры болезней, причиной которых могут быть нонсенс-мутации включают:
Полезными в лечении вышеприведенных болезней могут быть экспериментальные лекарства под названием PTC124 (в случаях, когда причиной нонсенс-мутация). PTC124 по расписанию должен был проходить финальную стадию испытаний в 2007 году.
Как нонсенс-мутации ген обижали, и что потом было
Автор
Редакторы
Статья на конкурс «био/мол/текст»: Мутации часто появляются совсем некстати и обижают гены живых организмов, ухудшая им жизнь. К счастью, организмы научились с ними справляться благодаря огромной помощи естественного отбора и некоторых молекулярных механизмов. Но что же произойдет, если ген обидит самая вредная и коварная из мелких мутаций — нонсенс-мутация? И сумеет ли ген найти способ, чтобы перехитрить ее?

Конкурс «био/мол/текст»-2018
Эта работа опубликована в номинации «Своя работа» конкурса «био/мол/текст»-2018.
Генеральный спонсор конкурса — компания «Диаэм»: крупнейший поставщик оборудования, реагентов и расходных материалов для биологических исследований и производств.
Спонсором приза зрительских симпатий выступил медико-генетический центр Genotek.
В генах живых организмов постоянно происходят мутации. Чаще всего они связаны с ошибками, происходящими во время репликации, то есть удвоения ДНК, которое необходимо при делении клетки, и с ошибками при восстановлении повреждений ДНК, которые могут появиться по разнообразным причинам. Почти всегда они чинятся, но все же остаются случаи, когда вся молекулярная машинерия бессильна, и ошибка остается неисправленной. Тогда возникает мутация. Она может быть и в кодирующей, и в некодирующей частях генома, но давайте сосредоточим свое внимание только на первой, то есть на белок-кодирующих генах. Вдобавок, будем рассматривать только точечные мутации, которые заменяют лишь один нуклеотид.
Многие люди заведомо считают, что мутации всегда приносят только вред. На самом деле это верно, но не полностью. Есть «молчащие», или синонимичные мутации, которые не меняют кодируемую аминокислоту. Например, CGT, CGC, CGA, CGG кодируют аргинин и любая замена в третьей позиции кодона им не страшна. Такие мутации вреда не приносят. Другие мутации называются несинонимичными и меняют аминокислоту. Крайний, и как считается, самый вредный их вариант — нонсенс-мутация. О ней мы и будем говорить.
Но нужно начать с рассказа о самой сути нонсенс-мутаций. В начале транслируемой части каждого гена есть старт-кодон ATG (однако когда он расположен не в начале, то кодирует аминокислоту метионин), а в конце — стоп-кодон (один из трех: TAA, TAG или TGA), и для рибосомы они, словно дорожная разметка, обозначают откуда нужно начать синтезировать белок, а где — завершить. Поэтому синтез белка, трансляция, идет от старт-кодона до стоп-кодона. Но если из-за мутации любой кодон между настоящим стартом и стоп-кодоном превратится в стоп-кодон, то трансляция будет заканчиваться именно на этом, преждевременном стоп-кодоне. Так он и называется, «преждевременный стоп-кодон», или уже знакомым нам термином «нонсенс-мутация». По ее достижении трансляция обрывается и получается не хороший рабочий белок, а нефункциональный полипептид. Эффект от этого может и не проявиться в фенотипе, то есть не изменить никакой признак организма. Например, если организм по данному гену гетерозигота и одной функциональной копии гена для него достаточно. Но в некоторых случаях, в том числе если обе копии несут нонсенс-мутацию, или организм гаплоидный (второй копии гена у него попросту нет), или ген важный, то эффект виден невооруженным глазом.
В структуре белок-кодирующего гена можно выделить экзоны и интроны. Такое подразделение касается только ядерных организмов, а так как мы в статье говорим именно о них, то не станем обсуждать другие варианты строения гена. Информация о белке закодирована только в экзонах. Бывают гены, которые состоят всего из одного экзона, их меньшинство. А бывают гены, состоящие из нескольких десятков экзонов, которые при этом могут в разных сочетаниях давать разные белки — это называется «альтернативный сплайсинг». Например, если есть 5 экзонов [A, B, C, D, E], то они могут давать разные изоформы: и [A, B, C, D, E], и [A, B, C], и [B, C, E], и [A, C, D]. Таким образом достигается большое количество отличающихся по функциям белков, кодируемых одним геном.

Давно известно, что в геномах организмов присутствуют псевдогены — это участки, которые раньше были генами, но из-за мутации перестали таковыми быть и не кодируют функционирующий рабочий белок. Их еще называют мусорной ДНК [1]. Причиной может быть любая мутация — от однонуклеотидной, приводящей, например, к несинонимичной или нонсенс-мутации, до большой делеции, то есть удаления почти всего гена, или вставки в него транспозона, превращающего ген в псевдоген. В случае однонуклеотидной мутации или небольших делеции или инсерции можно посмотреть, как этот псевдоген появился и что с ним происходит. Нередко выяснялось, что после поломки ген начинал накапливать несинонимичные и синонимичные мутации с одинаковой скоростью. Это явление называется нейтральной эволюцией. Правда, по результатам было заметно, что в некоторых случаях из-за молодого возраста в псевдогене вообще не произошло никаких мутаций. А порой бывший ген просто не получалось найти, потому что он «старый» и сильно мутировал. Ученые описали нейтральную эволюцию для отдельных случаев, например, для семейств альфа- и бета-глобиновых генов [2], [3] или псевдогенов обонятельных рецепторов [4]. Но насколько наблюдаемое явление распространяется на все псевдогены, известно не было.
Ничто так не подстегивает ученых сделать обобщающее исследование, как противоречивость имеющейся информации. Разумеется, существует немало статей, в которых нонсенс-мутацию в гене ассоциировали с причиной развития заболевания [5], [6]. И стоит ожидать, что после такой поломки ген станет настоящим псевдогеном и начнет эволюционировать нейтрально. С другой стороны, есть статьи [7], в которых указывают на преимущество, получаемое организмом благодаря нонсенс-мутации. Выходит, здесь нонсенс-мутация благоприятна, и такой аллель не станет превращаться в псевдоген. А для некоторых генов вообще показали, что они — псевдо-псевдогены! [8] В них происходит редкое явление «чтение сквозь стоп-кодон», когда трансляция не обрывается, а продолжается дальше до настоящего стоп-кодона, и ген остается функциональным. Но это все же нечастое событие, и псевдогены в большинстве своем функций не несут.
Здесь мы, наконец, добрались до великой силы эволюции — естественного отбора. Он как раз и является силой, которая помогает генам не превратиться в бессмысленные бесполезные последовательности. Отбор очищает популяцию от вредных замен, в том числе и от интересных нам нонсенс-мутаций. Под столь возвышенной фразой имеется в виду лишь то, что особи с вредными мутациями либо не появляются на свет, потому что нежизнеспособны, либо не дают потомство. По очевидным причинам ген с «плохой» мутацией ну никак не может распространиться в популяции и удаляется из нее отбором.
В функционально важных генах естественный отбор поддерживает консервативность, потому как от их правильной работы зависит жизнь организма. И в них синонимичных мутаций накапливается намного больше, чем несинонимичных. В таких случаях говорят, что на ген действует отрицательный отбор, который как раз убирает несинонимичные замены. Повторю, многих генов с бóльшим числом несинонимичных мутаций мы не видим, потому как особи с ними не могут жить, либо не оставляют потомства. С другой стороны, есть меньшее число генов, в которых число несинонимичных замен больше числа синонимичных. Это указывает на работу положительного отбора, то есть наоборот хорошо, что ген сильно мутирует и аминокислоты в нем меняются. Например, так обстоит дело с генами главного комплекса гистосовместимости [9–11]. Третий вариант — равное соотношение синонимичных и несинонимичных замен. Это знак того, что отбору все равно, что происходит с геном, и он уже не ген вовсе, а псевдоген.
Зная о видах отбора и задумываясь о том, насколько противоречивы знания ученого мира о псевдогенах, мы решили установить порядок и понять, что же все-таки с ними происходит. И выбрали для этого модельный объект эволюционных биологов и генетиков — плодовую мушку Drosophila melanogaster. Поскольку она любимый объект биологов, о ней есть достаточно подходящей нам информации (что, на самом деле, даже в эпоху Больших данных иногда становится огромной проблемой и может заставить отложить исследование на некоторый срок). Поэтому решено было взять самый большой набор данных для одной замбийской популяции. Мы решили изучить случаи, когда стоп-кодон возникает из-за однонуклеотидной мутации. Скажем, был нормальный глутаминовый триплет CAA, а стал стоп-кодон TAA.
Чтобы узнать, что происходит с геном после появления нонсенс-мутации, мы решили посмотреть на силу естественного отбора в нем — на отношение несинонимичных замен к синонимичным, по-научному оно называется pN/pS. Здесь логика простая — если это значение будет равно 1, то мы наблюдаем искомую нейтральную эволюцию, и перед нами яркий пример псевдогенизации гена. А вот если оно будет ниже 1, то мы уже начинаем наблюдать отрицательный отбор, при котором несинонимичным мутациям не позволяется копиться с той же скоростью, что и синонимичным. А так как нас интересовало то, что происходит с геном после появления нонсенс-мутации, мы учитывали только те замены, которые появились именно после. Здесь слово «после» означает временные масштабы и указывает на то, что произошло с геном «потом». Это проще объяснить, глядя на рисунок ниже и объяснение в подписи к нему.
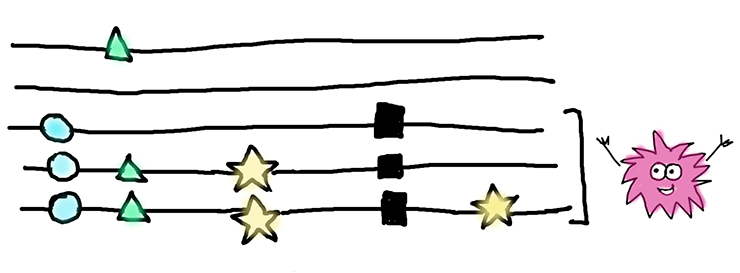
Перед нами — выравнивание для пяти последовательностей, или аллелей, одного гена. Черными квадратами отмечены нонсенс-мутации. Для нашей статистики интересны только замены, изображенные звездочками — именно они произошли после того, как в этих аллелях появилась нонсенс-мутация. Замены, изображенные кругами и треугольниками, скорее всего, произошли до нонсенс-мутации, и мы их в расчет не брали. Те, которые отмечены кругами встречаются во всех последовательностях, несущих нонсенс-мутацию, и, вероятнее всего, присутствовали в гене еще до нее. А мутации, обозначенные треугольниками, есть и у одной «здоровой» последовательности — они явно появились намного раньше нонсенса.
Казалось бы, у нас все есть — и данные, и статистика, которая поможет в подсчетах. Поэтому не терпится скорее приступить к анализу и получить ответ на терзающий душу ученого вопрос! И все же перед тем как брать все имеющиеся у нас нонсенс-аллели, то есть варианты гена с нонсенс-мутациями, и смотреть на замены внутри них, нужно эти аллели отфильтровать. Такой шаг может показаться странным, ведь нонсенс-аллелей и так мало (потому что они вредны и изымаются отрицательным отбором), куда еще фильтры использовать?! Но для того чтобы получить разумный результат, мы должны понимать, что работаем действительно с настоящими псевдогенами. Для генов, как мы уже обсудили, вообще-то нехорошо становиться псевдогенами. Поэтому все вредные мутации должны быть очень редкими в популяции, то есть, говоря научным языком, находиться на низкой частоте. А если мы видим потенциально вредную мутацию, но она присутствует у многих, то это говорит о том, что она безвредна, и организмам, у которых она есть, преспокойно живется. Иначе отрицательный отбор не позволил бы такой мутации добраться до высоких частот. Исходя из этого, в нашем исследовании мы брали во внимание только те нонсенс-мутации, которые встречались у небольшого количества мух, то есть были низкочастотными.
И, конечно, не нужно брать в расчет те нонсенс-мутации, которые расположены в самом начале и самом конце гена. Для расположенных в самом начале легко может оказаться так, что следующий за нонсенс-мутацией кодон будет метионином и сможет использоваться как альтернативный старт, а белок никакого вреда от этого не понесет. С расположением нонсенс-мутации в конце гена все еще проще — часто она находится в неважной для функции белка части и поэтому никакой роли не играет. Кстати, это не наше предположение: действительно, с большой частотой нонсенс-мутации предпочитают находиться в конце гена (это показывали во многих работах и на приматах, и на человеке, и мы — на плодовых мушках). Здесь внимательный читатель может возмутиться: «Что значит “предпочитают”? Они же не живые!». Все так, мы говорим «предпочитают», имея в виду, что нонсенс-мутаций там особенно много. Но это говорит лишь о том, что, находясь там, вреда они никакого не приносят, и поэтому не изымаются из популяции под действием отрицательного отбора.
И когда мы, наконец, отфильтровали ненужное и оставили только те гены, в которых нонсенс-мутация точно влияет на белок, то оказалось, что они действительно превратились в псевдогены и эволюционируют нейтрально [12]. Об этом нам рассказало соотношение pN/pS, которое равнялось 1, то есть несинонимичные замены накапливались с той же скоростью, что и синонимичные. Но ведь всегда хочется найти что-то интересное и неожиданное! И хоть у нас было совсем немного генов, мы решились и поделили их на две категории (к слову, больше мы выборку не подразделяли и глобальные выводы по двум–трем генам сделать не пытались). В первой категории были гены, которые состоят из одного экзона. Разумно, что если в них появился преждевременный стоп-кодон, то они должны сразу же превратиться в псевдоген. Так и оказалось. Чуть другой результат мы получили для генов из второй категории — для многоэкзонных генов. Мы заметили, что соотношение pN/pS в них меньше 1. Но в чем может быть причина? Во-первых, они длиннее одноэкзонных, а во-вторых, у них возможен альтернативный сплайсинг, с которым мы познакомились ранее. Поэтому некоторый экзон может присутствовать лишь в части изоформ. И, подсчитав pN/pS отдельно для экзона с нонсенс-мутацией и экзонов без нее, мы получили такой результат, о котором даже не мечтали!
Экзон, в котором появился преждевременный стоп-кодон, полностью ломался и начинал эволюционировать нейтрально. Его такая мутация «обижала», как и в случае одноэкзонного гена. А вот другие экзоны нонсенс-мутация, по-видимому, не затрагивала. Соотношение pN/pS в них было меньше 1, то есть мы наблюдаем воздействие на них отрицательного отбора, а вовсе не нейтральную эволюцию.
Вероятной причиной может быть рекомбинация, которая «разделяет» и «перемешивает» фрагменты генома. Поэтому даже если нонсенс-мутация «поразила» весь ген и другие экзоны тоже начали эволюционировать нейтрально, мы этого не увидим, потому что рекомбинация «принесла» эти самые другие экзоны из геномов мух без нонсенс-мутации. А у них они находятся под отрицательным отбором! Подробнее это изображено на рисунке ниже и разъяснено в подписи к нему.
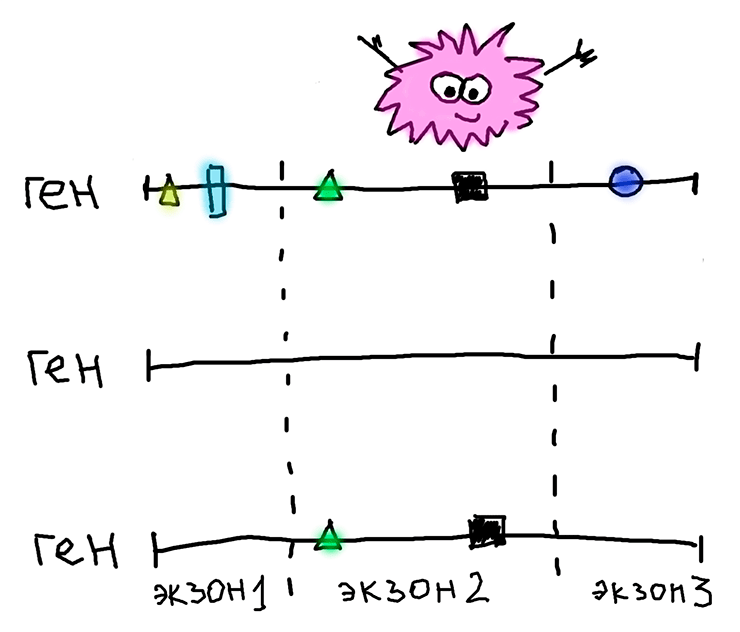
Три варианта одного и того же многоэкзонного гена, принадлежащие трем разным мухам. Первый (вверху) — ген, в котором после появления нонсенс-мутации «сломались» все экзоны, он целиком стал псевдогеном и начал эволюционировать нейтрально (накопившиеся замены изображены геометрическими фигурами). Второй (посередине) — тот же ген, но без нонсенс-мутации, то есть кодирующий нормальный функциональный белок. Никаких замен в нем нет. Третий (нижний) — результат, который получился после рекомбинации между геномами первой и второй мух. Накопившиеся замены остались только в пораженном нонсенс-мутацией экзоне, а экзон 1 и экзон 3 были «принесены» от «здоровой» мухи (средний вариант на рисунке) без такой вредной мутации. Поэтому, из-за рекомбинации, мы и можем наблюдать отрицательный отбор, который на самом деле воздействовал на геном «здоровой» мухи, а у мухи с нонсенс-мутацией его и в помине нет.
Другая сила, которая может быть причастна к такому эффекту, — альтернативный сплайсинг, из-за которого у гена могут быть разные изоформы: и с экзоном с вредной нонсенс-мутацией, и без него. Получается, что другие, не имеющие «сломанного» экзона изоформы, могут спокойно функционировать, и с них получается нормальный работающий белок. И на такие варианты гена будет воздействовать отрицательный отбор.
Здесь внимательный читатель может поинтересоваться, откуда же возникают изоформы с экзонами, расположенными после нонсенс-мутации, если такая мутация, скорее всего, останавливает транскрипцию? Все дело кроется именно в ней. В геномах живых организмов транскрипция может проходить и с кодирующих, и с некодирующих участков ДНК. Она совсем неизбирательна, поэтому нельзя говорить о том, что она остановится именно на нонсенс-мутации. Исходя из этого, ученый мир считает, что такие мутации не влияют на транскрипцию, и полученная в результате пре-мРНК полноценна. Поэтому в результате сплайсинга могут появляться изоформы, включающие в себя экзоны, расположенные после экзона с нонсенс-мутацией.
Кстати, а когда мы взяли данные транскриптомов для плодовых мушек D. melanogaster, то стало видно, что изоформы, включающие в свой состав экзоны с нонсенс-мутациями, редко используются. Что видится логичным, ведь если бы эти вредные нонсенс-мутации были в часто используемых экзонах, то мы скорее всего такого организма не увидели — он просто не смог бы существовать. А в редко используемых вариантах гена нонсенс-мутации, видимо, не так губительны.
Сложно точно сказать, что же именно является причиной того, что мы видим отбор в других экзонах, — рекомбинация или сплайсинг, но результат однозначно интересный!

Таким образом, в своей статье [12] мы свели вместе разрозненные знания об эволюции генов, которые становятся псевдогенами из-за вредной и хитрой нонсенс-мутации. Она и вправду их обижает, но оказалось, что многоэкзонные гены ловко с ней справляются, исключая из использования пораженный экзон. В таких псевдогенах (и баловливых нонсенс-мутациях) скрыто еще немало интересного для изучения, и мы активно продолжаем их исследовать!
21. Клетка как биологическая система  Читать 0 мин.
Читать 0 мин.
 Читать 0 мин.
Читать 0 мин.21.297. Генетический код и его свойства
Ранее мы подчёркивали, что нуклеотиды имеют важную для формирования жизни на Земле особенность ― при наличии в растворе одной полинуклеотидной цепочки спонтанно происходит процесс образования второй (параллельной) цепочки на основании комплементарного соединения родственных нуклеотидов. Одинаковое число нуклеотидов, в обоих цепочках и их химическое родство, является непременным условием для осуществления такого рода реакций. Однако при синтезе белка, когда информация с иРНК реализуется в структуру белка никакой речи о соблюдении принципа комплементарности идти не может. Это связано с тем, что в иРНК, и в синтезированном белке различно не только число мономеров, но и, что особенно важно, отсутствует структурное сходство между ними (с одной стороны нуклеотиды, с другой аминокислоты). Понятно, что в этом случае возникает необходимость создания нового принципа точного перевода информации с полинуклеотида в структуру полипептида. В эволюции такой принцип был создан и в его основу был заложен генетический код.
Генетический код ― это система записи наследственной информации в молекулах нуклеиновых кислот, основанная на определённом чередовании последовательностей нуклеотидов в ДНК или РНК, образующих кодоны, соответствующие аминокислотам в белке.
Генетический код имеет несколько свойств:
Следует отметить, что некоторые авторы предлагают ещё и другие свойства кода, связанные с химическими особенностями входящих в код нуклеотидов или с частотой встречаемости отдельных аминокислот в белках организма и т.д. Однако эти свойство вытекают из вышеперечисленных, поэтому там мы их и рассмотрим.
Генетический код, как и многое сложно организованные система имеет наименьшую структурную и наименьшую функциональную единицу. Триплет ― наименьшая структурная единица генетического кода. Состоит она из трёх нуклеотидов. Кодон ― наименьшая функциональная единица генетического кода. Как правило, кодонами называют триплеты иРНК. В генетическом коде кодон выполняет несколько функций. Во-первых, главная его функция заключается в том, что он кодирует одну аминокислоту. Во-вторых, кодон может не кодировать аминокислоту, но, в этом случае, он выполняет другую функцию (см. далее). Как видно из определения, триплет ― это понятие, которое характеризует элементарную структурную единицу генетического кода (три нуклеотидов). Кодон ― характеризует элементарную смысловую единицу генома ― три нуклеотида определяют присоединение к полипептидной цепочки одной аминокислоты.
Элементарную структурную единицу вначале расшифровали теоретически, а затем её существование подтвердили экспериментально. И действительно, 20 аминокислот невозможно закодировать одним или двумя нуклеотидом т.к. последних всего 4. Три нуклеотида из четырёх дают 43 = 64 варианта, что с избытком перекрывает число имеющихся у живых организмах аминокислот (см.табл. 1).
Представленные в таблице 64 сочетания нуклеотидов имеют две особенности. Во-первых, из 64 вариантов триплетов только 61 являются кодонами и кодируют какую-либо аминокислоту, их называют смысловые кодоны. Три триплета не кодируют.
Таблица 1.
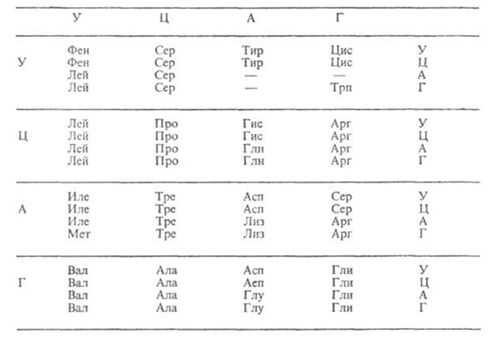
Кодоны информационной РНК и соответствующие им аминокислотыявляются стоп-сигналами, обозначающие конец трансляции. Таких триплетов три ― УАА, УАГ, УГА, их ещё называют «бессмысленные» (нонсенс кодоны). В результате мутации, которая связана с заменой в триплете одного нуклеотида на другой, из смыслового кодона может возникнуть бессмысленный кодон. Такой тип мутации называют нонсенс-мутация. Если такой стоп-сигнал сформировался внутри гена (в его информационной части), то при синтезе белка в этом месте процесс будет постоянно прерываться ― синтезироваться будет только первая (до стоп-сигнала) часть белка. У человека с такой патологией будет ощущаться нехватка белка и возникнут симптомы, связанные с этой нехваткой. Например, такого рода мутация выявлена в гене, кодирующем бета-цепь гемоглобина. Синтезируется укороченная неактивная цепь гемоглобина, которая быстро разрушается. В результате формируется молекула гемоглобина, лишённая бета-цепи. Понятно, что такая молекула вряд ли будет полноценно выполнять свои обязанности. Возникает тяжёлое заболевания, развивающееся по типу гемолитической анемии (бета-ноль талассемия, от греческого слова «Таласа» ― Средиземное море, где эта болезнь впервые обнаружена).
Механизм действия стоп-кодонов отличается от механизма действия смысловых кодонов. Это следует из того, что для всех кодоны, кодирующие аминокислоты, найдены соответствующие тРНК. Для нонсенс-кодонов тРНК не найдены. Следовательно, в процессе остановки синтеза белка тРНК не принимает участие.
Кодон АУГ (у бактерий иногда ГУГ) не только кодируют аминокислоту метионин и валин, но и является инициатором трансляции.
61 из 64 триплетов кодируют 20 аминокислот. Такое трёхразовое превышение числа триплетов над количеством аминокислот позволяет предположить, что в переносе информации могут быть использованы два варианта кодирования. Во-первых, не все 64 кодона могут быть задействованы в кодировании 20 аминокислот, а только 20 и, во-вторых, аминокислоты могут кодироваться несколькими кодонами. Исследования показали, что природа использовала последний вариант.
Код, при котором одна аминокислота кодируется несколькими триплетами, называется вырожденным или избыточным. Почти каждой аминокислоте соответствует несколько кодонов. Так, аминокислота лейцин может кодироваться шестью триплетами — УУА, УУГ, ЦУУ, ЦУЦ, ЦУА, ЦУГ. Валин кодируется четырьмя триплетами, фенилаланин — двумя и только триптофан и метионин кодируются одним кодоном. Свойство, которое связано с записью одной и той же информации разными символами носит название вырожденность.
Число кодонов, предназначенных для одной аминокислоты, хорошо коррелируется с частотой встречаемости аминокислоты в белках. И это, скорее всего, не случайно. Чем больше частота встречаемости аминокислоты в белке, тем чаще представлен кодон этой аминокислоты в геноме, тем выше вероятность его повреждения мутагенными факторами. Поэтому понятно, что мутированный кодон имеет больше шансов кодировать туже аминокислоту при высокой его вырожденности. С этих позиций вырожденность генетического кода является механизмом защищающим геном человека от повреждений.
Необходимо отметить, что термин вырожденность используется в молекулярной генетики и в другом смысле. Так основная часть информации в кодоне приходится на первые два нуклеотида, основание в третьем положении кодона оказывается малосущественным. Этот феномен называют “вырожденностью третьего основания”. Последняя особенность сводит до минимума эффект мутаций. Например, известно, что основной функцией эритроцитов крови является перенос кислорода от легких к тканям и углекислого газа от тканей к легким. Осуществляет эту функцию дыхательный пигмент — гемоглобин, который заполняет всю цитоплазму эритроцита. Состоит он из белковой части ― глобина, который кодируется соответствующим геном. Кроме белка в молекулу гемоглобина входит ген, содержащий железо. Мутации в глобиновых генах приводят к появлению различных вариантов гемоглобинов. Чаще всего мутации связаны с заменой одного нуклеотида на другой и появлением в гене нового кодона, который может кодировать новую аминокислоту в полипептидной цепи гемоглобина. В триплете, в результате мутации может быть заменён любой нуклеотид ― первый, второй или третий.
Известно несколько сотен мутаций, затрагивающих целостность генов глобина. Около 400 из них связаны с заменой единичных нуклеотидов в гене и соответствующей аминокислотной заменой в полипептиде. Из них только 100 замен приводят к нестабильности гемоглобина и различного рода заболеваниям от легких до очень тяжелых. 300 (примерно 64%) мутаций-замен не влияют на функцию гемоглобина и не приводят к патологии. Одной из причин этого является упомянутая выше “вырожденность третьего основания”, когда замена третьего нуклеотида в триплете, кодирующем серин, лейцин, пролин, аргинин и некоторые другие аминокислоты приводит к появлению кодона-синонима, кодирующего ту же аминокислоту. Фенотипически такая мутация не проявится. В отличие от этого любая замена первого или второго нуклеотида в триплете в 100 % случаях приводит к появлению нового варианта гемоглобина. Но и в этом случае тяжёлых фенотипических нарушений может и не быть. Причиной этому является замена аминокислоты в гемоглобине на другую сходную с первой по физико-химическим свойствам. Например, если аминокислота, обладающая гидрофильными свойствами, заменена на другую аминокислоту, но с такими же свойствами.
Гемоглобин состоит из железопорфириновой группы гема (к ней и присоединяются молекулы кислорода и углекислоты) и белка — глобина. Гемоглобин взрослого человека (НвА) содержит две идентичные a-цепи и две b-цепи. Замена в гене, кодирующем b-цепь гемоглобина первого или второго нуклеотида практически всегда приводит к появлению в белка новых аминокислот, нарушению функций гемоглобина и тяжёлым последствия для больного. Например, замена “Ц” в одном из триплетов ЦАУ (гистидин) на “У” — приведет к появлению нового триплета УАУ, кодирующего другую аминокислоту — тирозин. Фенотипически это проявится в тяжёлом заболевании. Аналогичная замена в 63 положении b-цепи полипептида гистидина на тирозин приведет к дестабилизации гемоглобина. Развивается заболевание метгемоглобинемия. Замена, в результате мутации, глутаминовой кислоты на валин в 6-м положении b-цепи является причиной тяжелейшего заболевания — серповидно-клеточной анемии. Не будем продолжать печальный список. Отметим только, что при замене первых двух нуклеотидов может появится аминокислота по физико-химическим свойствам похожая на прежнюю. Так, замена 2-го нуклеотида в одном из триплетов, кодирующего глутаминовую кислоту (ГАА) в b-цепи на “У” приводит к появлению нового триплета (ГУА), кодирующего валин, а замена первого нуклеотида на “А” формирует триплет ААА, кодирующий аминокислоту лизин. Глутаминовая кислота и лизин сходны по физико-химическим свойствам — они обе гидрофильны. Валин — гидрофобная аминокислота. Поэтому, замена гидрофильной глютаминовой кислоты на гидрофобный валин, значительно меняет свойства гемоглобина, что, в конечном итоге, приводит к развитию серповидноклеточной анемии, замена же гидрофильной глютаминовой кислоты на гидрофильный лизин в меньшей степени меняет функцию гемоглобина — у больных возникает легкая форма малокровия. В результате замены третьего основания новый триплет может кодировать туже аминокислоты, что и прежней. Например, если в триплете ЦАУ урацил был заменён на цитозин и возник триплет ЦАЦ, то практически никаких фенотипических изменений у человека выявлено не будет. Это понятно, т.к. оба триплета кодируют одну и туже аминокислоту ― гистидин.
В заключении уместно подчеркнуть, что вырожденность генетического кода и вырожденность третьего основания с общебиологических позиция являются защитными механизмами, которые заложены в эволюции в уникальной структуре ДНК и РНК.
Каждый триплет (кроме бессмысленных) кодирует только одну аминокислоту. Таким образом, в направлении кодон ― аминокислота генетический код однозначен, в направлении аминокислота ― кодон ― неоднозначен (вырожденный).
И в этом случае необходимость однозначности в генетическом коде очевидна. При другом варианте при трансляции одного и того же кодона в белковую цепочку встраивались бы разные аминокислоты и в итоге формировались белков с различной первичной структурой и разной функцией. Метаболизм клетки перешёл бы в режим работы «один ген ― несколько поипептидов». Понятно, что в такой ситуации регулирующая функция генов была бы полностью утрачена.
Считывание информации с ДНК и с иРНК происходит только в одном направлении. Полярность имеет важное значение для определения структур высшего порядка (вторичной, третичной и т.д.). Ранее мы говорили о том, что структуры низшего порядка определяют структуры более высшего порядка. Третичная структура и структуры более высокого порядка у белков, формируются сразу же как только синтезированная цепочка РНК отходит от молекулы ДНК или цепочка полипептида отходит от рибосомы. В то время, когда свободный конец РНК или полипептида приобретает третичную структуру, другой конец цепочки ещё продолжает синтезироваться на ДНК (если транскрибируется РНК) или рибосоме (если транскрибируется полипептид).
Поэтому однонаправленный процесс считывания информации (при синтезе РНК и белка) имеет существенное значение не только для определения последовательности нуклеотидов или аминокислот в синтезируемом веществе, но для жёсткой детерминации вторичной, третичной и т.д. структур.
Код может быть перекрывающимся и не перекрывающимся. У большинства организмов код не перекрывающийся. Перекрывающийся код найден у некоторых фагов.
Сущность не перекрывающего кода заключается в том, что нуклеотид одного кодона не может быть одновременно нуклеотидом другого кодона. Если бы код был перекрывающим, то последовательность из семи нуклеотидов (ГЦУГЦУГ) могла кодировать не две аминокислоты (аланин-аланин) (рис.33, А) как в случае с не перекрывающимся кодом, а три (если общим является один нуклеотид) (рис. 33, Б) или пять (если общими являются два нуклеотида) (см. рис. 33, В). В последних двух случаях мутация любого нуклеотида привела бы к нарушению в последовательности двух, трёх и т.д. аминокислот.
Однако установлено, что мутация одного нуклеотида всегда нарушает включение в полипептид одной аминокислоты. Это существенный довод в пользу того, что код является не перекрывающимся. Неперекрываемость генетического кода связана с ещё одним свойством ― считывание информации начинается с определённой точки ― сигнала инициации. Таким сигналом инициации в иРНК является кодон, кодирующий метионин АУГ. Следует отметить, что у человека всё-таки имеется небольшое число генов, которые отступают от общего правила и перекрываются.
Между кодонами нет знаков препинания. Иными словами триплеты не отделены друг от друга, например, одним ничего не значащим нуклеотидом. Отсутствие в генетической коде «знаков препинания» было доказано в экспериментах.
Код един для всех организмов, живущих на Земле. Прямое доказательство универсальности генетического кода было получено при сравнении последовательностей ДНК с соответствующими белковыми последовательностями. Оказалось, что во всех бактериальных и эукариотических геномах используется одни и те же наборы кодовых значений. Есть и исключения, но их не много.
Первые исключения из универсальности генетического кода были обнаружены в митохондриях некоторых видов животных. Это касалось кодона терминатора УГА, который читался так же как кодон УГГ, кодирующий аминокислоту триптофан. Были найдены и другие более редкие отклонения от универсальности.
Для повторения:
Генетический код ― это система записи наследственной информации в молекулах нуклеиновых кислот, основанная на определённом чередовании последовательностей нуклеотидов в ДНК или РНК, образующих кодоны, соответствующие аминокислотам в белке. Генетический код имеет несколько свойств.
1. Триплетность. Триплет состоит из трёх нуклеотидов. 61 кодон ― смысловые, т.е. кодируют какую-либо аминокислоту, три ― бессмысленные, т.е. не кодируют аминокислоты.
2. Вырожденность или избыточность. Одна аминокислота может кодироваться несколькими кодонами.
3. Однозначность. Один кодон кодирует только одну аминокислоту.
4. Полярность. Считывание информации с ДНК и с иРНК происходит только в одном направлении.
5. Неперекрываемость. Генетический код является не перекрывающимся.
6. Компактность. Между кодонами нет знаков препинания.
7. Универсальность. Код един для всех живущих на земле организмов.



