Окислительное фосфорилирование. Особенности субстратного и коферментного фосфорилирования
Окислительное фосфорилирование – это использование энергии, выделяющейся при окислении органич 939f52gj ских веществ, для синтеза АТФ. Окислительное фосфорилирование бывает 2 типов – субстратное и коферментное.
При субстратном фосфорилировании электроны, отщепленные от данного вещества, затем передаются на это же вещество; однако при этом энергия в молекуле перераспределяется таким образом, что выделяется энергия, идущая на синтез АТФ. Субстратное фосфорилирвание идет в ходе гликолиза; в результате субстратного фосфорилирования выделяется мало энергии.
При коферментном фосфорилировании электрон отщепляется от молекулы органического вещества и передается в цепь переносчиков электронов. В процессе передачи электрона от одного переносчика другому он постепенно теряет энергию, и она используется для синтеза АТФ. В итоге электрон передается на кислород. Коферментное фосфорилирование осуществляется на внутренней мембране митохондрий.
Субстратное фосфорилирование АДФ идет за счет энергии макроэргических связей некоторых соединений (например, метаболитов гликолиза – 1,3-бисфосфоглицерата и фосфоенолпирувата; сукцинил-КоА, креатинфосфата и др.). Этот процесс может происходить как в матриксе митохондрий, так и в цитоплазме клеток независимо от присутствия кислорода. Используется реже, чем окислительное фосфорилирование АДФ.
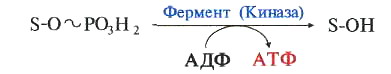
Окислительное фосфорилирование АДФ – превращение АДФ в АТФ происходит с использованием энергии переноса электронов от органических веществ к кислороду. Энергию для окислительного фосфорилирования поставляют окислительно-восстановительные реакции. Процесс может происходить только в аэробных условиях с участием ферментов цепи переноса электронов (ЦПЭ) и АТФ-синтазы.
Окислительное фосфорилирование АДФ – основной механизм синтеза АТФ в организме. Оно происходит в митохондриях, которые являются основными поставщиками АТФ и могут рассматриваться как “энергетические станции” клетки. Митохондрии представляют собой небольшие органеллы овальной формы (рис. 5.3), их количество в клетке может достигать 2000. Митохондрии защищены двумя мембранами: наружной и внутренней. Внутренняя мембрана имеет многочисленные глубокие складки (гребневидные выросты), называемые кристами, которые значительно увеличивают ее поверхность.
Разница между фосфорилированием уровня субстрата и окислительным фосфорилированием
Фосфорилирование на субстратном уровне и окислительное фосфорилирование представляют собой два типа процессов фосфорилирования, которые происходят внутри живых организмов. Фосфорилирование относится

Содержание:
Фосфорилирование на субстратном уровне и окислительное фосфорилирование представляют собой два типа процессов фосфорилирования, которые происходят внутри живых организмов. Фосфорилирование относится к переносу фосфатной группы от одного соединения к другому. Как правило, термин «фосфорилирование» используется для описания образования АТФ. Организмы используют энергию в форме АТФ. У эукариот органеллой, которая продуцирует АТФ, является митохондрий. Но некоторые из АТФ также продуцируются внутри цитоплазмы. главное отличие между уровнем фосфорилирования субстрата и окислительным фосфорилированием является то, что фосфорилирование субстрата на уровне субстрата представляет собой прямое фосфорилирование АДФ с фосфатной группой с использованием энергии, полученной в результате связанной реакции, тогда как окислительное фосфорилирование является продукцией АТФ из окисленных NADH и FADH2.
Ключевые области покрыты
1. Что такое фосфорилирование на уровне субстрата?
— Определение, Процесс, Характеристики
2. Что такое окислительное фосфорилирование
— Определение, Процесс, Характеристики
3. Каковы сходства между фосфорилированием уровня субстрата и окислительным фосфорилированием
— Краткое описание общих черт
4. В чем разница между фосфорилированием уровня субстрата и окислительным фосфорилированием
— Сравнение основных различий
Ключевые слова: аденозиндифосфат (АДФ), аденозинтрифосфат (АТФ), цитоплазма, гликолиз, цикл Кребса, митохондрии, окислительное фосфорилирование, фосфорилирование на субстратном уровне

Что такое фосфорилирование на уровне субстрата?
Фосфорилирование на уровне субстрата относится к типу фосфорилирования, при котором фосфатная группа переносится из субстрата в ADP. Он также может добавлять фосфатную группу к гуанозиндифосфату (GDP) для образования гуанозин трифосфата (GTP). Фосфатная группа напрямую удаляется из субстрата с помощью связанной реакции и переносится в ADP или GDP. Пример реакции субстрата уровня фосфорилирования показан на Рисунок 1.

Рисунок 1: Фосфорилирование уровня субстрата
Фосфорилирование на уровне субстрата происходит в гликолизе и цикле Кребса. Гликолиз является первой стадией как аэробного, так и анаэробного дыхания. При гликолизе происходят две реакции фосфорилирования на уровне субстрата и образуются четыре молекулы АТФ. Фосфоглицераткиназа и пируваткиназа являются двумя ферментами, которые участвуют в фосфорилировании субстрата на уровне гликолиза. Цикл Кребса происходит только при аэробном дыхании. В цикле Кребса фосфорилирование субстрата происходит в митохондриальном матриксе. Две реакции фосфорилирования на уровне субстрата происходят и в цикле Кребса. Фосфоенолпируваткарбоксикиназа и сукцинат КоА лигаза являются двумя ферментами, участвующими в фосфорилировании на уровне субстрата в цикле Кребса. Во время цикла Кребса 2ATP продуцируются путем фосфорилирования на уровне субстрата. Кроме АТФ, 6NADH и 2FADH2s производятся, и их восстановительные потенциалы используются в генерации АТФ путем окислительного фосфорилирования при аэробном дыхании.
Что такое окислительное фосфорилирование
Окислительное фосфорилирование относится к типу фосфорилирования, при котором энергия, выделяющаяся из цепи переноса электронов, используется для генерации АТФ. Он обнаружен во внутренней мембране митохондрий у эукариот. У прокариот окислительное фосфорилирование происходит в плазматической мембране. Высокоэнергетические молекулы, такие как NADH и FADH2 которые образуются в гликолизе, цикле Кребса и цикле жирных кислот, окисляются обратно в цепи переноса электронов. Энергия, выделяемая этими молекулами, используется для генерации АТФ при окислительном фосфорилировании. Окислительное фосфорилирование происходит только при аэробном дыхании. Он производит 26 АТФ на одну молекулу глюкозы. Окислительное фосфорилирование показано на фигура 2.

Рисунок 2: Окислительное фосфорилирование
Ферменты, которые участвуют в окислительном фосфорилировании, представляют собой АТФ-синтазу, цитохромредуктазу, цитохром С-оксидазу и NADH-Q-редуктазу.
Сходства между фосфорилированием на субстратном уровне и окислительным фосфорилированием
Разница между фосфорилированием уровня субстрата и окислительным фосфорилированием
Определение
Уровень субстрата Фосфорилирование: Фосфорилирование на уровне субстрата относится к типу фосфорилирования, при котором фосфатная группа переносится из субстрата в ADP.
Окислительного фосфорилирования: Окислительное фосфорилирование относится к типу фосфорилирования, при котором энергия, выделяющаяся из цепи переноса электронов, используется для генерации АТФ.
Место нахождения
Уровень субстрата Фосфорилирование: Фосфорилирование на уровне субстрата происходит в цитоплазме и митохондриальном матриксе.
Окислительного фосфорилирования: Окислительное фосфорилирование происходит на внутренней мембране митохондрий.
Механизм
Уровень субстрата Фосфорилирование: Фосфатную группу непосредственно удаляют из субстрата с помощью связанной реакции и переносят в ADP.
Окислительного фосфорилирования: Фосфатные группы добавляются из энергии, выделяющейся в цепи переноса электронов.
корреляция
Уровень субстрата Фосфорилирование: Фосфорилирование субстрата является прямым фосфорилированием.
Окислительного фосфорилирования: Окислительное фосфорилирование является косвенным фосфорилированием.
Вхождение
Уровень субстрата Фосфорилирование: Фосфорилирование на уровне субстрата происходит в гликолизе и цикле Кребса.
Окислительного фосфорилирования: Окислительное фосфорилирование происходит в цепи переноса электронов.
Окисление / восстановление коферментов
Уровень субстрата Фосфорилирование: NAD и FAD снижаются во время фосфорилирования уровня субстрата.
Окислительного фосфорилирования: NADH + и FADH + окисляются при окислительном фосфорилировании.
Чистое производство АТФ
Уровень субстрата Фосфорилирование: Четыре АТФ продуцируются во время фосфорилирования на уровне субстрата.
Окислительного фосфорилирования: Тридцать четыре АТФ продуцируются во время окислительного фосфорилирования.
Окислительно-восстановительный потенциал
Уровень субстрата Фосфорилирование: Изменение окислительно-восстановительного потенциала субстрата меньше при уровне фосфорилирования субстрата.
Окислительного фосфорилирования: Изменение окислительно-восстановительного потенциала субстрата происходит больше при окислительном фосфорилировании.
Уровень окисления
Уровень субстрата Фосфорилирование: Частичное окисление субстрата происходит на уровне фосфорилирования субстрата.
Окислительного фосфорилирования: Полное окисление доноров электронов происходит при окислительном фосфорилировании.
Заключение
Фосфорилирование на субстратном уровне и окислительное фосфорилирование являются двумя способами генерирования АТФ внутри живых организмов. АТФ является основной формой молекул энергии, используемых в клеточных механизмах. Фосфорилирование на уровне субстрата происходит в гликолизе и цикле Кребса. Окислительное фосфорилирование происходит в цепи переноса электронов. Фосфорилирование на уровне субстрата представляет собой прямой тип фосфорилирования, при котором фосфатная группа непосредственно переносится в молекулу АДФ. Окислительное фосфорилирование является косвенным методом фосфорилирования, при котором энергия, высвобождаемая в цепи переноса электронов, используется для генерации АТФ. Основное различие между уровнем фосфорилирования субстрата и окислительным фосфорилированием заключается в механизмах продукции АТФ.
Ссылка:
1. «Фосфорилирование на субстратном уровне». Википедия, Фонд Викимедиа, 6 октября 2017 г.,
Цикл Кребса. Биологическое значение




![]()
![]()
Макроэрги. Окислительное и субстратное фосфорилирование.
Связующим звеном между клеточными реакциями, идущими с выделением и потреблением энергии, служит аденозинтрифосфат (АТФ) (рис. 1). При катаболизме часть свободной энергии используется для синтеза АТФ из аденозиндифосфата (АДФ) и неорганического фосфата (Рi, Фн). Молекула АТФ содержит две высокоэнергетические (макроэргические) связи, которые обозначают символом « » (тильда).
 |
Рис. 1. Схема строения аденозинтрифосфата (АТФ).
Гидролиз АТФ, в зависимости от потребностей конкретной реакции в энергии, может происходить двумя способами:
Суммарная реакция: АТФ + Н2О АМФ + 2 Фн – 14,6 ккал/моль (59 кДж/моль).
В клетках кроме АТФ присутствуют и другие высокоэнергетические соединения, которые содержат макроэргические связи, при гидролизе которых выделяется не менее 7 ккал/моль (30 кДж/моль) стандартной свободной энергии (таблица 1).
Таблица 1. Стандартная свободная энергия гидролиза некоторых макроэргических соединений (макроэргов)
Способы синтеза АТФ в клетке:
1) Субстратное фосфорилирование – синтез АТФ из АДФ и фосфорной кислоты с использованием энергии высокоэнергетического субстрата. Этот способ синтеза АТФ не требует присутствия кислорода, т.е. происходит в анаэробных условиях.
Реакции субстратного фосфорилирования происходят:
3) в одной реакции ЦТК, с использованием макроэргического субстрата – сукцинил-КоА.
2) Окислительное фосфорилирование – синтез АТФ из АДФ и фосфорной кислоты с использованием энергии окисления водорода в дыхательной цепи. Этот способ синтеза АТФ требует присутствия кислорода, т.е. происходит в аэробных условиях. Дыхательная цепь локализуется во внутренней мембране митохондрий.
Биологическое окисление. Основные положения теорий теории А.М. Баха и В.И. Палладина. Современные представления о биологическом окислении.
Функции биологического окисления:
I. Энергетическое обеспечение: поддержание температуры тела, химических синтезов, осмотических явлений, электрических процессов, механической работы.
II. Синтез важнейших метаболитов.
III. Регуляция обмена веществ.
IV. Обезвреживание ксенобиотиков (чужеродных веществ).
V. Устранение вредных для клетки продуктов обмена.
История развития представлений о механизмах биологического окисления.
Перекисная теория А.Н. Баха. В 1896-1897 гг. русский ученый А.Н. Бах сформулировал теорию медленного окисления, согласно которой молекулярный кислород активируется с помощью ненасыщенной органической молекулы, при этом образуется пероксид этого соединения. Возникшие пероксиды органических соединений, как и пероксид водорода, могут окислять другие вещества при каталитическом воздействии пероксидазы.
1-ая фаза: SH2 + R↔ S+ RH2 2-ая фаза: RH2 + ½ O2 → H2O + R

В общем виде окислительно-восстановительную реакцию можно написать:
Способы передачи электронов от одной молекулы к другой:
I. Прямой перенос электронов.
Например, окислительно-восстановительная пара Fe 2+ и Fe 3+ :
III. Перенос электронов путем прямого взаимодействия органического восстановителя с кислородом. В результате образуется продукт, в котором содержится ковалентно связанный кислород. Например, введение в состав органической молекулы атома кислорода с образованием гидроксильной группы:
R–CH3 + ½ О2 ↔ R–CH2–OH. В этой реакции донором электронов является органическая молекула, а атом кислорода играет роль акцептора.
Выделяют два типа биологического окисления:
I. Свободное окисление, при котором свободная энергия, высвобождающаяся при окислении органических молекул, переходит в тепловую энергию и рассеивается. К таким реакциям относятся реакции: микросомального окисления, генерации активных форм кислорода и антиоксидантной защиты.
II. Окисление, сопряженное с фосфорилированием АДФ и синтезом АТФ. Этот тип биологического окисления осуществляется в электронтранспортной цепи (дыхательной цепи), локализованной во внутренней мембране митохондрий или сопряжен с фосфорилированием АДФ на уровне субстрата.
Цикл трикарбоновых кислот (ЦТК), цикл Кребса или цикл лимонной кислоты является общим этапом катаболизма углеводов, жиров и белков. Сложные молекулы органических веществ превращаются в один общий метаболит – активную форму уксусной кислоты:
ацетил-коферментА: СН3 – С S-КоА.
Ацетил-КоА окисляется в ЦТК, который локализуются в матриксе митохондрий. ЦТК включает ряд последовательных реакций:
1. Конденсация ацетил-КоА с оксалоацетатом (щавелевоуксусной кислотой) с образованием цитрата (лимонной кислоты) фермент – цитратсинтаза:
Ацетил-КоА + оксалоацетат + Н2О → цитрат + HS-KoA
Параграф 23 Синтез АТФ субстратное и окислительное
Автор текста – Анисимова Елена Сергеевна.
Авторские права защищены. Продавать текст нельзя.
Курсив не зубрить.
Замечания можно присылать по почте: exam_bch@mail.ru
https://vk.com/bch_5
см. также 22, 20 и 21, 25.
Сокращения:
СФ – субстратное фосфорилирование,
ОФ – окислительное фосфорилирование,
ММП – межмембранное пространство митохондрий.
Главное.
СФ и ОФ – это способы синтеза АТФ.
23.1. Для чего нужен синтез АТФ клеткам.
При использовании АТФ в качестве источника энергии
АТФ превращается в АДФ и фосфат (Н3РО4).
(Иногда просто в АДФ – см. киназные реакции;
даже если АТФ расщепляется на АМФ и дифосфат,
то далее АМФ превращается в АДФ
за счет получения фосфата от АТФ: АМФ + АТФ ; 2 АДФ).
Чтобы [АТФ] в клетке поддерживалась на необходимом уровне,
нужно постоянно превращать АДФ в АТФ.
Иначе все молекулы АТФ превратятся в АДФ,
а без АТФ клетка умирает (гипоэнергетический шок).
Примеры ситуаций, когда клетки и организм в целом
умирают от дефицита АТФ:
Если бы в этих ситуациях клетки не теряли бы возможность синтезировать АТФ,
то эти ситуации не приводили бы к смерти.
23. 2. Фосфорилирование АДФ – синтез АТФ.
Для синтеза АТФ из АДФ
нужно к АДФ присоединить фосфат,
то есть фосфорилировать АДФ.
Поэтому в названии синтеза АТФ из АДФ есть слово «фосфорилирование».
Фосфорилированию могут подвергаться разные вещества,
но когда говорят о СФ или ОФ,
то имеют в виду только фосфорилирование АДФ,
то есть синтез АТФ.
Как синтезируется АДФ, здесь не разбирается.
См. вопрос о синтезе пуриновых нуклеотидов в п.72.
Нужны Р-5-Ф из ПФП и некоторые аминокислоты в качестве сырья для синтеза АДФ.
23.3. Чем отличаются СФ и ОФ.
СФ и ОФ отличаются по источникам энергии,
которая тратится на образование макроэргической связи
между присоединяемым фосфатом и АДФ.
При СФ источником энергии для фосфорилирования АДФ
является расщепление макроэргической связи другого вещества,
а при ОФ источником энергии для синтеза АТФ
является перенос электронов в дыхательной цепи.
Хоть и не напрямую, как будет показано далее.
Процесс назвали ОФ, потому что
при переносе электронов происходит окисление тех веществ,
от которых отщеплен электрон.
23.4. Источники энергии для ОФ – прямой и непрямой.
Непосредственным источником энергии для ОФ
является не перенос электронов,
а пассивный транспорт протонов через канал АТФ-синтАзы,
при котором тратится энергия ЭХП протонов – далее об этом подробнее.
Из-за того, что источником энергии для ОФ является ДЦ,
которая требует кислорода в качестве субстрата,
ОФ протекает только в присутствии кислорода
(то есть только в аэробных условиях).
Это значит, что в анаэробных условиях клетка не может получать энергию способом ОФ.
Поскольку СФ дает намного меньше АТФ, чем ОФ
(от одной молекулы глюкозы:
2 АТФ при СФ вместо 32 АТФ при СФ вместе с ОФ),
то в анаэробных условиях возникает острый дефицит АТФ (и угроза гибели)
в тех клетках, которые тратят много АТФ:
— в головном мозге,
— в коре почек,
— в миокарде и в других аэробных тканях.
Клетки анаэробных тканей могут существовать без ОФ
намного дольше, чем клетки аэробных тканей:
жгут на конечность допускается держать 1,5 часа,
но и они нуждаются в «подключении» ОФ
(за счет восстановления кровотока)
после того как просуществовали 1,5 часа в анаэробных условиях (со жгутом).
Только эритроциты могут существовать без ОФ,
хотя в них и много кислорода.
(в ЭЦ ОФ не происходит, т.к. нет митохондрий).
Тут надо добавить таблицу
«О т л и ч и я способов синтеза АТФ
(из АДФ и фосфата;
= присоединения фосфата к АДФ
= фосфорилирования АДФ)»
Но пока она в отдельном файле.
23.5. Субстратное фосфорилирование.
Вещество, расщепление макроэргической связи которого дает энергию для синтеза АТФ,
называется первичным макроэргом.
АТФ при СФ называется вторичным макроэргом.
Образование
первичного макроэрга происходит
за счет энергии, которая выделяется при окислении другого вещества,
которое называется субстратом СФ
(то есть не просто субстратом,
а именно субстратом для процесса СФ);
обычно первичный макроэрг образуется из субстрата СФ – см. далее примеры.
23.5.1. Механизм СФ на примере с сукциниломКоА из ЦТК.
В ЦТК кетоглутарат превращается в сукцинилКоА,
при этом происходит окислительное декарбоксилирование кетоглутарата.
При окислении кетоглутарата выделяется энергия,
за счет которой образуется макроэргическая связь
между сукцинилом и коферментом А (КоА).
При расщеплении МС сукцинилаКоА выделяется энергия,
за счет которой образуется
между ГДФ и фосфатом,
в результате чего образуется ГТФ.
ГДФ и ГТФ – это вещества, аналогичные АДФ и АТФ,
и синтез ГТФ происходит путем фосфорилирования ГДФ
(в данном случае – способом СФ).
Затем ГТФ вступает в реакцию с АДФ
и отдает ему фосфат,
в результате чего АДФ превращается в АТФ, а ГТФ – в ГДФ.
23.5.2. Механизм СФ на 2-х примерах реакций гликолиза.
См. реакции и формулы в п.32.
1-й пример СФ из гликолиза.
В одной из реакций гликолиза
ФГА превращается в 1,3-бис/фосфо/глицерат,
при этом происходит окисление ФГА
и присоединение фосфата макроэргической связью.
При окислении ФГА
(точнее, промежуточного метаболита, связанного с ферментом)
выделяется энергия,
за счет которой образуется
с присоединяемым по 1-му положению фосфатом.
1,3-бис/фосфо/глицерата
выделяется энергия,
за счет которой образуется
между АДФ и фосфатом
(переносимым от 1,3-бис/фосфо/глицерата),
в результате чего образуется АТФ (вторичный макроэрг).
1-й пример СФ из гликолиза.
В другой реакции гликолиза
2-ФГ превращается в фосфо/енол/пируват (ФЕП),
при этом происходит отщепление воды,
а связь с фосфатом становится макроэргической.
фосфо/енол/пирувата
выделяется энергия,
за счет которой образуется
между АДФ и фосфатом (переносимым от ФЕПа),
в результате чего образуется АТФ.
Между СФ в ЦТК и СФ в гликолизе есть важное отличие:
СФ в ЦТК, как и весь ЦТК,
не происходит в отсутствие кислорода и вне митохондрий
(то есть в анаэробных условиях;
МХ нет в ЭЦ;
ЦТК не работает в анаэробных условиях,
потому что должен превращать НАД+ в НАДН для ДЦ,
но без кислорода ДЦ не работает,
не потребляет НАДН и не превращает НАДН в НАД+).
А СФ в гликолизе может происходить в отсутствие кислорода
и без митохондрий.
Поэтому СФ в гликолизе –
единственный источник АТФ для клеток,
когда в них нет кислорода :
при ишемии, гипоксии,
из-за жгута, спазма, низкого артериального давления и т.д..
Или если в клетках нет митохондрий, как в эритроцитах.
Тут надо добавить таблицу
«П р и м е р ы субстратного фосфорилирования.»
Но пока она в отдельном файле.
Уже говорилось, что при ОФ
АТФ синтезируется за счет энергии дыхательной цепи.
Поэтому говорят, что работа ДЦ сопряжена (связана) с синтезом АТФ
(с фосфорилированием АДФ).
Существует теория, которая описывает механизм ОФ
(то есть механизм сопряжения окисления в ДЦ и фосфорилирования АДФ).
Автор теории – Митчелл.
Названия теории ОФ – хемоосмотическая, протондвижущая.
Согласно теории Митчелла (ТМ),
энергия, выделяющаяся при переносе электронов по ДЦ,
не сразу тратится на синтез АТФ.
Сначала энергия ДЦ тратится на транспорт протонов (Н+)
из матрикса МХ в межмембранное пространство
через внутреннюю мембрану МХ
(против градиента протонов).
Затем протоны возвращаются в матрикс МХ
через канал фермента АТФ-синтазы,
и АТФ-синтаза синтезирует АТФ
за счет энергии, которая выделяется при транспорте протонов
(т.к. транспорт протонов в матрикс происходит по градиенту протонов,
пассивный, энергия не тратится, а выделяется).
Транспорт протонов через мембрану происходит с помощью белков-транспортеров. (Потому что липидный слой мембраны не проницаем для ионов.)
23.6.1. О транспорте протонов из матрикса
(в межмембранное пространство,
через внутреннюю мембрану МХ
с внутренней стороны внутренней мембраны
на внешнюю сторону внутренней мембраны;
но из митохондрий протоны при этом не выходят,
они оказываются между внутренней мембраной и внешней мембраной,
то есть в межмембранном пространстве,
ближе к внутренней мембране, «на» ее внешней стороне).
Предполагается, что транспорт протонов из матрикса
осуществляется с участием белков ДЦ и кофермента Q.
При транспорте протонов из матрикса
происходит их накопление в ММП,
с внешней стороны мембраны оказывается больше протонов,
чем с внутренней стороны мембраны,
то есть возникает разница концентраций протонов
(градиент протонов)
с разных сторон мембраны.
Транспорт протонов из матрикса
происходит из области меньшей [Н+] в область большей [Н+],
то есть против градиента протонов
(«оттуда, где протонов меньше,
туда, где протонов больше»).
Транспорт частиц против градиента
требует затрат энергии
(и поэтому называется активным).
Источником энергии для транспорта протонов из матрикса
является энергия, выделяющаяся при работе дыхательной цепи
(то есть при переносе электронов по ДЦ).
23.6.2. О транспорте протонов в матрикс.
(из межмембранного пространства,
через внутреннюю мембрану МХ
с внешней стороны внутренней мембраны
на внутреннюю сторону внутренней мембраны;
протоны оказываются в матриксе ближе к внутренней мембране,
«на» ее внутренней стороне).
Транспорт протонов в матрикс
происходит, в основном (о других вариантах позже)
через канал АТФ-синтазы.
При транспорте протонов в матрикс
[Н+] в матриксе увеличивается,
а в ММП [Н+] уменьшается,
в результате чего снижается разница концентраций протонов
с разных сторон мембраны.
Транспорт протонов в матрикс
происходит из области большей [Н+] в ММП
в область меньшей [Н+] в матриксе,
то есть по градиенту протонов
(«оттуда, где протонов больше, туда, где протонов меньше»).
Транспорт частиц по градиенту
НЕ требует затрат энергии
(и поэтому называется пассивным).
(Точнее, источником энергии является сам градиент, его снижение).
Более того, при транспорте частиц по градиенту
энергия выделяется
и может использоваться для совершения работы
(то есть процессов, требующих затрат энергии).
При транспорте протонов в матрикс энергия тоже выделяется.
Поскольку при транспорте протонов в матрикс
(то есть при возвращении протонов в матрикс)
протоны проходят через канал АТФ-синтазы,
то выделяющаяся энергия «достается» АТФ-синтазе.
АТФ-синтаза тратит энергию на синтез АТФ
(поэтому и называется АТФ-синтазой).
Как говорилось выше, способ синтеза АТФ при этом называется ОФ.
Работа, которая совершается за счет энергии,
выделяющейся при транспорте протонов в матрикс –
это синтез АТФ
(эта работа называется химической,
потому что заключается в обеспечении энергией химической реакции).
23.7. На что тратится энергия, кроме синтеза АТФ.
АТФ-синтаза использует на синтез АТФ
не 100% энергии, выделяющейся при транспорте протонов,
а около 40% энергии
(при разобщении ещё меньше – см. далее).
Остальная часть энергии (60%)
рассеивается в виде тепла
(это тепло называется первичным – см. далее)
и тратится на транспорты веществ через мембрану.
То есть на осмотическую работу.
23.8. Градиент протонов – форма конвертируемой энергии.
ЭХП протонов.
Поскольку пассивный транспорт протонов
может быть источником энергии для совершения работы,
то градиент протонов,
при наличии которого возможет пассивный транспорт протонов,
является формой конвертируемой энергии.
Эта форма энергии называется электро-химическим потенциалом (ЭХП) протонов
и обозначается ;µН+.
(Знак ; (дельта) означает, что есть разная концентрация;
µ означает, что это форма энергии).
Энергия, заключенная в ;µН+,
выделяется
и может быть использована при пассивном транспорте протонов,
то есть при транспорте протонов в область меньшей концентрации:
при этом разница концентраций протонов уменьшается
(градиент протонов уменьшается),
;µН+ уменьшается.
;µН+ возникает при появлении разных концентраций протонов
с разных сторонах внутренней мембраны митохондрий
(то есть при появлении градиента протонов).
И увеличивается при увеличении разницы [H+].
23.8.1. Как создать ЭХП протонов.
Для увеличения разницы [H+] (и создания ;µН+)
нужно транспортировать протоны
из области меньшей концентрации
в область большей концентрации
(то есть против градиента протонов),
то есть должен произойти активный транспорт протонов.
Для активного транспорта нужны затраты энергии.
Энергию для активного транспорта протонов
(и тем самым – для создания ;µН+) дает ДЦ.
23.8.2. Два компонента ЭХП протонов.
;µН+ называется ЭХП,
потому что в ;µН+ есть два компонента (и они отражены в названии): электрический и химический.
Электрический компонент ЭХП обусловлен тем, что
(при наличии на мембране градиента ионов)
с разных сторон мембраны
разные концентрации заряженных частиц
и в результате – разные заряды.
Электрический компонент ЭХП обозначается ;; («пси», ;;).
Поскольку при наличии ;µН+ протонов
(у которых есть положительный заряд)
больше на внешней стороне внутренней мембраны МХ,
чем на внутренней,
то внешняя сторона заряжена более положительно,
чем внутренняя
(а внутренняя – более отрицательно, чем внешняя).
Другое название электрического компонента ЭХП – мембранный потенциал.
Химический компонент ЭХП
обусловлен тем, что
(при наличии на мембране градиента ионов)
с разных сторон мембраны
есть разные концентрации химического вещества.
Химический компонент ЭХП обозначается ;; («фи»).
Поскольку протоны создают кислую среду
и протонов при наличии ;µН+ больше
на внешней стороне внутренней мембраны МХ
(чем на внутренней стороне),
то внешняя сторона имеет более кислую реакцию
(и меньший рН),
чем внутренняя сторона,
а внутренняя – более менее кислую реакцию,
чем внешняя сторона,
и больший рН.
Другое название химического компонента ЭХП –
концентрационный градиент.
Тут надо добавить таблицу
Но пока она в отдельном файле.
«Транспорт Н+ через внутреннюю мембрану митохондрий из матрикса и обратно».
(К теории Митчелла).
Протоны создают кислую среду:
где больше протонов, там кислее среда и меньше рН.
Н+ имеет положительный заряд,
поэтому – чем больше протонов на данной стороне мембраны,
тем более положительно заряжена эта сторона мембраны.
Н+ больше в ММП (при работе ДЦ),
то есть на внешней стороне внутренней мембраны
(со стороны ММП).
На внешней стороне внутренней мембраны
больше положительно заряженных частиц.
Внешняя сторона внутренней мембраны МХ заряжена более положительно.
23.9. Сопряжение и разобщение
окисления и фосфорилирования
(окислительного фосфорилирования).
(Имеется в виду – окисления в ДЦ и фосфорилирования АДФ).
Когда энергия ДЦ используется для синтеза АТФ,
то говорят, что процесс ДЦ и процесс синтеза АТФ сопряжены
(окисление и фосфорилирование сопряжены).
При этом способ синтеза АТФ называется окислительным фосфорилированием.
Сопряжение имеет место тогда,
когда Н+, транспортированные из матрикса в ММП
за счет энергии ДЦ,
возвращаются из ММП обратно в матрикс через АТФ-синтазу.
23.10. Разобщение ОФ.
Но если протоны возвращаются в матрикс
НЕ через АТФ-синтазу, а другим способом,
то энергия, выделяющаяся при транспорте протонов в матрикс,
не «достается» АТФ-синтазе,
и она не синтезируется АТФ.
Это явление называется разобщением ДЦ и синтеза АТФ
или разобщением окислительного фосфорилирования.
При разобщении энергия возвращения протонов в матрикс
просто рассеивается в виде тепла,
что приводит к увеличению температуры тела.
Причины разобщения ОФ:
к разобщению приводят различные факторы,
которые позволяют протонам не проходить через АТФ-синтазу.
Протоны могут не проходить через АТФ-синтазу
(одна из причин разобщения),
если есть вещества,
способные присоединять к себе протоны в ММП,
проходить с протонами через мембрану в матрикс,
а в матриксе «освобождать» протоны,
присоединенные к себе в ММП.
Пример – динитрофенол (применялся в промышлености)
и (возможно) иодтиронины.
Протоны могут не проходить через АТФ-синтазу,
если в мембране есть другой белок,
способный пропускать протоны
(протонный канал) –
примером такого белка является термогенин,
который есть в митохондриях бурой жировой ткани.
Разные факторы, которые приводят к разобщению
(являются причинами разобщения)
называются разобщителями.
Разобщителями являются жирные кислоты в буром жире
(способствуют работе термогенина),
некоторые микробные токсины
(дифтерийный).
Из всех названных разобщителей только жирные кислоты (в буром жире)
относят к физиологическим –
то есть разобщение ими не вредно для организма.
Остальные разобщители считаются патологическими.
(Об ЙТ далее).
Тут надо добавить таблицу.
Но пока она в отдельном файле.
«Сравнение сопряжения и разобщения ДЦ и синтеза АТФ
(путем окислительного фосфорилирования фосфорилирования)».
23.11. С р а в н е н и е блокаторов и разобщителей.
Ещё раз про блокаторы ДЦ.
Нужно не путать действие блокаторов ДЦ и разобщителей ОФ.
Блокаторы прекращают работу ДЦ,
поэтому энергия ни запасается в виде АТФ (слабость),
ни рассеивается в виде тепла (озноб).
А разобщители не прекращают работу ДЦ
и даже (ЙТ) могут усиливать работу ДЦ.
Тут надо добавить таблицу.
Но пока она в отдельном файле.
«С р а в н е н и е блокаторов и разобщителей».
23.12. Действие ЙТ на работу ДЦ и на разобщение (п.104) –
ЙТ активируют ДЦ
и при этом разобщают ОФ.
При небольшом избытке ЙТ (при начальном гипертиреозе)
соотношение эффектов активации ДЦ и разобщения ОФ таково,
что АТФ и тепла больше,
чем при нормальном количестве ЙТ
(это увеличивает энергичность и устойчивость к холоду).
Но при значительном избытке ЙТ (при запущенном гипертиреозе)
разобщение настолько сильное,
что количество АТФ становится меньше,
чем в норме
(при том, что ДЦ активнее, чем в норме).
При снижении количества ЙТ (при гипотиреозе) ДЦ менее активна,
энергии дает меньше,
при этом снижается количество и АТФ, и тепла.
Тут надо добавить таблицу.
Но пока она в отдельном файле.
«Окислительное фосфорилирование при ; и ; функции ЩВЖ».
Тут надо добавить таблицу.
Но пока она в отдельном файле.
«Р а з о б щ е н и е окисления и фосфорилирования»:
то есть окисления в ДЦ и фосфорилирования АДФ,
приводящего к синтезу АТФ способом ОФ.



