Что такое перинуклеарное пространство
• Ядро окружено оболочкой, состоящей из двух мембран
• В ядерной оболочке присутствуют многочисленные ЯПК, которые представляют собой единственные каналы обмена молекулами и макромолекулами между ядром и цитоплазмой
Наружная ядерная мембрана переходит в мембраны ЭПР и, так же как большая часть его мембран, покрыта рибосомами, принимающими участие в синтезе белка. На рисунке ниже показана связь наружной мембраны с ЭПР.
Пространство между наружной и внутренней ядерными мембранами представляет собой перинуклеарное пространство (ПП). Так же как наружная мембрана связана с мембраной ЭПР, ПП ядерной оболочки контактирует с внутренним пространством ЭПР. Толщина каждой из двух мембран составляет 7-8 нм (нм), а ширина ПП ядерной оболочки — 20-40 нм.
При исследовании препаратов ядерной оболочки в электронном микроскопе, наиболее заметной особенностью структуры являются ЯПК (ядерные поровые комплексы), которые служат каналами транспортировки большинства молекул между ядром и цитоплазмой. Оболочка ядер большинства клеток содержит около 10-20 ЯПК на квадратный микрон поверхности. Так, клетки дрожжей содержат 150-250 ЯПК, а соматические клетки млекопитающих 2000-4000.
Однако некоторые клетки обладают гораздо большей плотностью пор, вероятно, потому, что для них характерна высокая интенсивность процессов транскрипции и трансляции, что предполагает транспорт большого количества макромолекул в ядро и из него. Например, поверхность ядра ооцитов амфибий почти полностью покрыта ЯПК.
Наиболее убедительное доказательство в пользу эндосимбиотического происхождения митохондрий и хлоропластов заключается в том, что рибосомы обеих органелл больше напоминают рибосомы современных прокариот, и в меньшей степени эти же микроструктуры цитоплазмы эукариотической клетки. Гораздо менее ясным представляется происхождение ядра. Однако существование двойной ядерной мембраны, подобно мембране митохондрий и хлоропластов, позволяет предполагать, что захваченная прокариотическая клетка превратилась в ядро, содержащее всю клеточную ДНК.
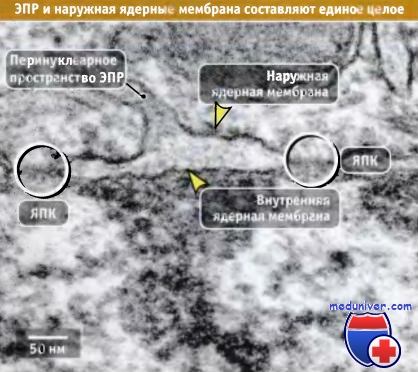 Ядерная оболочка соединена с эндоплазматическим ретикулумом (ЭПР).
Ядерная оболочка соединена с эндоплазматическим ретикулумом (ЭПР).  Поверхность ядерной мембраны ооцита Xenopus laevis покрыта комплексами ядерных пор.
Поверхность ядерной мембраны ооцита Xenopus laevis покрыта комплексами ядерных пор. 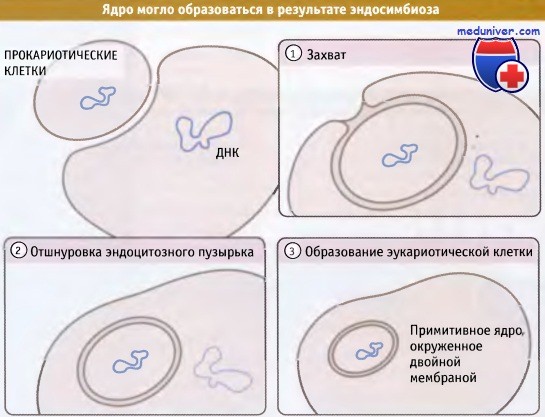 Ядро могло образоваться в результате эндосимбиоза, процесса,
Ядро могло образоваться в результате эндосимбиоза, процесса,
при котором одна прокариотическая клетка захватывает другую клетку; затем захваченная клетка становится примитивным ядром.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Что такое перинуклеарное пространство
Внутренняя среда клетки — цитоплазма — сложно организованная система, включающая ядро, мембранные и немембранные органеллы, включения, которые находятся во взвешенном состоянии в гиалоплазме. Последняя представляет собой гель с изменяющейся в зависимости от функционального состояния клетки степенью вязкости.
В составе гиалоплазмы находятся структурные и ферментные белки клетки, различные метаболиты, ионы. Здесь присутствуют ферменты, участвующие в синтезе аминокислот, нуклеотидов, жирных кислот, биосинтезе Сахаров. В гиалоплазме происходят процессы гликолиза и синтез части АТФ, модификация ферментов (например, фосфорилирование), приводящая к их активации, либо инактивации. В гиалоплазме начинается ряд биосинтетических процессов, которые в дальнейшем продолжаются в той или иной внутриклеточной системе.
В электронном микроскопе гиалоплазма выглядит гомогенной и характеризуется низкой электронной плотностью. Мегавольтная электронная микроскопия обнаруживает в ней микротрабекулярную сеть, состоящую из тончайших фибрилл, пересекающих гиалоплазму в различных направлениях. В ячейках этой сети располагаются органеллы, а в ее «узлах» фиксированы полисомы. Микротрабекулярная сеть гиалоплазмы образует связи с микротрубочками и микрофиламентами опорно-двигательной системы клетки и совместно с этими элементами участвует в перемещении и функционировании внутриклеточных структур.
Органеллы — это обязательно присутствующие в клетке ультрамикроскопические структуры, представляющие собой отграниченные от окружающей гиалоплазмы биологической мембраной отсеки, или компартменты (мембранные органеллы), и расположенные в гиалоплазме рибонуклеопротеиды, биополимеры и их комплексы (немембранные органеллы).
С точки зрения цитофизиологии целесообразно рассмотреть строение и функции внутриклеточных структур в связи с их участием в метаболических процессах, таких как хранение и передача поколениям генетической информации, синтез и транспорт веществ, энергообеспечение метаболических процессов и др.
Ядро — важнейший компонент, с деятельностью которого связаны хранение генетической информации, размножение клеток и передача генетического материала поколениям, участие в синтезе белков.
В неделящихся клетках (интерфазе клеточного цикла) ядро хранит закодированную в ДНК хромосом информацию о белковом синтезе и обеспечивает синтез тех белковых молекул, которые необходимы клетке в процессе ее роста, дифференцировки и физиологической регенерации; в этот период в ядре синтезируются участвующие в образовании белка рибосомальная, информационная и трансферная РНК, формируются субъединицы рибосом. При подготовке клетки к делению ядро удваивает генетическую информацию о белковом синтезе, создавая ее точную копию для передачи дочерним клеткам.
В интерфазе в составе ядра обнаруживаются оболочка (нуклеолемма), хроматин, нуклеоплазма, ядрышко.
Ядерная оболочка представляет собой часть внутриклеточной мембранной системы (совместно с гранулярной и агранулярной эндоплазматической сетью). Она состоит из внутреннего и наружного листков, между которыми находится щелевид-ное пространство — перинуклеарное, ширина которого варьирует в зависимости от функциональной активности клетки. Наружный листок нуклеолеммы со стороны гиалоплазмы окружен сетью виментиновых промежуточных филаментов и имеет на своей поверхности свободные рибосомы, прикрепленные к ней большими субъединицами. Внутренний листок — гладкий, не содержит рибосом, образует связи с пластинкой (ламиной) ядра и участвует в фиксации интерфазных хромосом. Листки нуклеолеммы выполняют по отношению к ядру две важные функции — формообразовательную и рецепторно-барьерно-транспортную.
— Вернуться в оглавление раздела «гистология»
Что такое перинуклеарное пространство
• Ядерные поровые комплексы (ЯПК) представляют собой симметричные структуры, расположенные в местах слияния внешней и внутренней ядерной мембран
• В клетках человека каждый ЯПК обладает массой около 120 х 10 6 Да, что в 40 раз больше массы рибосомы, и состоит из множества копий молекул, включающих 30 белков
• ЯПК содержит филаменты, простирающиеся в цитоплазму, и напоминающие корзину структуры, проникающие в ядро
Ядерный поровой комплекс (ЯПК) ядерной мембраны представляют собой единственные каналы, которые связывают ядро и цитоплазму. В клетках человека ЯПК имеют молекулярную массу около 120 х 106 Да и наружный диаметр порядка 120 нм. Общая масса ЯПК в 40 раз превышает массу эукариотической рибосомы. Комплекс ядерной поры состоит из многих копий примерно 30 разных полипептидов, нуклеопоринов. В противоположность ЯПК, рибосомы содержат по одной копии четырех типов РНК, и около 80 различных полипептидов.
Ядерный поровой комплекс (ЯПК) представляют собой бочкообразные структуры, проходящие через ядерную оболочку и несколько выступающие за пределы обеих мембран, образуя структуры кольцеобразной формы. Как показано на рисунке ниже, для большинства ЯПК характерна симметрия восьмого порядка. Со стороны ядра и цитоплазмы пора выглядит по-разному. Части ЯПК, выступающие в нуклеоплазму и цитоплазму, называются терминальными структурами.
Модели, описывающие строение ядерной поры, были предложены на основании анализа сотен электронных микрофотографий отдельных ЯПК, полученных при высоком разрешении. Для наложения изображений и их анализа использовались математические методы, дающие возможность получить усредненную картину распределения электронной плотности или обобщенную структуру кора ЯПК (этот метод не обеспечивает оптимального разрешения терминальных структур).
На рисунке ниже представлены модели структуры кора ЯПК клеток дрожжей и Xenopus. Размеры ЯПК клеток S. cerevisiae и других одноклеточных эукариот составляют около 60 х 106 Да — т. е. вполовину меньше, чем размеры ЯПК многоклеточных организмов. Однако, несмотря на разницу в размерах, их общее строение одинаково. Размер центрального канала поры, а также его транспортные свойства у Metazoa и дрожжей также одинаковы. В настоящее время наилучшие изображения ЯПК получены методом криоэлектронной микроскопии.
 ЯПК характеризуется осью симметрии восьмого порядка, расположенной перпендикулярно оболочке ядра.
ЯПК характеризуется осью симметрии восьмого порядка, расположенной перпендикулярно оболочке ядра.
Иногда встречаются поры, обладающие симметрией седьмого или девятого порядка.
Симметрия восьмого порядка легко видима на увеличенных изображениях отдельных ЯПК (фотографии внизу).
Усредненная микрофотография в электронном микроскопе, полученная по результатам нескольких сотен отдельных фотографий (внизу справа).
Как показано на рисунке ниже, при любом расположении ЯПК происходит слияние наружной и внутренней мембран ядра. Мы не знаем, каким образом это происходит, но, скорее всего, слияние является интегральной частью процесса сборки ЯПК в ядерной оболочке. Комплексы закрепляются в оболочке с помощью интегральных мембранных белков, которые являются частью основной структуры. Эти белки проходят в перинуклеарное пространство. ЯПК проникают в ядерную ламину и также скрепляются с ней.
Фиксация клеток позволяет наблюдать этапы продвижения материалов по каналу ЯПК. При исследовании препаратов в электронном микроскопе часто видно, что полость центрального канала заполнена плотной средой. Относительно состава этой среды существуют различные точки зрения. Согласно одной из них, среда представляет собой часть ЯПК, которая наиболее прочно связывается с карго, транспортируемым через канал. Поэтому для ее обозначения используют термин транспортер или втулка. Альтернативная точка зрения предполагает, что в действительности электронноплотный материал является комплексом карго с рецептором. На основании исследований, проведенных с помощью электронного микроскопа с высоким разрешением, этот материал, по-видимому, характеризуется различными размерами и переменной локализацией в канале ЯПК, что более совместимо с точкой зрения о том, что он состоит из комплексов карго-рецептор.
В некоторых клетках ЯПК находятся не только в ядерной оболочке, но также в структурах, называемых окончатыми мембранами, которые представляют собой стопки двойных мембран, содержащих ЯПК и расположенных в цитоплазме. Часто ЯПК в слоях окончатых мембран располагаются так, как показано на рисунке ниже. Обычно окончатые мембраны присутствуют в ооцитах беспозвоночных и позвоночных, однако могут наблюдаться также и в других типах клеток. Происхождение и функции их остаются неизвестными.
Ядерный поровой комплекс (ЯПК) клеток млекопитающих трудно отделить от ядерной оболочки, поскольку они обычно связаны с ламиной, представляющей собой нерастворимую структуру, и поэтому являются неудобным объектом исследования. Поскольку окончатые мембраны не имеют подстилающей ламины, они представляют собой ценный источник выделения ЯПК для последующих биохимических и цитологических исследований. Вероятно, ЯПК окончатых мембран имеют такое же строение и состав, как и комплексы пор ядерной оболочки.
 ЯПК обладают различными терминальными структурами.
ЯПК обладают различными терминальными структурами.
Как показывают исследования в электронном микроскопе,
со стороны ядра по форме они напоминают корзину (слева),
а со стороны цитоплазмы представлены фибриллами (справа).  Цитоплазматические фибриллы и ядерные корзины ядерных пор,
Цитоплазматические фибриллы и ядерные корзины ядерных пор,
видимые в трансмиссионном электронном микроскопе. 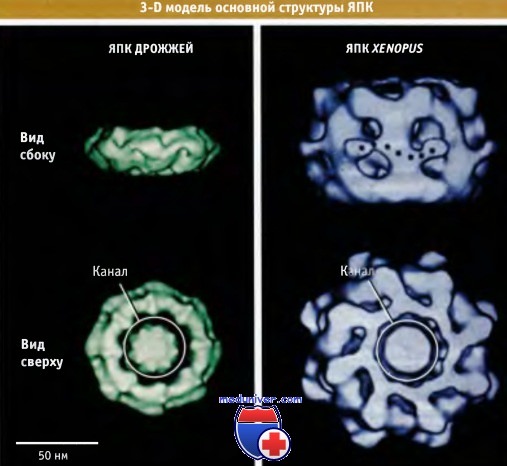 Трехмерные компьютерные модели ЯПК,
Трехмерные компьютерные модели ЯПК,
иллюстрирующие распределение средней электронной плотности.
Представлен вид моделей сбоку, вдоль плоскости ядерной оболочки, и сверху, перпендикулярно оболочке. 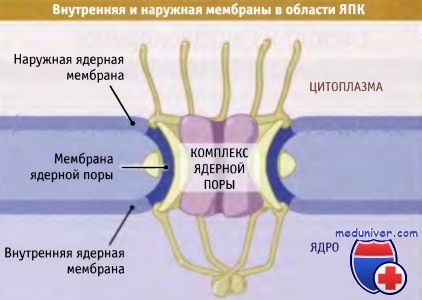 Наружная и внутренняя мембраны ядерной оболочки соединяются в области комплекса ядерной поры.
Наружная и внутренняя мембраны ядерной оболочки соединяются в области комплекса ядерной поры. 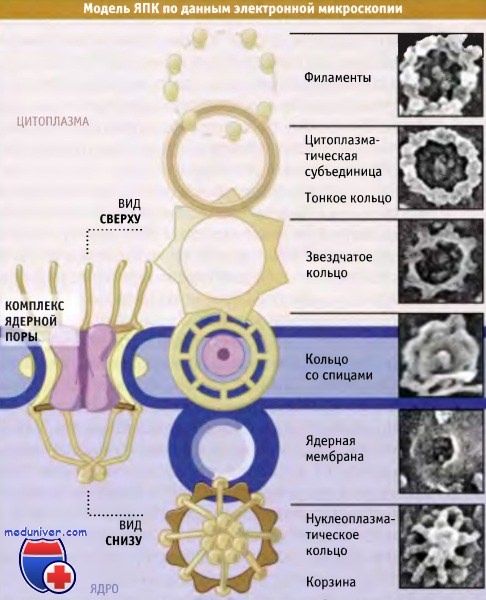 Предполагается, что ЯПК собраны из модульных компонентов.
Предполагается, что ЯПК собраны из модульных компонентов.
Показаны фотографии этих компонентов, сделанные в электронном микроскопе на разных стадиях сборки ЯПК после митоза. 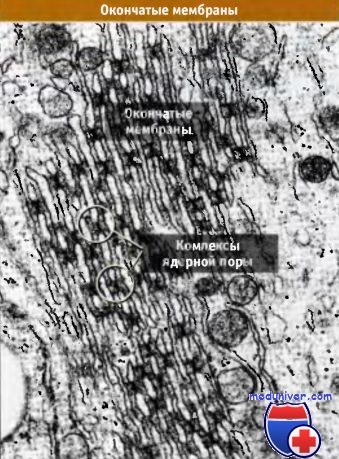 Окончатые мембраны в ооцитах Xenopus.
Окончатые мембраны в ооцитах Xenopus.
Фотография получена в трансмиссионном электронном микроскопе.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Что такое перинуклеарное пространство
 | |
| | ТОМ 4, СТ. 28 (стр. 31) // Февраль, 2003 г. СТАРЕНИЕ КАК УНИВЕРСАЛЬНАЯ ХРОНИЧЕСКАЯ «БОЛЕЗНЬ КОЛИЧЕСТВЕННЫХ ПРИЗНАКОВ» : КЛЕТОЧНОЕ СТАРЕНИЕ И РНК-ЗАВИСИМАЯ ИОННАЯ МОДУЛЯЦИЯ ПРОДУКТИВНОСТИ ГЕНОВ Рассматривается гипотеза, утверждающая, что по своему первичному механизму биологическое старение есть «болезнь количественных признаков», на течение которой влияет характер функционирования ионной модуляции транскрипционной продуктивности генов. Эта ионная регуляция основана на участии так называемой фонтанной РНК (фРНК). Согласно рассматриваемой фонтанной теории ионной модуляции экспрессии генов, их продуктивность меняется в зависимости от поступления ионов из перинуклеарной цистерны внутрь ядра через предполагаемые фРНК-зависимые ионные каналы внутренней ядерной мембраны. Когда теломеры делаются в стареющих клетках слишком короткими, фРНК-зависимые ионные каналы могут становиться (вероятно, до G1/S перехода в клеточном цикле) недоступными для субтеломерных генов, что обусловлено особенностями взаимодействия конденсированного укороченного теломерного спейсера с так называемым армирующим щитом ядерной оболочки. Это может количественно менять продуктивность соответствующих генов и служить фактором клеточного старения. Ключевые слова: старение, количественные признаки, ядро, фонтанные РНК (фРНК), ионы, теломера, энхансеры, транспозоны. ФОНТАННЫЙ МЕХАНИЗМ И ПАТОЛОГИЯ ФОНТАННЫЙ МЕХАНИЗМ И УКОРОЧЕНИЕ ТЕЛОМЕР ЗАКЛЮЧЕНИЕ AGING AS AN UNIVERSAL CHRONIC «DISEASE OF QUANTITATIVE TRAITS»: CELLULAR SENESCENCE AND RNA-DEPENDENT IONIC MODULATION OF GENES PRODUCTIVITY Key words: senescence, quantitative traits, nucleus, fountain RNAs (fRNAs), ions, telomere, enhancers, transposons.  Приветствую, уважаемые дети! Сегодня я хочу рассказать  Приветствую вас, уважаемые дети! Сегодня я хочу рассказать  Приветствую вас, дорогие дети! Сегодня я хочу рассказать  Приветствую вас, дорогие дети! Сегодня я хочу рассказать |
 Фундаментальные исследования
Фундаментальные исследования