Полуавтономные органоиды: строение и функции. Теория симбиогенеза.
Полуавтономные органоиды- органоиды, содержащие собственные ДНК, все типы РНК, рибосомы, способные синтезировать белки, а также преобразовывать Е в ЕАТФ (синтез АТФ).
К таким органоидам относятся двумембранные органоиды: митохондрии и пластиды.
Митохондрии и пластиды способны к самовоспроизведению и никогда не образуются из других компонентов клетки. Вне клеток они существовать не могут.
Хлоропласты имеют зеленый цвет, обусловленный присутствием основного пигмента — хлорофилла.
Возможные пути деления митохондрий: при образовании перегородок или перетяжек
Функции: 
согласно этой теории, митохондрии и хлоропласты произошли от симбиотических прокариотических организмов, захваченных протоэукариотом в результате фагоцитоза. Этот протоэукариот, о видимому, представлял собой амебоидный гетеротрофный, анаэробный организм с уже развитыми эукариотическими признаками.
Симбиоз большой анаэробной клетки (вероятно, относящейся к архебактериям и сохранившей ферменты гликолитического окисления) с аэробными бактериями оказался взаимовыгодным, причем бактерии со временем утратили самостоятельность и превратились в митохондрии
Потеря самостоятельности связана с утратой части генов, которые перешли в хромосомный аппарат клетки-хозяина. Но все же митохондрии сохранили собственный белоксинтезирующий аппарат и способность к размножению.
В пользу симбиотического происхождения митохондрий и хлоропластов говорят многие факты. Во-первых, их генетический материал представлен одной кольцевой молекулой ДНК (как и у прокариот), во-вторых, их рибосомы по массе, по строению рРНК и рибосомальных белков близки к таковым у аэробных бактерий и синезеленых. В-третьих, они размножаются как прокариоты и наконец, механизмы белкового синтеза в митохондриях и бактериях чувствительны к одним антибиотикам (стрептомицину), а циклогексимид блокирует синтез белка в цитоплазме. Кроме того, известен один вид амеб, которые не имеют митохондрий и живут в симбиозе с аэробными бактериями, а в клетках некоторых растений обнаружены цианобактерии (синезеленые), сходные по строению с хлоропластами.
15. Ядро: строение и функции. Организация генетического материала эукариотической клетки.
1. Хранение наследственной информации
2. Реализация наследственной информации
3. Воспроизведение молекулы днк
4. Передача информации дочерним клеткам при клеточном делении
Геном (совокупность наследственного материала, заключенного в клетке)каждой клетки несет информацию:
О первичной структуре всех белков всех клеток всего организма (последовательность аминокислот)
О первичной структуре примерно (последовательность нуклеотидов) 60 видов тРНК и различных рРНК (у эукариотов 4 вида)
О программе использования данной информации в разных клетках и разных моментах онтогенеза (онтогенез-индивидуальное развитие организма)
Хроматин или хромосомы (хроматин в неделящемся ядре, хромосомы-в митотическом)
Размер: от 1 мкм (простейшие) до 1 мм (в яйцеклетках рыб и земноводных)
Оболочка: внутренняя и наружная
Кариоплазма – желеобразный раствор, заполняет пространство между структурами ядра (хроматином и ядрышками)
Ядрышко не окружено мембраной, содержит фибриллярные белковые нити и РНК, исчезают в начале деления клетки и восстанавливаются после его окончания. В ядрышках происходит формирование рибосом, синтез ядерных белков. Образуются на участках перетяжек хромосом. участки хромосом, на которых происходит синтез рибосомных рибонуклеиновых кислот (рРНК
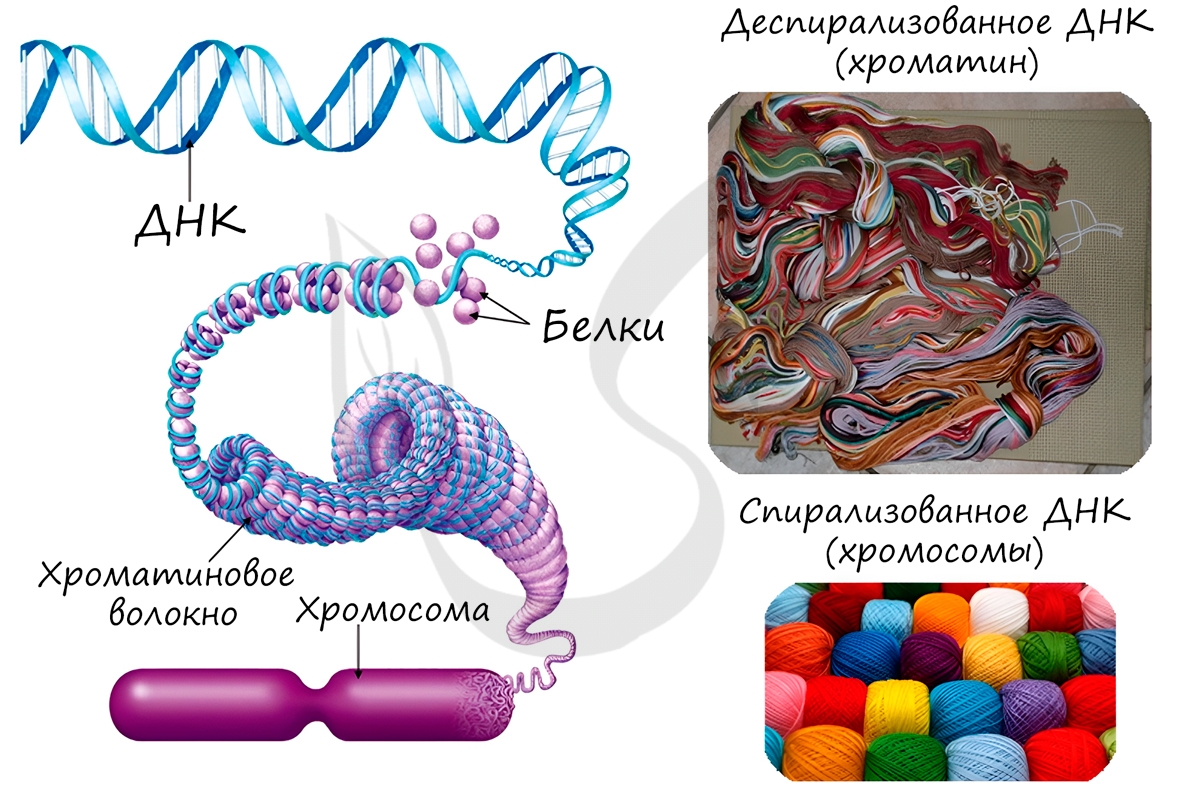
Хроматин- деспирализованная форма существования хромосом
ГЕТЕРОХРОМАТИН – участки хромосом, находящиеся в конденсированном (упакованном) состоянии в течение всего клеточного цикла. Таким образом, гетерохроматиновые участки в генетическом отношении являются практически неактивными.
ЭУХРОМАТИН – основная часть митотических хромосом, в которой локализована большая часть функциональных генов. Эухроматин претерпевает обычный цикл компактизации-декомпактизации во время митоза.
Хромосомы- органоиды ядра, являются носителями генов и определяют наследственные свойства клеток и организма.


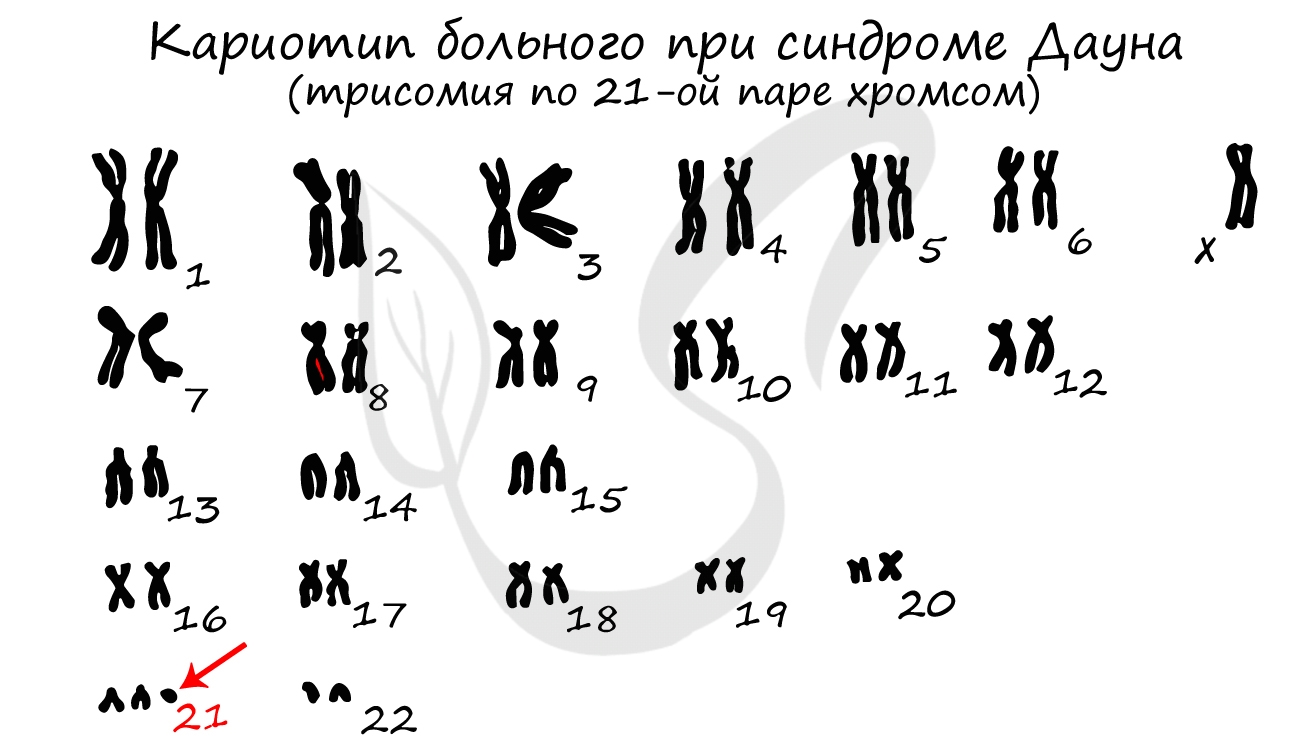
Кариотип- совокупность полного набора хромосом, присущего клеткам данного биологического вида (понятие ввел Левитский в 1924)
Кариограмма – систематизированное изображение хромосом, расположенных в ряд по мере убывания их длины.
Организация генетического материала эукариотической клетки
Длина ДНК диплоидного набора хромосом человека составляет примерно 174 см., средняя длина ДНК одной хромосомы – 5 см. В ядре длина одной хромосомы составляет 0,5 – 1 микрон. Такая упаковка двойной спирали ДНК объясняется ее дальнейшей последовательной компактизацией.
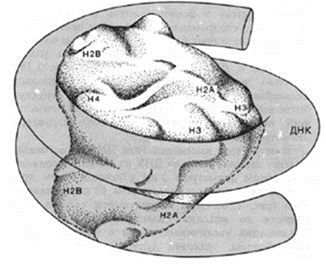
Белковый кор (сердцевина) содержит набор из 4 пар гистоновых белковН2А, Н2В, Н3, Н4. Это самые консервативные белки в любом геноме. Они практически одинаковы у гороха и у человека.
Нуклеосомы связываются участками ДНК (линкерная ДНК) свободными от контакта с белковым кором.
Укладка линкерного участка ДНК (60-80 п.н.) и соединение нуклеосом друг с другом идут с помощью гистона Н1. Молекула этого белка имеет центральную (глобулярную) часть и вытянутые «плечи». Центральная часть прикрепляется к специфическому участку на поверхности кора, вытянутые «плечи» соединяют соседние нуклеосомы. При этом ДНК наматывается на соседние коры каждый paз в противоположном направлении
Выделить нуклеосомы можно непродолжительной обработкой хромосом ферментами дезоксирибонуклеазами. При этом расщепляются участки состыковки нуклеосом. В геноме человека содержатся 1,5 х 107 нуклеосом.
Нуклеосомный уровень повышает плотность упаковки ДНК в 7-10 раз.
2. Нуклеомерный уровень. Дальнейшая компактизация ДНК в составе хроматина связана с образованием нуклеосомных комплексов Образуется компактная хроматиновая фибрилла построенная либо по типу соленоида (спиральный тип укладки), либо по нуклеомерному типу (4-12 нуклеосом образуют глобулу).
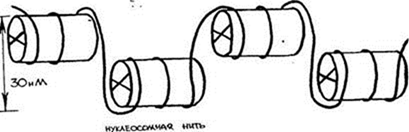
Нуклеомерная укладка хроматина способствует укорочению нити ДНК примерно в 6 раз, а оба уровня приводят к компактизации ДНК в среднем в 50 раз (42-60).
3. Хромомерный уровень.
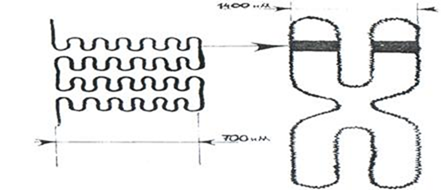
Следующий этап компактизации ДНК связан с образованием петлеобразных структур, которые называются хромомерами. При этом возможны два пути упаковки ДНК с помощью негистоновых белков: 
Укорочение фибриллы на этом уровне происходит в среднем 25 раз, а на всех 3 уровнях в 1000-1500 раз.

5.Хромосомный уровень. Дальнейшая компактизация хромосом обеспечивается петельной укладкой хромонемной нити, что сокращает их длину примерно в 10 раз.
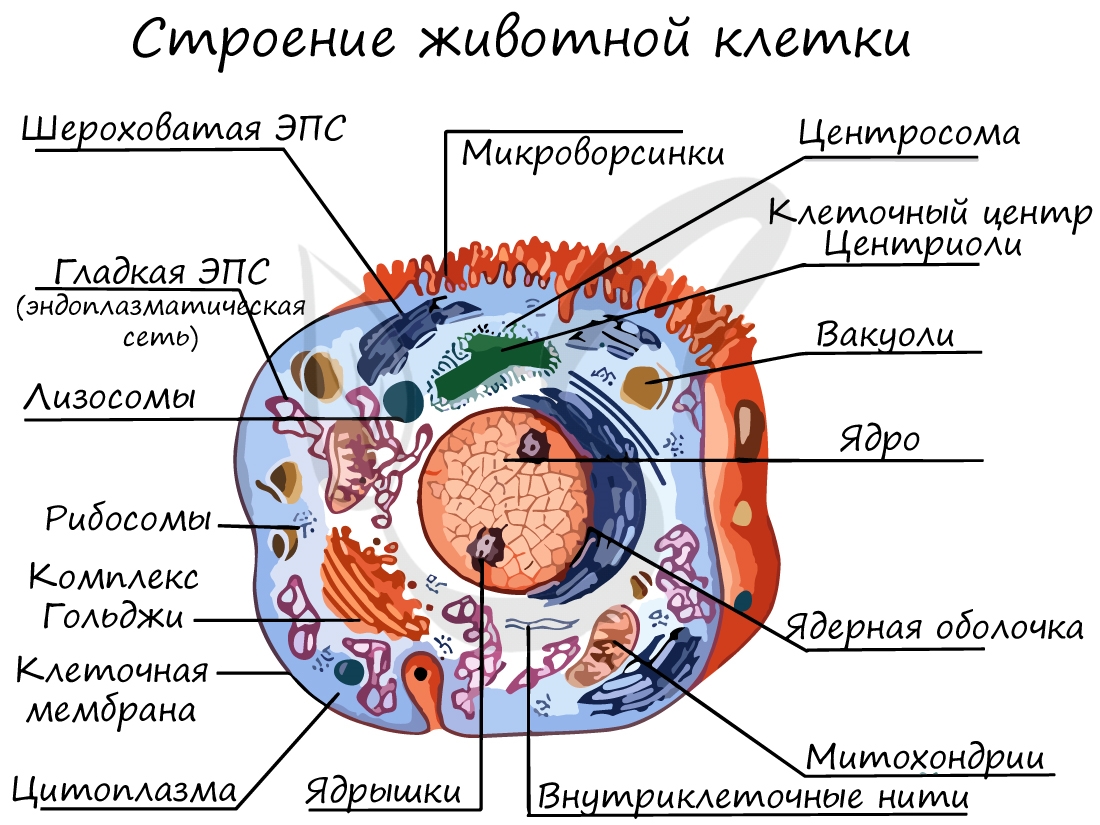
Органоиды клетки
Клеточная мембрана (оболочка)
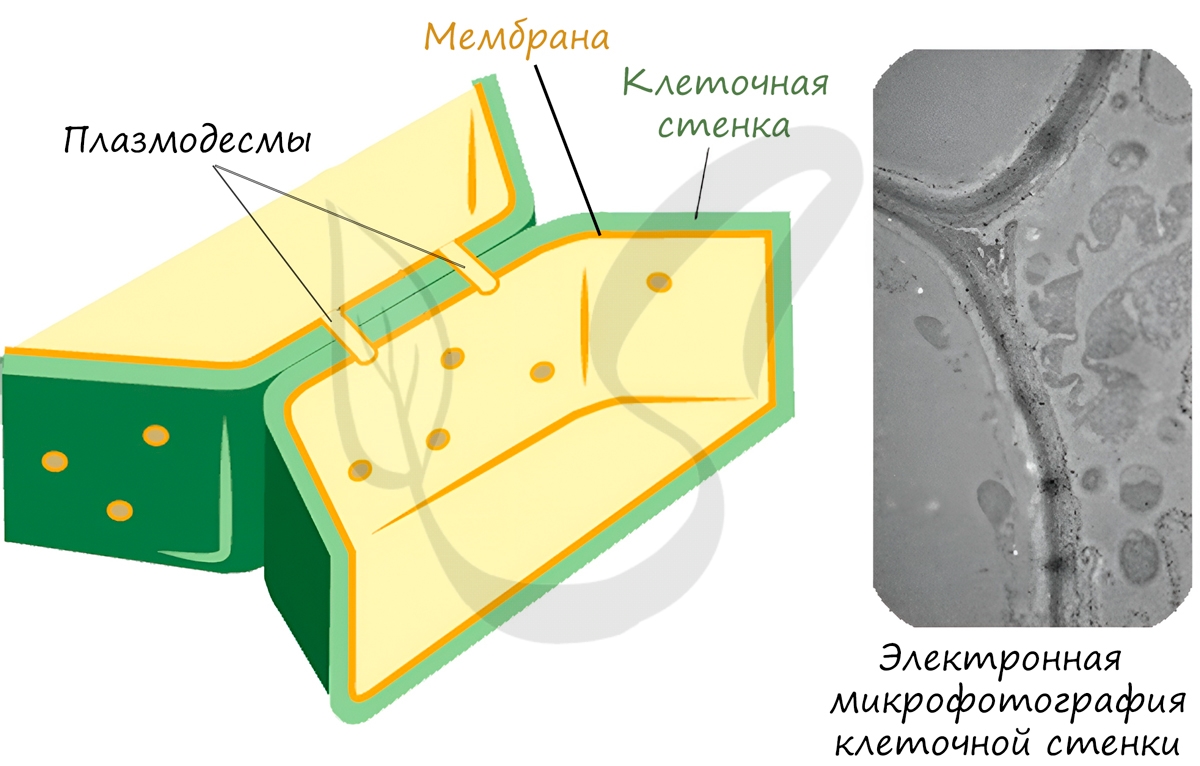
Запомните, что в отличие от клеточной стенки, которая есть только у растительных клеток и у клеток грибов (она придает им плотную, жесткую форму) клеточная мембрана есть у всех клеток без исключения! Этот важный момент объясню еще раз 🙂 У клеток животных имеется только клеточная мембрана, а у клеток растений и грибов есть и клеточная стенка, и клеточная мембрана.
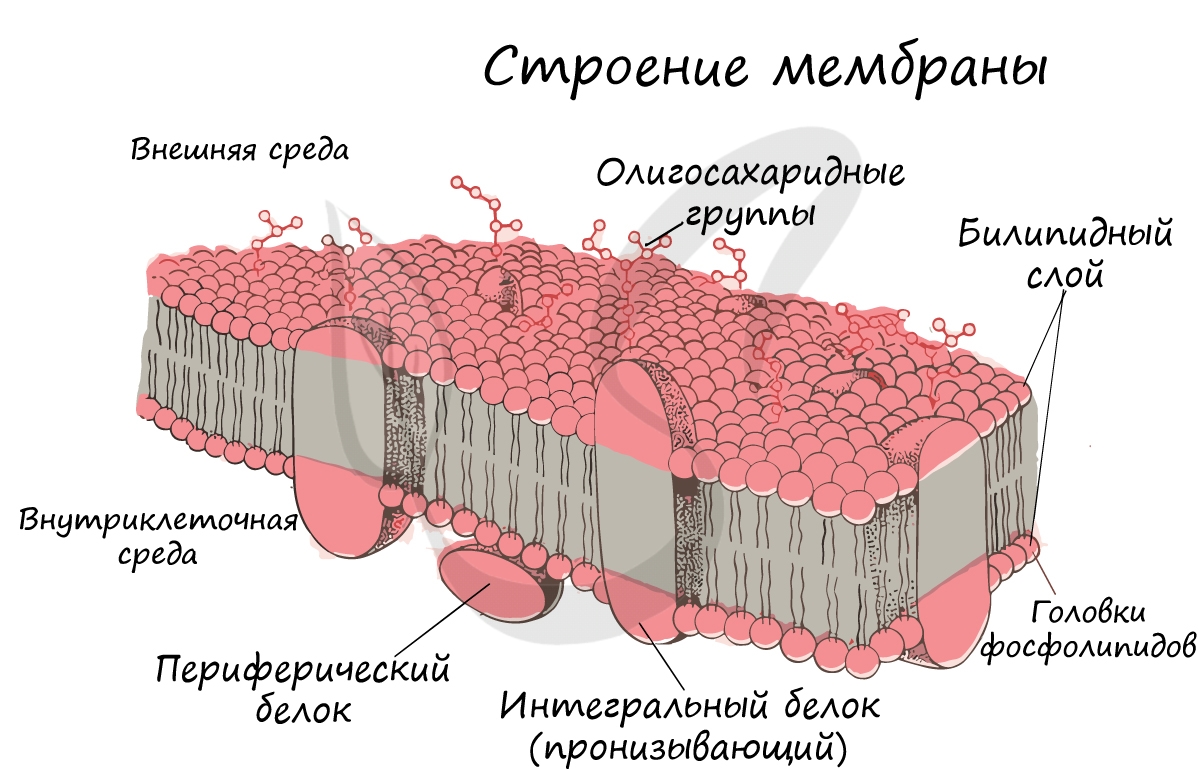
Интегральные (пронизывающие) белки образуют каналы, по которым молекулы различных веществ могут поступать в клетку или удаляться из нее. «Заякоренные» молекулы олигосахаридов на поверхности клетки образуют гликокаликс, который выполняет рецепторную функцию, участвует в избирательном транспорте веществ через мембрану.
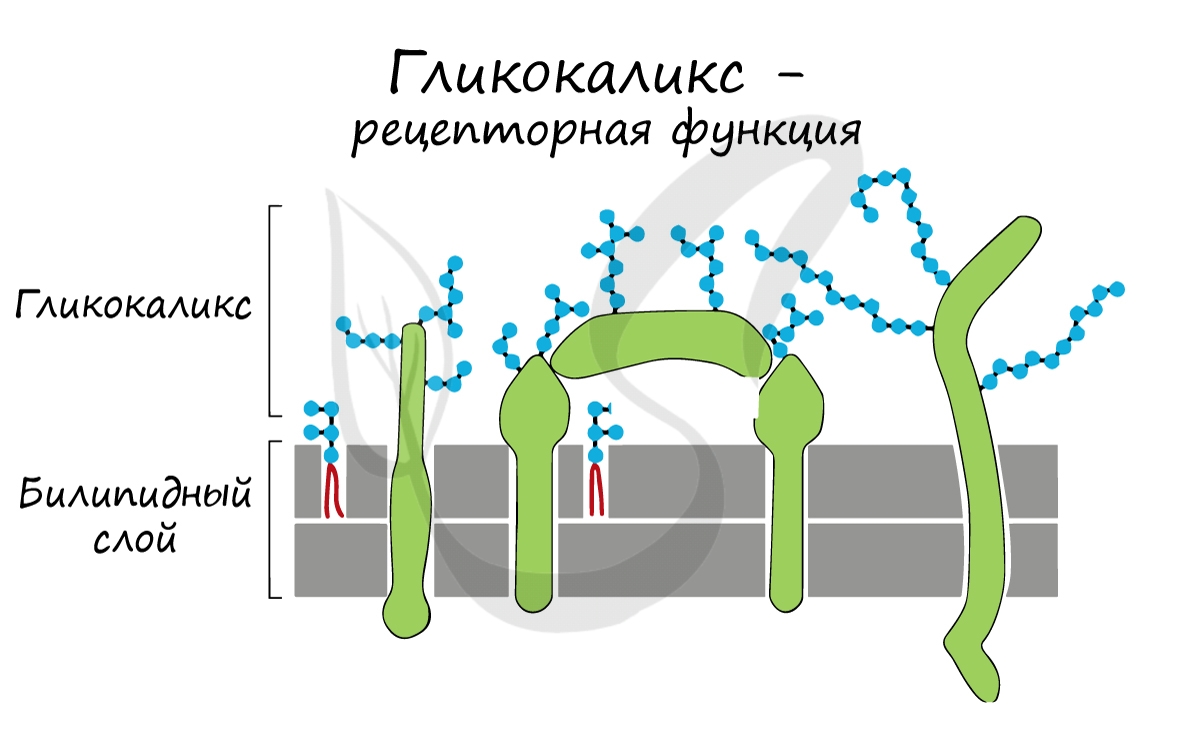
Вирусы и бактерии не являются исключением: они взаимодействуют только с теми клетками, на которых есть подходящие к ним рецепторы. Так, вирус гриппа поражает преимущественно клетки слизистой верхних дыхательных путей. Однако, если рецепторов нет, то вирус не может проникнуть в клетку, и организм приобретает невосприимчивость к инфекции. Вспомните врожденный иммунитет: именно по причине отсутствия рецепторов человек не восприимчив ко многим болезням животных.
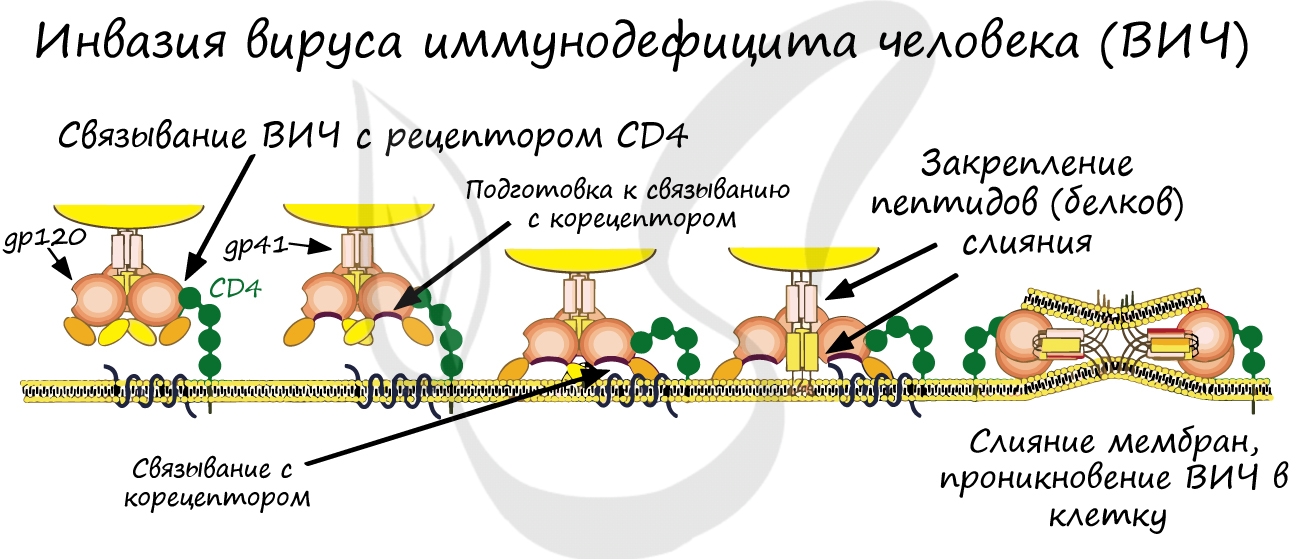
Итак, вернемся к клеточной мембране. Ее можно сравнить со стенами помещения, в котором, вероятно, вы находитесь. Стены дома защищают его от ветра, дождя, снега и прочих факторов внешней среды. Рискну предположить, что в вашем доме есть окна и двери, которые по мере необходимости открываются и закрываются 🙂 Так и клеточная мембрана может сообщать внутреннюю среду клетки с внешней средой: через мембрану вещества поступают в клетку и удаляются из нее.
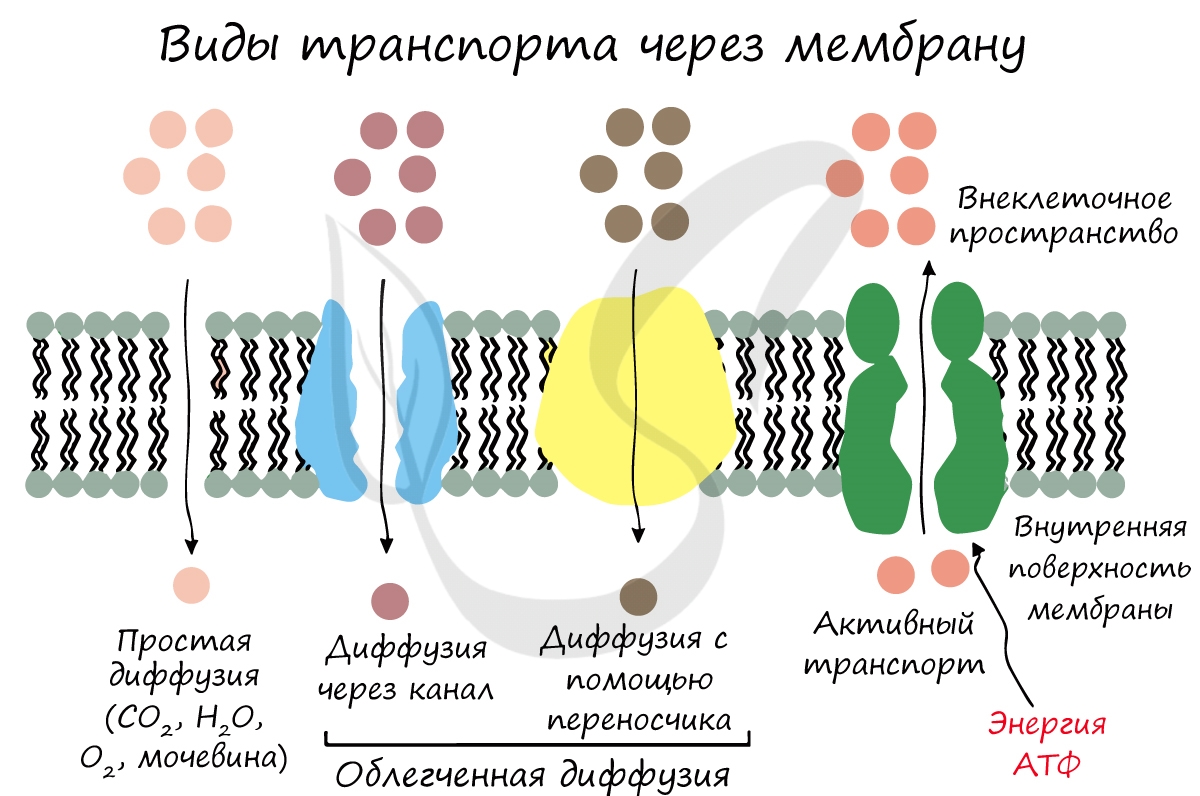
Внутрь клетки с помощью осмоса поступает вода. Путем простой диффузии в клетку попадают O2, H2O, CO2, мочевина. Облегченная диффузия характерна для транспорта глюкозы, аминокислот.
Активный транспорт чаще происходит против градиента концентрации, в ходе него используются белки-переносчики и энергия АТФ. Ярким примером является натрий-калиевый насос, который накачивает ионы калия внутрь клетки, а ионы натрия выводит наружу. Это происходит против градиента концентрации, поэтому без затрат энергии (АТФ) не обойтись.
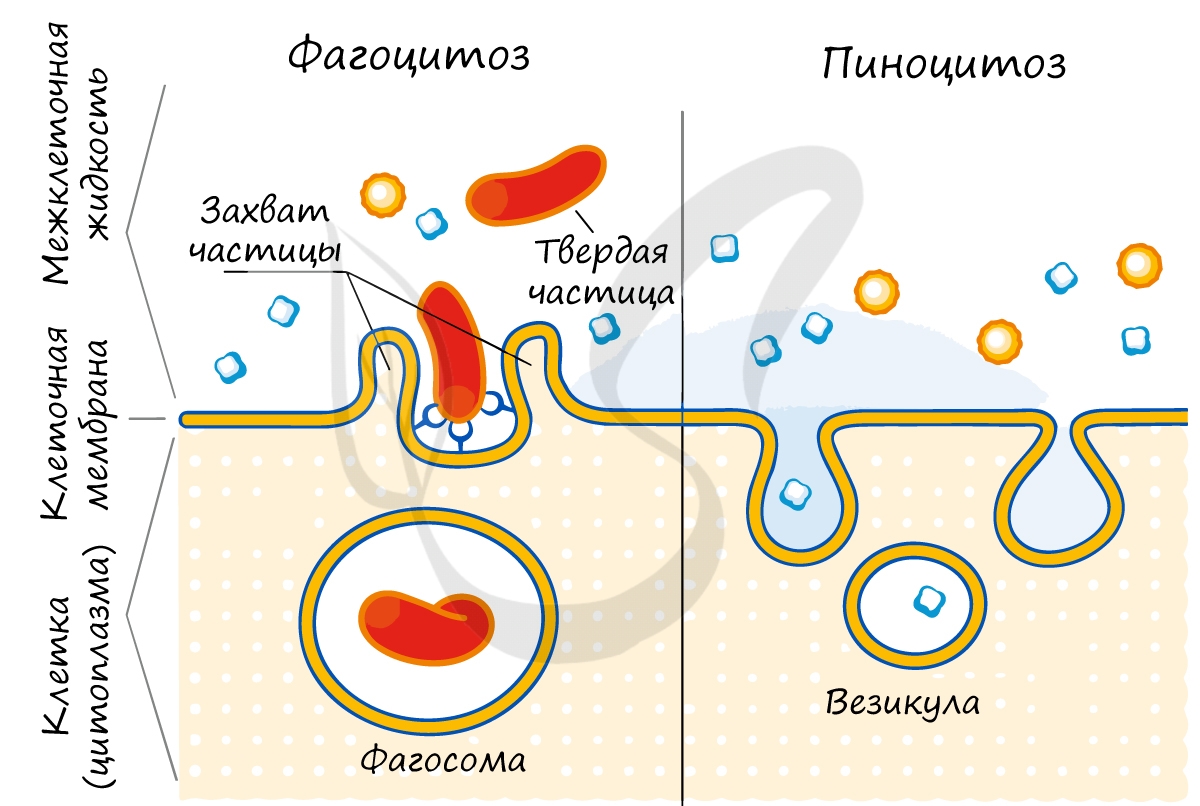
Фагоцитоз был открыт И.И. Мечниковым, который создал фагоцитарную теорию иммунитета. Это теория гласит, что в основе иммунной системы нашего организма лежит явление фагоцитоза: попавшие в организм бактерии уничтожаются фагоцитами (T-лимфоцитами), которые переваривают их.
В ходе эндоцитоза мембрана сильно прогибается внутрь клетки, ее края смыкаются, захватывая бактерию, пищевые частицы или жидкость внутрь клетки. Образуется везикула (пузырек), который движется к пищеварительной вакуоли или лизосоме, где происходит внутриклеточное пищеварение.
Клеточная стенка
Цитоплазма
Постоянное движение цитоплазмы поддерживает связь между органоидами клетки и обеспечивает ее целостность.
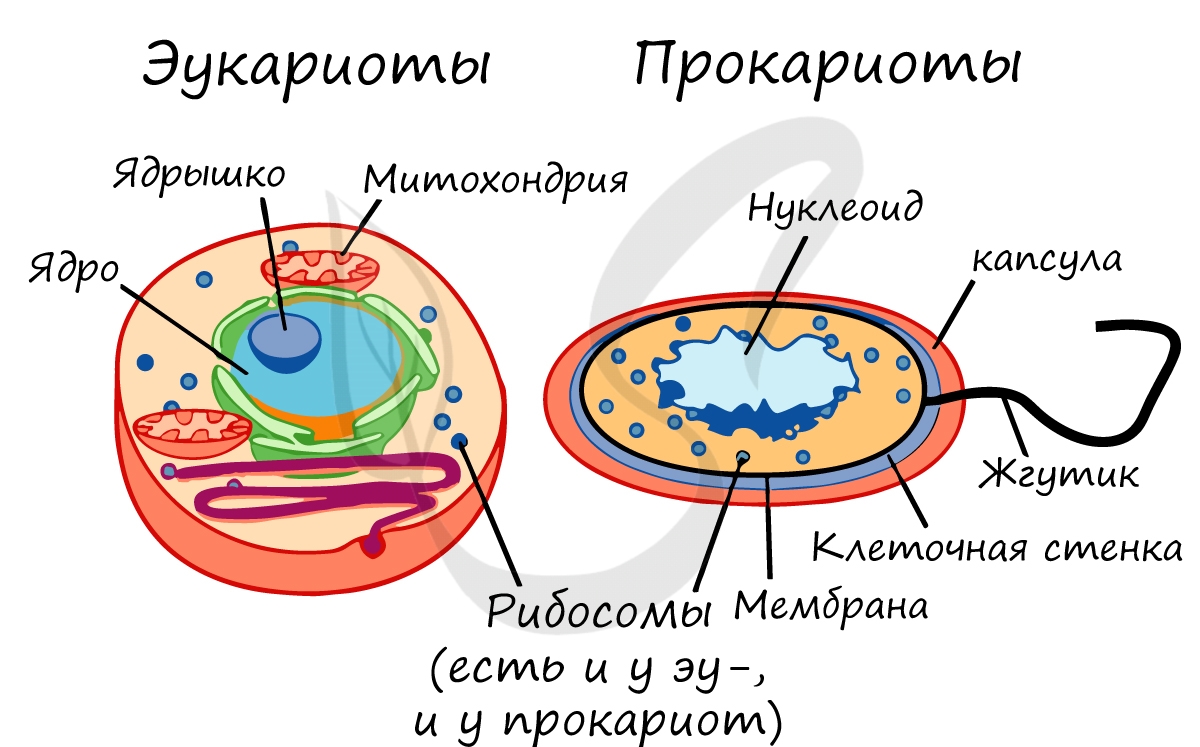
Прокариоты и эукариоты
Немембранные органоиды
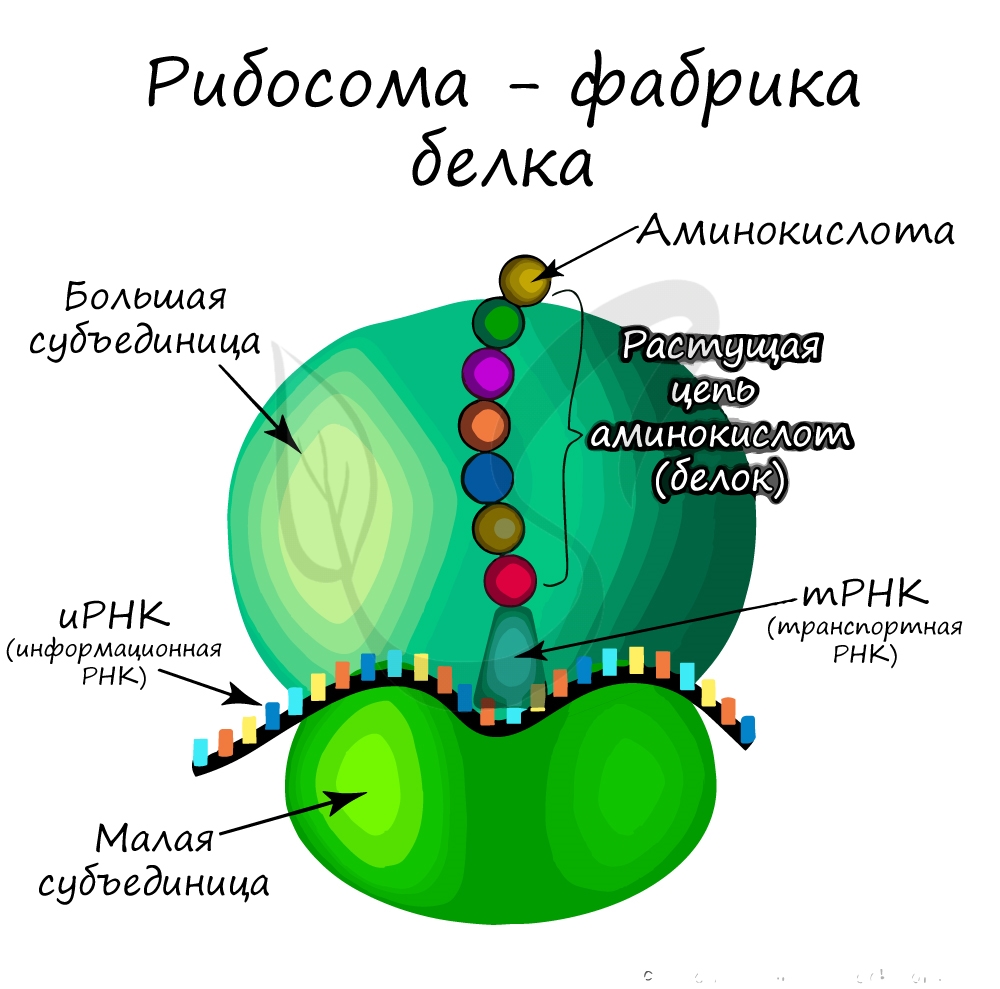
Очень мелкая органелла (около 20 нм), которая была открыта после появления электронного микроскопа. Состоит из двух субъединиц: большой и малой, в состав которых входят белки и рРНК (рибосомальная РНК), синтезируемая в ядрышке.
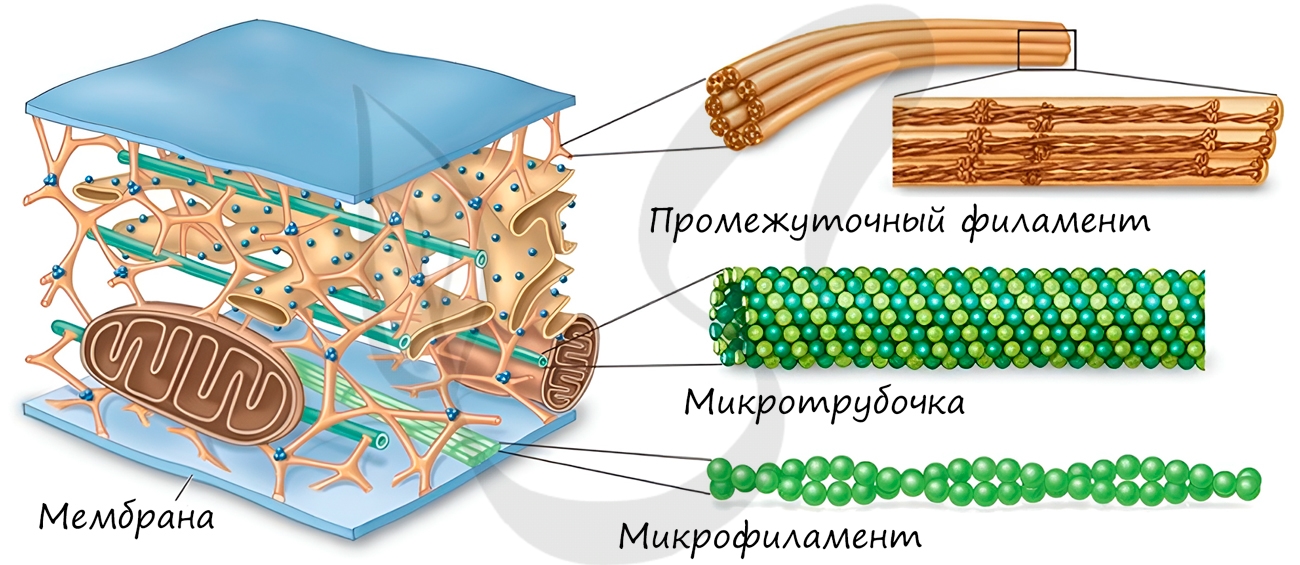
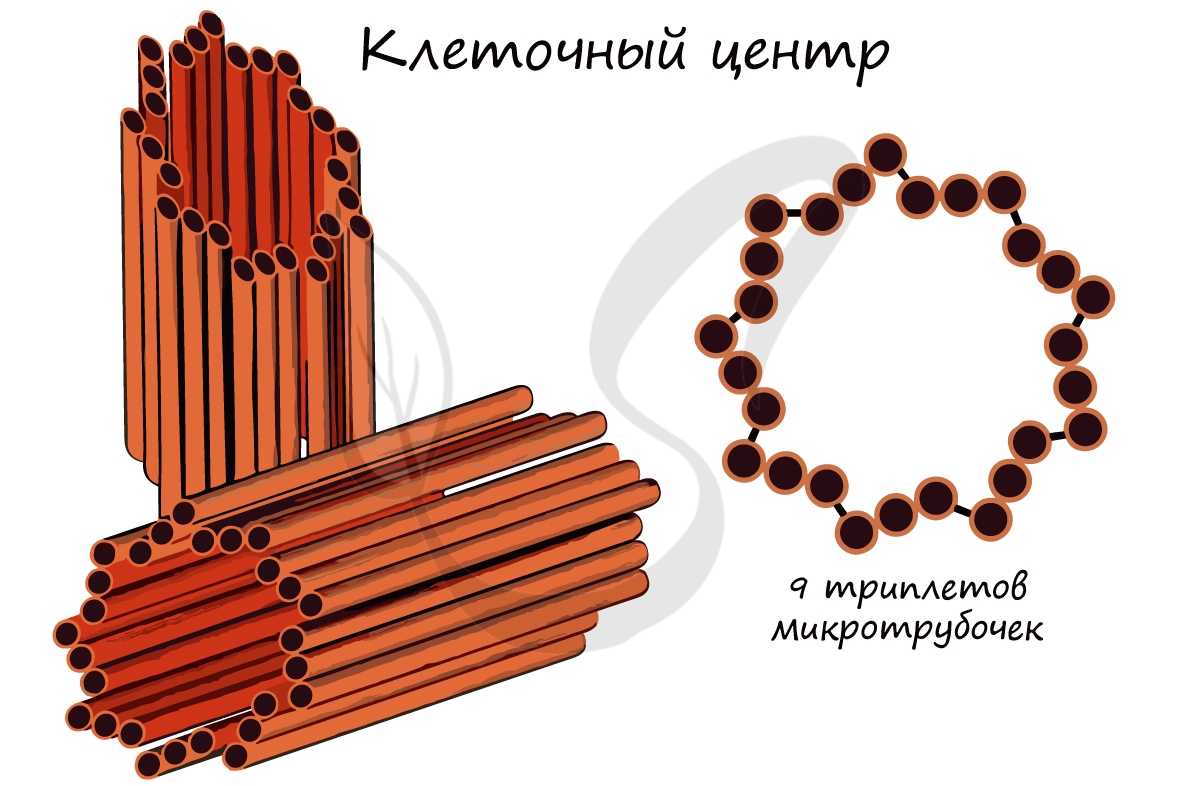
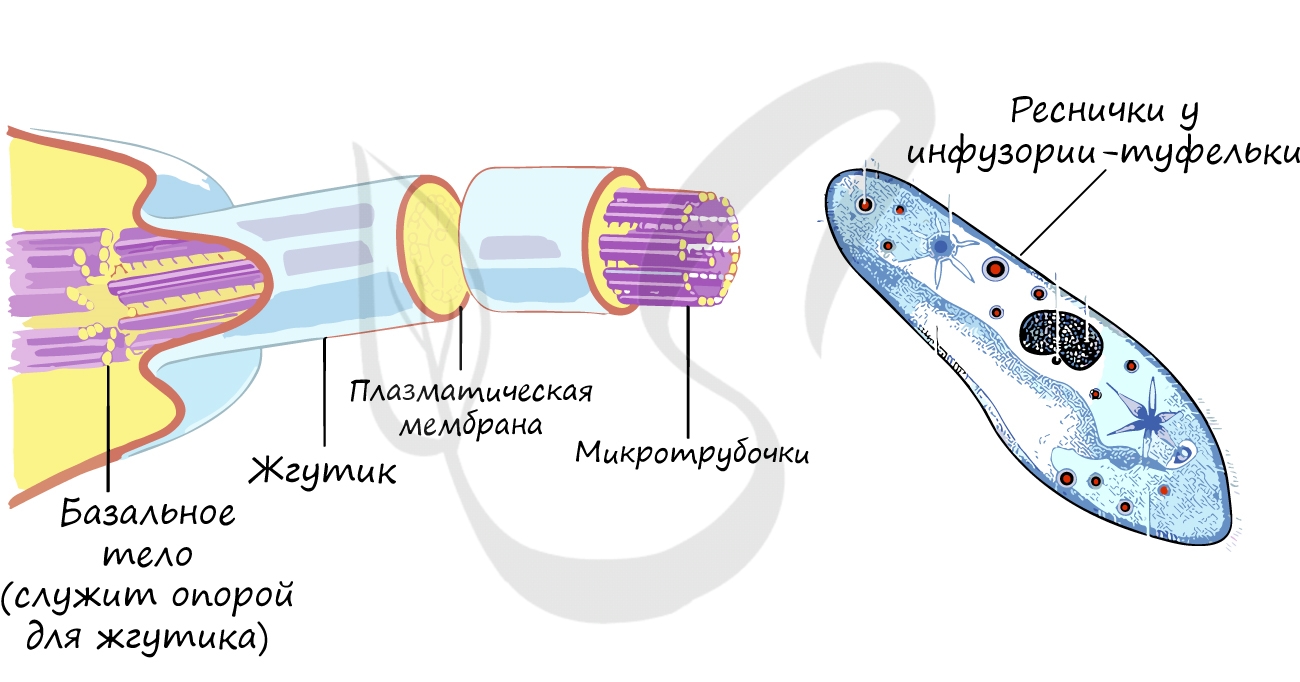
Это органоиды движения, которые выступают над поверхностью клетки и имеют в основе пучок микротрубочек. Реснички встречаются только в клетках животных, жгутики можно обнаружить у животных, растений и бактерий.
Одномембранные органоиды
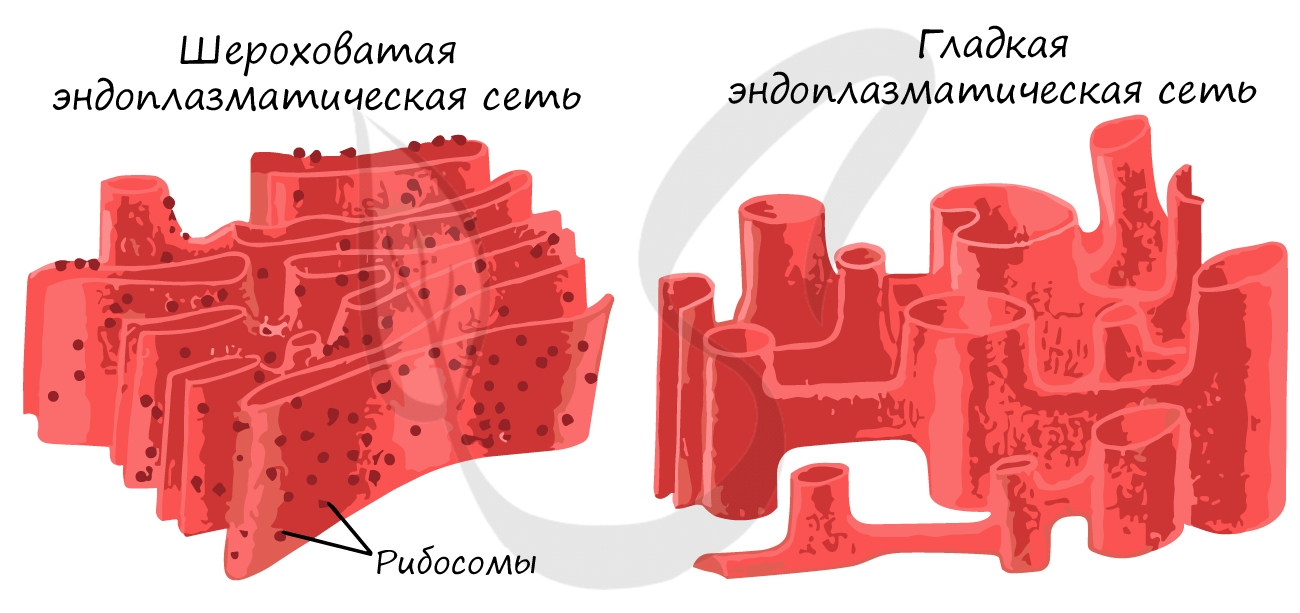
ЭПС представляет собой систему мембран, пронизывающих всю клетку и разделяющих ее на отдельные изолированные части (компартменты). Это крайне важно, так как в разных частях клетки идут реакции, которые могут помешать друг другу, что нарушит процессы жизнедеятельности.
Выделяют гладкую ЭПС и шероховатую ЭПС. Обе они выполняют функцию внутриклеточного транспорта веществ, однако между ними имеются различия. На мембранах гладкой ЭПС происходит синтез липидов, обезвреживаются вредные вещества. Шероховатая ЭПС синтезирует белок, так как имеет на мембранах многочисленные рибосомы (потому и называется шероховатой).
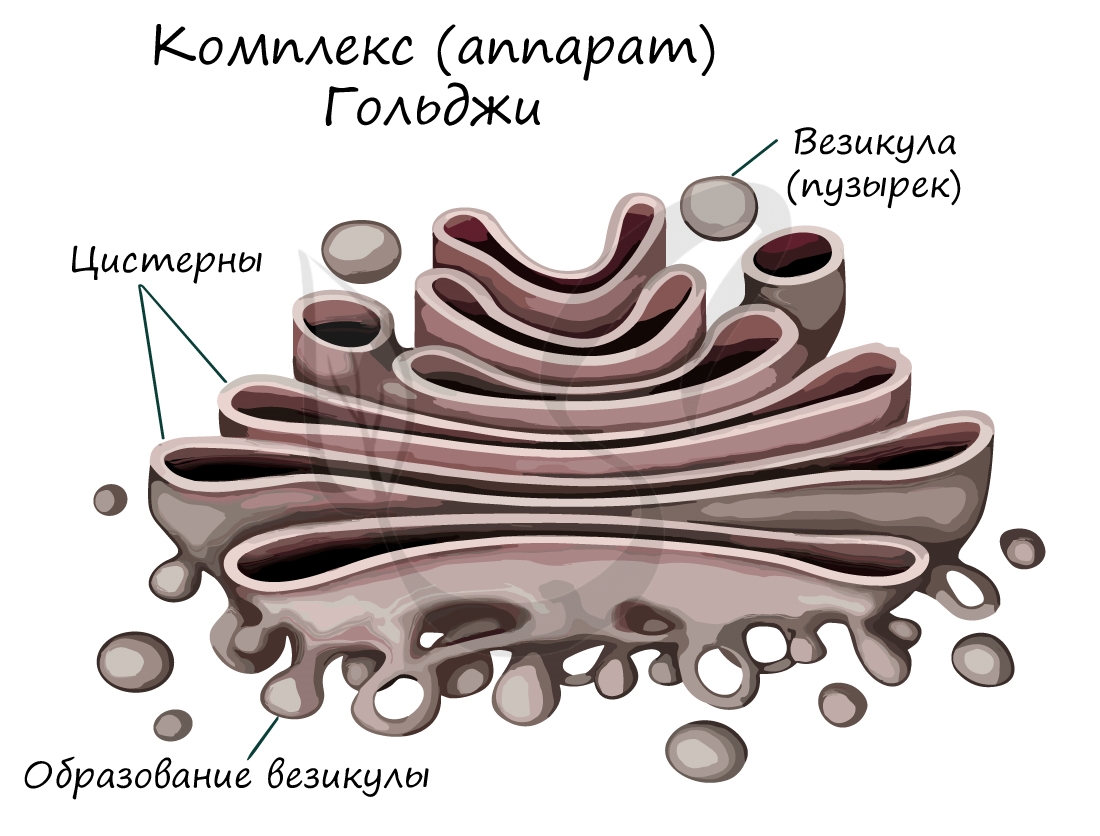
Модифицированные вещества упаковываются в пузырьки и могут перемещаться к мембране клетки, соединяясь с ней, они изливают свое содержимое во внешнюю среду. Можно догадаться, что комплекс Гольджи хорошо развит в клетках эндокринных желез, которые в большом количестве синтезируют и выделяют в кровь гормоны.
В комплексе Гольджи появляются первичные лизосомы, которые содержат ферменты в неактивном состоянии.

В ходе апоптоза ферменты лизосомы изливаются внутрь клетки, ее содержимое переваривается. Предполагают, что нарушение апоптоза в раковых клетках ведет к бесконтрольному росту опухоли.
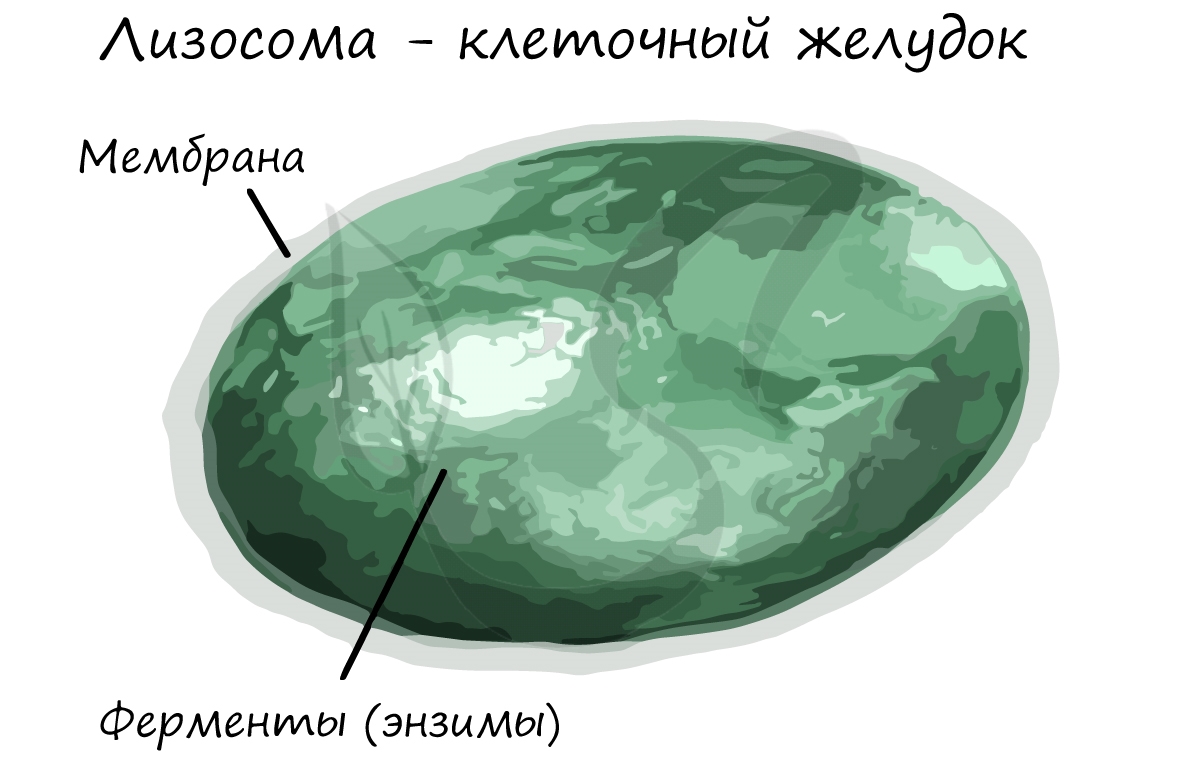
Пероксисомы (микротельца) содержат окислительно-восстановительные ферменты, которые разлагают H2O2 (пероксид водорода) на воду и кислород. Если бы пероксид водорода оставался неразрушенными, это приводило бы к серьезным повреждениям клетки.
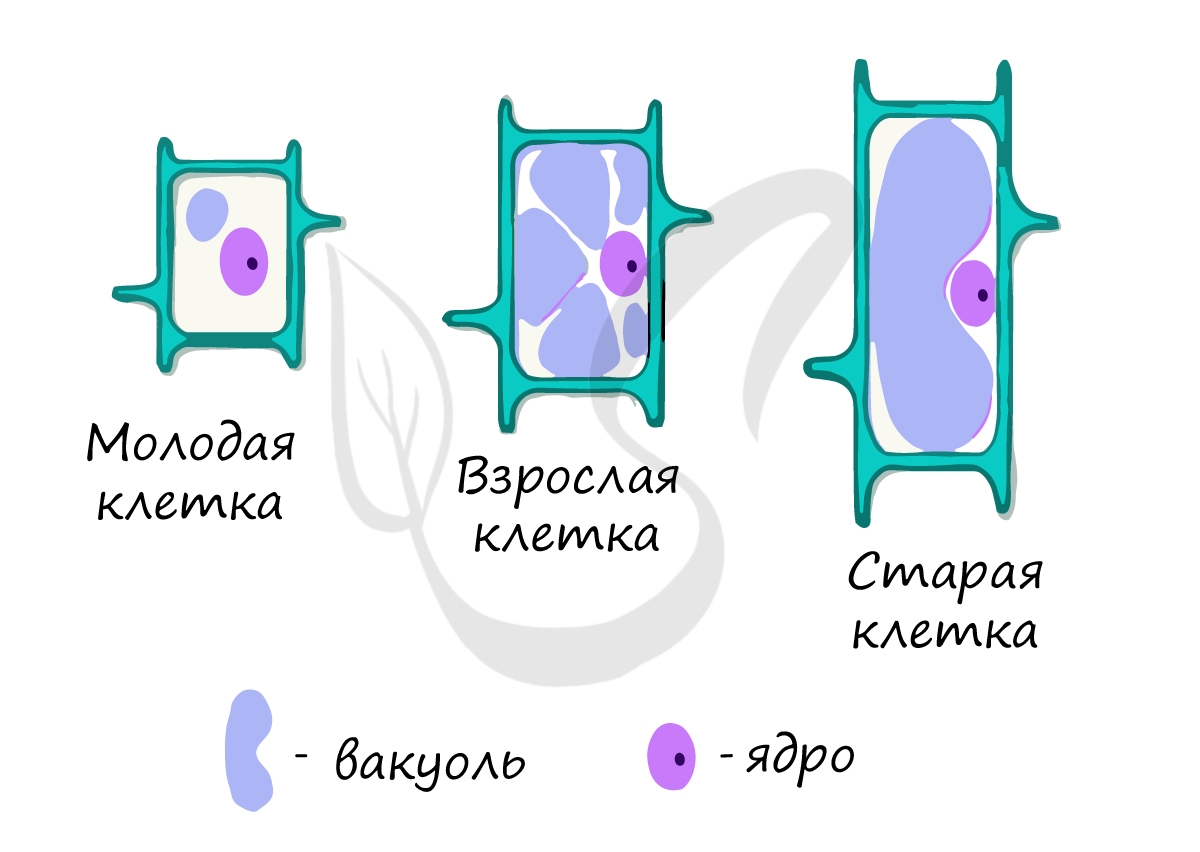
Трудно переоценить значение вакуолей в жизнедеятельности растительной клетки. Вакуоли создают осмотическое давление, придают клетке форму.
Примечательно, что по размеру вакуолей можно судить о возрасте клетки: молодые клетки имеют вакуоли небольшого размера, а в старых клетках вакуоли могут настолько увеличиваться, что оттесняют ядро и остальные органоиды на периферию.
Двумембранные органоиды
Оболочка ядра состоит из двух мембран и пронизана большим количеством ядерных пор, через которые происходит сообщение между кариоплазмой и цитоплазмой. Главными функциями ядра является хранение, защита и передача наследственного материала дочерним клеткам.
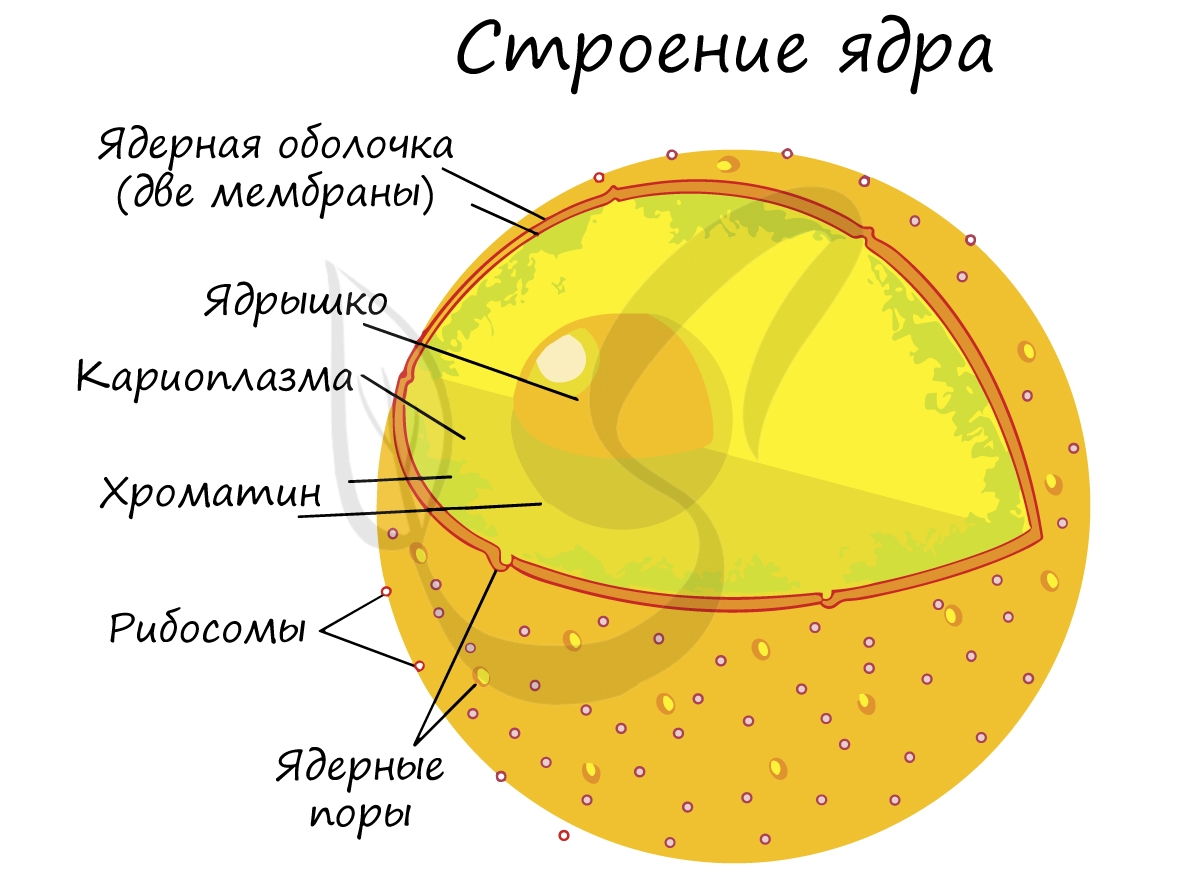
Замечу, что хромосомы видны только в момент деления клетки. Хромосомы представляют собой сильно спирализованные молекулы ДНК, связанные с белками.
Хромосомы отличаются друг от друга по строению, форме, размерам. Совокупность всех признаков (форма, число, размер) хромосом называется кариотип. Кариотип может быть представлен по-разному: существует кариотип вида, особи, клетки.
В связи с этим, митохондрия считается полуавтономным органоидом. Вероятнее всего, изначально митохондрии были самостоятельными организмами, однако со временем вступили в симбиоз с эукариотами и стали частью клетки.
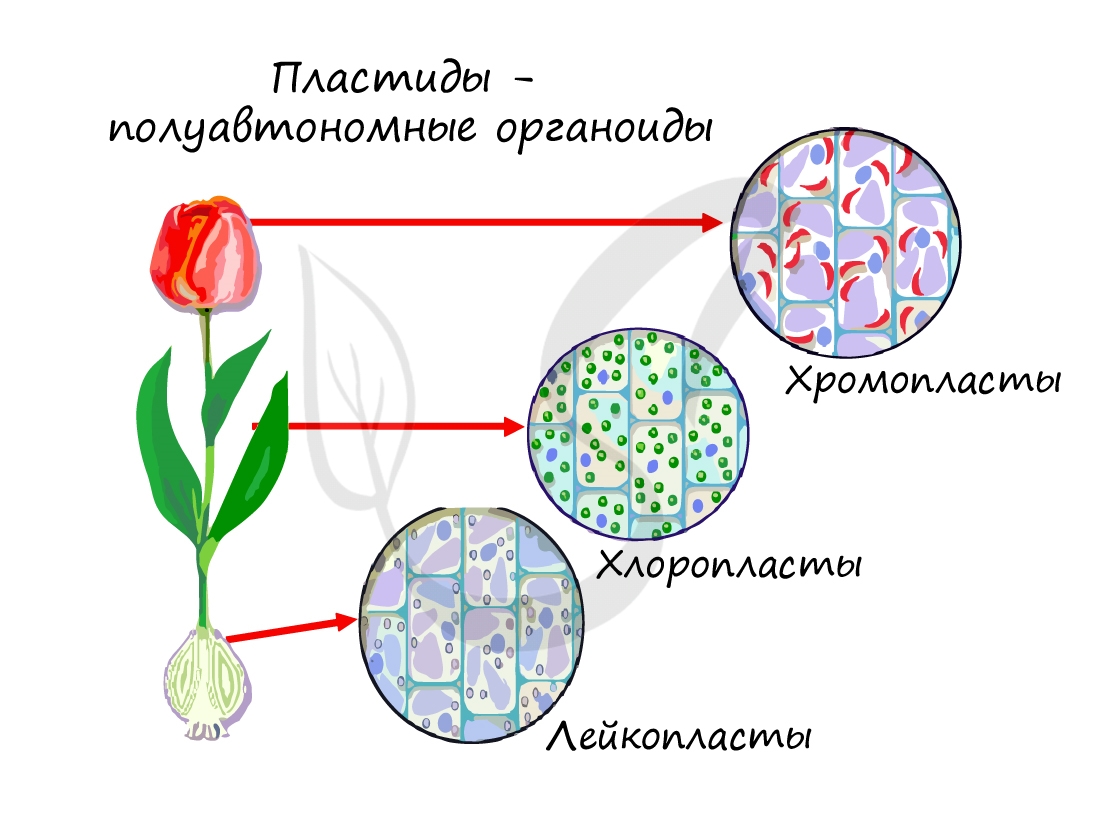
Так же, как и митохондрии, пластиды относятся к полуавтономным органоидам: в них имеется кольцевидная ДНК (находится в нуклеоиде), рибосомы.
Пластиды, которые содержат пигменты каратиноиды в различных сочетаниях. Сочетание пигментов обуславливает красную, оранжевую или желтую окраску. Находятся в плодах, листьях, лепестках цветков.
Хромопласты могут развиваться из хлоропластов: во время созревания плодов хлоропласты теряют хлорофилл и крахмал, в них активируется биосинтез каротиноидов.
Не содержат пигментов, образуются в запасающих частях растения (клубни, корневища). В лейкопластах накапливается крахмал, липиды (жиры), пептиды (белки). На свету лейкопласты могут превращаться в хлоропласты и запускать процесс фотосинтеза.
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Общая характеристика полуавтономных органоидов
Они отделены от цитоплазмы двумя мембранами – внешней и внутренней.
2. У них различают основную полость, ограниченную внутренней мб: матрикс у Мх и стромуу Пл.
3. В матриксе или в строме расположены ДНК и РНК, рибосомы, а также ферменты.
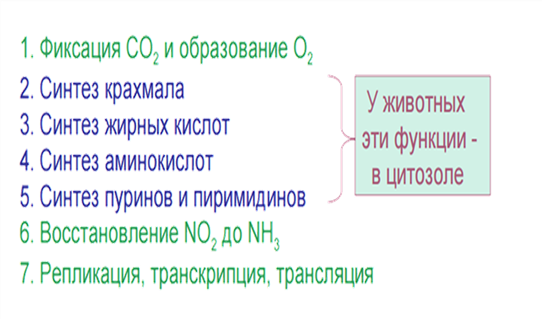
4. Мх и Пл способны к автономному (самостоятельному) синтезу ДНК, РНК, специфичных белков, жиров и углеводов.
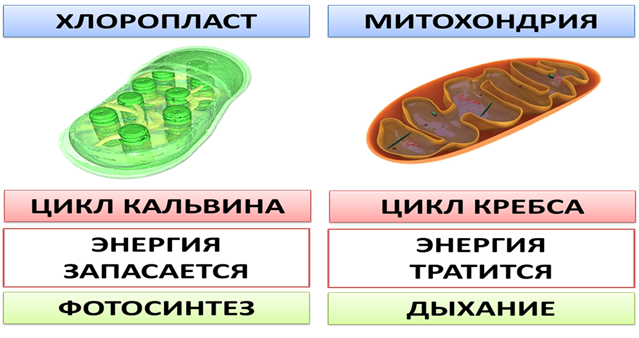
5. Главной функцией Пл является фотосинтез, а Мх – дыхание. Эти процессы связаны с синтезом АТФ.
М и т о х о н д р и и(Мх)
Мх (митос = нить, хондрион =зернышко) – гранулярные или нитевидные органоиды простейших, растений, животных и грибов. В животных клетках митохондрии двигаются, сливаются др. с др. В кл. может быть от 200 до 500 000 мх. Однако многие клетки имеют одну сильно разветвленную мх, к-рая образует митохондриальную сеть.
Мх мало в кл. зеленых растений, так как часть их ф-ций берут на себя Хпл.
Расположение Мх в кл. м. б. различным. В дифференцированных [1] растительных кл. они расположены в периферических участках цпл. В сперматозоидах млекопитающих Мх сливаются, образуя спиральную гигантскую Мх, к-рая расположена в шейке.
 Мх в растительной клетке
Мх в растительной клетке



Спиральная Мх Мх в клетках почечных Мх в миоците
в шейке сперматозоида канальцев
В кл. почечного эпителия Мх расположены вдоль впячиваний цпм. Обычно мх скапливаются вблизи тех участков цпл, где возникает потребность в АТФ. В скелетных мышцах они нах-ся вблизи миофибрилл (мышечных волокон).
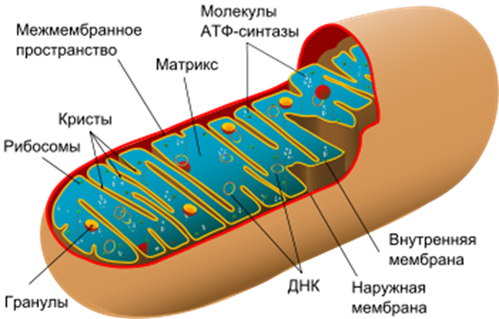
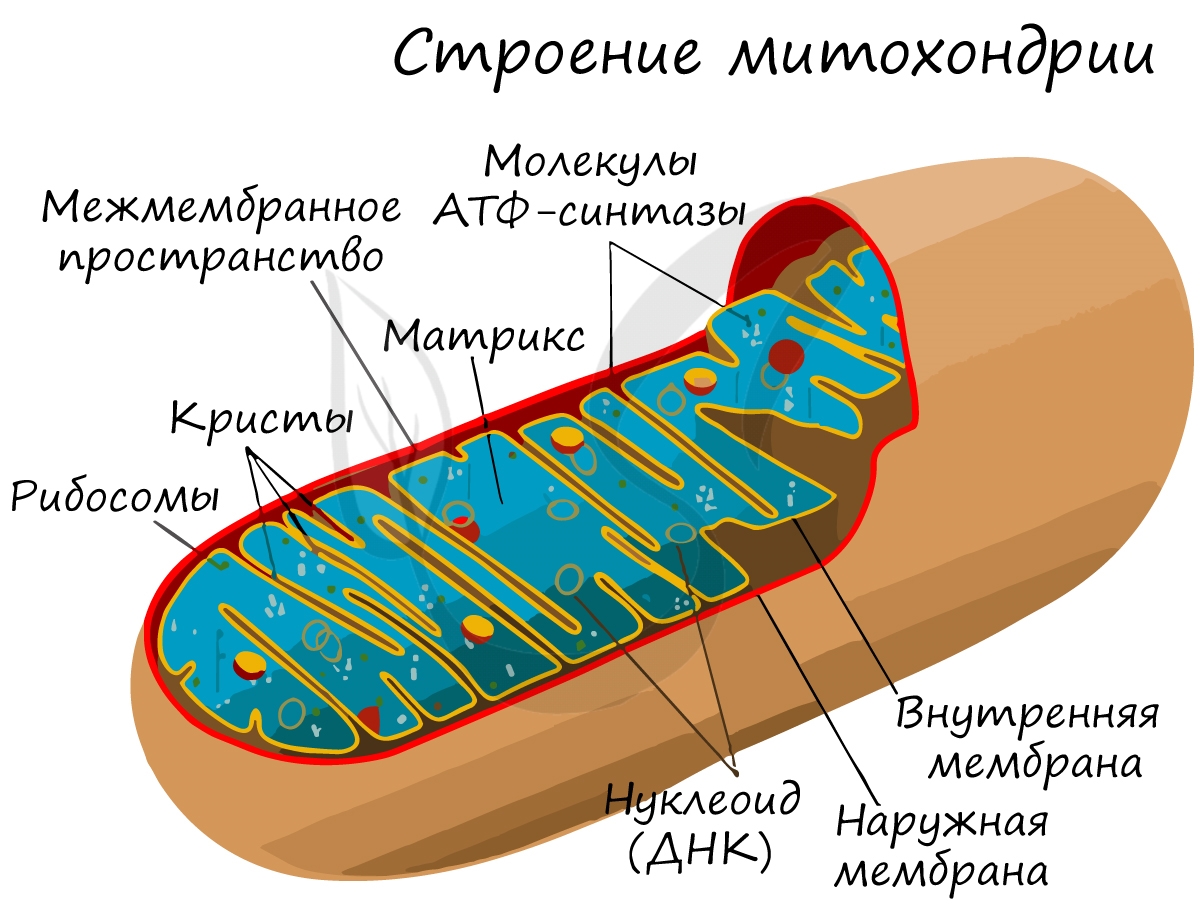
Под ЭМС Мх представляют собою чаще овальные тельца, стенка к-рых образована двумя элементарными мб: внешней и внутренней.

У митохондрий 2 мембраны, создающие 2 пространства: межмембранное и матричное. Межмембранное пространство создается между внешней и внутренней мембранами. Внешняя мб отделяет Мх от цитоплазмы и определяет ее форму. Внутренняя мб ограничивает матрикс, образуется матричное пространство. Впячивания внутренней мембраны в матричное пространство называются кристами (= гребнями).
Матрикс митохондрий – гомогенный коллоидный раствор, в к-ром растворены белки, углеводы и соли, а также нах-ся т-РНК, и-РНК, различные ферменты. В коллоидном р-ре матрикса расположены ДНК и рибосомы. Митохондриальные Рс имеют типичное строение и похожи на Рс бактерий (константы седиментации 50S, 30S и Σ 70S).
В матриксе митохондрий осуществляется биологическое окисление АК, ЖК и ПВК (пировиноградной кислоты) до СО2 и [Н], т.е. их диссимиляция. Эти процессы окисления сопровождаются отщеплением [H], т.е. осуществляются без участия кислорода. Реакции расщепления происходят с выделением энергии и сопряжены с эндотермическими реакциями синтеза АТФ, которые связаны с внутренней мембраной.
Внутренняя мембрана митохондрий содержит дыхательную цепь – цепь транспорта электронов (ЦТЭ). Во внутренней мембране происходит разделение зарядов [H] на Н + и ē. В результате транспорта электронов по дыхательной цепи происходит трансформация энергии в макроэргические связи АТФ.
 Во внутренней мембране находятся грибовидные тельца, содержащие фермент АТФ-синтетазу. Грибовидные тельца называют АТФ-сомами, так как они участвуют в синтезе АТФ.
Во внутренней мембране находятся грибовидные тельца, содержащие фермент АТФ-синтетазу. Грибовидные тельца называют АТФ-сомами, так как они участвуют в синтезе АТФ.
 |
2. Синтез АТФ. Энергия транспорта Н + через АТФ-синтетазу идет на синтез АТФ. Этот фермент катализирует р-цию синтеза АТФ:
 АДФ + Фн АТФ + Н2О
АДФ + Фн АТФ + Н2О
3. Автономный синтез Б, Ж и У.
4. Синтез ДНК. ДНК удваивается перед делением Мх.
Ученые предполагают, что Мх ведут свое начало от древних аэробных бактерий, некогда внедрившихся в предшественники эукариотических клеток. Доказательством тому м. служить следующие факты:
 — метаболические процессы связаны с мб
— метаболические процессы связаны с мб
— сходство рибосом: 50S и 30S
— сходный способ деления.
— сходный способ синтеза АТФ.
Встречаются у фотосинтезирующих эукариот:
 — высших растений (риниофиты – мхи – папоротники – Гс – Пс )
— высших растений (риниофиты – мхи – папоротники – Гс – Пс )

 — низших: водорослей
— низших: водорослей
протисты — простейших (эвглена, вольвокс) Бых, с. 16
Они способны превращаться др. в др. Лейкопласты – не содержат пигмента, хлоропласты содержат зеленый пигментхлорофилл, хромопласты содержат пигменты: красный фикоэритрин,оранжевый каротин, желтый ксантофилл, амилопласты – бесцветны, содержат крахмал.
1. Пожелтение и покраснение листьев осенью (Хпл ® Хрпл)

2. Изменение окраски листиков у лилейных (Хпл ® Хрпл или Лпл)




3. Позеленение корнеплода моркови (Хрпл ® Хпл)


сахарная свекла морковь турнепс брюква картофель
4. Позеленение клубней картофеля (Апл ® Хпл)



В растительных клетках Хпл имеют овальную форму. Фотосинтезирующие органоиды водорослей имеют различную форму (овальную, чашевидную и др.) и называются хроматофоры (хрома = краска, фор = несу). Кор. § 79-80, 202, 204.



Под СМС Хпл хорошо видны в неокрашенных клетках. В молодых кл. они расположены по всему объему цпл., в дифференцированных локализуются в пристеночном слое цпл.

 циклоз
циклоз
Хпл в цпл. движутся, увлекаемые током цитоплазмы. Движение цитоплазмы эукариотических клеток называется циклозом.
При изменении солнечного освещения Хпл меняют своё расположение в клетке. При очень интенсивном свете они перемещаются на вертикальные поверхности клеток, экранируя, таким образом, др. др. При недостаточном освещении они перемещаются на горизонтальные поверхности, чтобы улавливать больше лучей.








 с в е т о в о й п о т о к
с в е т о в о й п о т о к


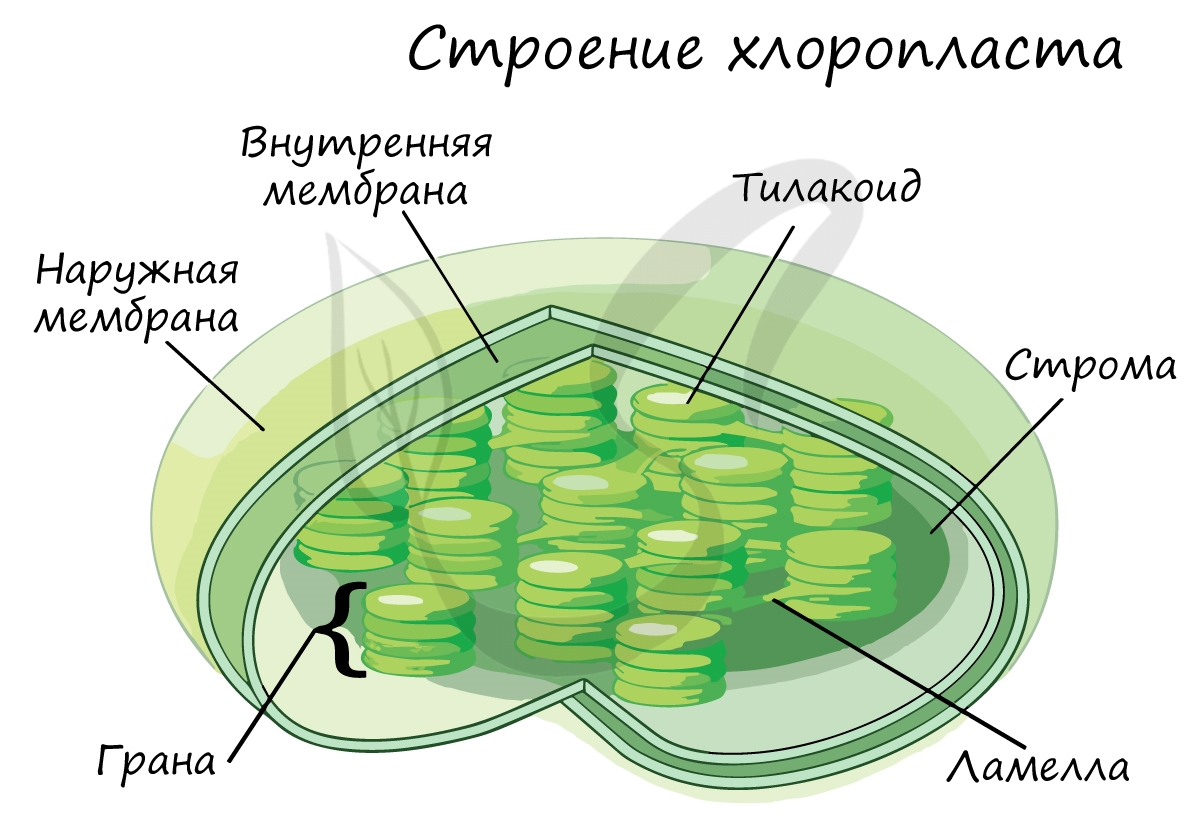
Под ЭМС хорошо различимо, что у Хпл овальная форма. Они покрыты двумя мембранами – наружной и внутренней.

В Хпл различают 3 мембраны:
 — наружную
— наружную
Внутренняя мб Хпл в отличии от Мх не образует впячиваний и не содержит в себе цепи переноса ē. ЦТЭ находится в тилакоидной мб.

3 мб делят Хпл на 3 внутренних отсека:
— межмембранный
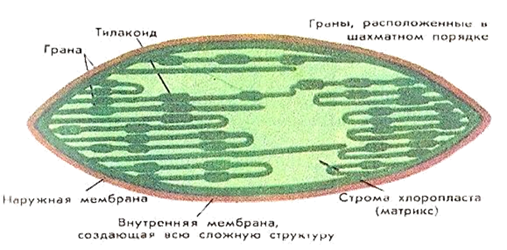
Межмембранное пространство возникает между внешней и внутренней мембранами. Внутренняя мембрана ограничивает строму, которая представляет собой коллоидный раствор. В стромальном пространстве находятся ДНК, имеющая кольцевидную форму, крахмальные зерна, рибосомы, и-РНК, т-РНК, ферменты и некоторые другие вещества.
 В строме расположены тилакоиды – плоские мембранные мешочки, стенки которых образованы третьими мембранами – тилакоидными. Одни тилакоиды расположены друг над другом, наподобие столбика монет, и образуют граны. Мешочки гран соединены друг с другом и образуют тилакоидное пространство. Число гран в Хпл может достигать 40 – 60.
В строме расположены тилакоиды – плоские мембранные мешочки, стенки которых образованы третьими мембранами – тилакоидными. Одни тилакоиды расположены друг над другом, наподобие столбика монет, и образуют граны. Мешочки гран соединены друг с другом и образуют тилакоидное пространство. Число гран в Хпл может достигать 40 – 60.
В грану помимо тилакоидных мешочков могут входить тилакоиды стромы. Тилакоиды стромы простираются через всю строму и не связаны друг с другом, они имеют удлиненную форму и не содержат хлорофилла.
Тилакоидная мембрана гран содержит зеленый пигмент – хлорофилл. Поглощенная энергия Солнца трансформируется в мембране тилакоида в энергию АТФ. Энергия Солнца идет на разделение зарядов [H] воды на Н + и ē.


На темновой стадии фотосинтеза энергия АТФ используется в реакциях ассимиляции, протекающих в строме.
1. Фотосинтез (FS). Световая стадия FS связана с гранами пластид, а темновая протекает в строме.
2. Синтез АТФ. Во время световой стадии FS происходит синтез АТФ.
3. Автономный синтез Б, Ж и У.
4. Запасная ф-ция. Накопление крахмала в виде зерен.
6.Деление пополам. Делятся путем перетяжки.



