Репликационная вилка

Репликационная вилка (репликативная вилка) — cтруктура, которая образуется во время репликации ДНК.
В подавляющем большинстве случаев (кроме некоторых вирусов и искусственно созданных молекул, содержащих одноцепочечную ДНК) макромолекула ДНК состоит из двух комплементарных цепей, имеющих противоположную направленность. Комплементарность цепей означает, что информационное содержание обеих цепей ДНК идентично. Разные концы цепочки ДНК называются 3’-конец и 5’- конец. Фермент ДНК-полимераза, осуществляющий удвоения цепочки нуклеотидов, может добавлять свободные нуклеотиды только к 3’-концу собираемой цепочки. Поэтому, для осуществления репликации обе цепи родительской ДНК должны быть отделены друг от друга (хотя бы на время).
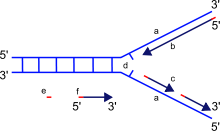
Четко ограниченная область, перемещающаяся вдоль родительской спирали ДНК, характеризующаяся местным расхождением двух ее цепей из-за своей Y-образной формы названа репликационной вилкой.
Ссылки
Полезное
Смотреть что такое «Репликационная вилка» в других словарях:
репликационная вилка — репликационная вилка. См. точка роста. (Источник: «Англо русский толковый словарь генетических терминов». Арефьев В.А., Лисовенко Л.А., Москва: Изд во ВНИРО, 1995 г.) … Молекулярная биология и генетика. Толковый словарь.
репликативная (репликационная) вилка — точка роста Молекулярная форма, образуемая материнской и двумя дочерними двухцепочечными молекулами ДНК в процессе полуконсервативной репликации. [Арефьев В.А., Лисовенко Л.А. Англо русский толковый словарь генетических терминов 1995 407с.]… … Справочник технического переводчика
Репликация ДНК — У этого термина существуют и другие значения, см. Репликация. Схематическое изображение процесса репликации, цифрами отмечены: (1) запаздыв … Википедия
Хеликаза — Структура геликазы RuvA кишечной палочки. Геликазы (хеликаза, helicase) (лат. helix «спираль») это класс ферментов, которые встречаются у всех живых организмов. Геликазы перемещаются по сахаро фосфатному остову нуклеи … Википедия
Лигаза — (лат. ligāre сшивать, соединять) фермент, катализирующий соединение двух молекул с образованием новой химической связи (лигирование[1][2]). При этом обычно происходит отщепление (гидролиз) небольшой химической группы от одной из… … Википедия
Репликация (биология) — Схематическое изображение процесса репликации, цифрами отмечены: (1) запаздывающая нить, (2) лидирующая нить, (3) ДНК полимераза (Polα), (4) ДНК лигаза, (5) РНК праймер, (6) ДНК праймаза, (7) фрагмент Оказаки, (8) ДНК полимераза (Polδ), (9)… … Википедия
ДНК-топоизомеразы — Топоизомераза I разрешает топологические проблемы, связанные со спирализацией деспирализацией ДНК. Топоизомераза I оборачивается вокруг ДНК и вносит разрыв, который позволяет спирали ДНК вращаться и снимает напряжение. После релаксации,… … Википедия
Хеликазы — Структура геликазы RuvA Escherichia coli Хеликазы (англ. helicase, от лат. helix спираль, по русски и … Википедия
DNA clamp — Белки скользящей застежки (DNA clamp) это белки, которые выполняют функцию усилителя процессивности при репликации ДНК. Белки DNA clamp являются важным компонентом холофермента ДНК полимеразы III и предотвращают диссоциацию фермента от… … Википедия
DnaB — Геликаза DnaB это бактериальный фермент, который открывает репликационную вилку во время репликации ДНК. Первоначально, когда DnaB связывается с DnaA, последний связан с DnaC, белком репрессором. После диссоциации DnaC, DnaB связывает DnaG.[1] У… … Википедия
Что такое вилка репликации?
Содержание:
В вилка репликации Это точка, в которой происходит репликация ДНК, ее также называют точкой роста. Он имеет форму буквы Y, и по мере репликации шпилька перемещается через молекулу ДНК.
Репликационная вилка формируется точно между стыком между недавно разделенной матрицей или цепями матрицы и дуплексной ДНК, которая еще не дублировалась. При инициации репликации ДНК одна из цепей может легко дублироваться, в то время как другая цепь сталкивается с проблемой полярности.
Репликация ДНК и вилка репликации
Этот огромный полимер, состоящий из четырех разных нуклеотидов (A, T, G и C), находится в ядре эукариот, в каждой из клеток, составляющих ткани этих существ (за исключением зрелых красных кровяных телец млекопитающих, в которых отсутствуют ядро).
Каждый раз, когда клетка делится, ДНК должна реплицироваться, чтобы создать дочернюю клетку с генетическим материалом.
Односторонняя и двусторонняя репликация
Репликация может быть однонаправленной или двунаправленной, в зависимости от формирования репликационной вилки в точке происхождения.
Логично, что в случае репликации в одном направлении образуется только одна шпилька, тогда как при двунаправленной репликации образуются две шпильки.
Вовлеченные ферменты
Для этого процесса необходим сложный ферментативный механизм, который работает быстро и может точно воспроизводить ДНК. Наиболее важными ферментами являются ДНК-полимераза, ДНК-примаза, ДНК-геликаза, ДНК-лигаза и топоизомераза.
Начало репликации и формирование шпильки
Репликация ДНК начинается не в каком-либо случайном месте молекулы. В ДНК есть определенные области, которые отмечают начало репликации.
Когда ДНК начинает открываться, образуется Y-образная структура: вилка репликации.
Удлинение и движение вилки
ДНК-полимераза не может начать синтез дочерних цепей с нуля. Вам нужна молекула с 3 ‘концом, чтобы полимераза могла начать полимеризацию.
Этот свободный 3′-конец предлагается небольшой нуклеотидной молекулой, называемой праймером или праймером. Первый действует как своего рода крючок для полимеразы.
В процессе репликации вилка репликации может перемещаться по ДНК. Прохождение репликационной вилки оставляет две однозонные молекулы ДНК, которые направляют образование двухзонных дочерних молекул.
Шпилька может продвигаться вперед благодаря действию ферментов геликазы, раскручивающих молекулу ДНК. Этот фермент разрывает водородные связи между парами оснований и позволяет перемещать шпильку.
Прекращение
Репликация прекращается, когда две шпильки находятся при температуре 180 ° C от источника.
В этом случае мы говорим о том, как протекает процесс репликации в бактериях, и необходимо выделить весь процесс кручения кольцевой молекулы, который подразумевает репликация. Топоизомеразы играют важную роль в раскручивании молекулы.
Репликация ДНК полуконсервативная
Вы когда-нибудь задумывались, как происходит репликация в ДНК? То есть из двойной спирали должна выйти еще одна двойная спираль, но как это происходит? В течение нескольких лет этот вопрос оставался открытым для биологов. Может быть несколько перестановок: две старые нити вместе и две новые нити вместе, или одна новая нить и одна старая, чтобы образовать двойную спираль.
В 1957 году на этот вопрос ответили исследователи Мэтью Мезельсон и Франклин Шталь. Предложенная авторами модель репликации была полуконсервативной.
Мезельсон и Шталь утверждали, что результатом репликации являются две молекулы двойной спирали ДНК. Каждая из полученных молекул состоит из старой цепи (исходной или исходной молекулы) и вновь синтезированной новой цепи.
Проблема полярности
Как работает полимераза?
Спираль ДНК состоит из двух антипараллельных цепей: одна идет в направлении 5′-3 ‘, а другая 3′-5’.
Наиболее важным ферментом в процессе репликации является ДНК-полимераза, которая отвечает за катализирование объединения новых нуклеотидов, которые будут добавлены в цепь. ДНК-полимераза может удлинять цепь только в направлении 5′-3 ‘. Это препятствует одновременному дублированию цепей в репликационной вилке.
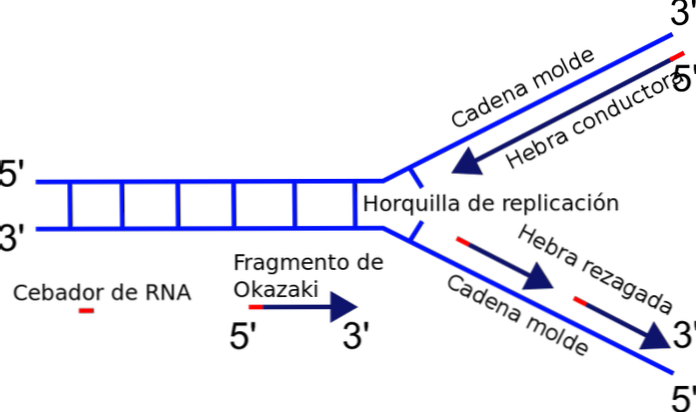
Зачем? Добавление нуклеотидов происходит на свободном конце 3 ‘, где есть гидроксильная группа (-ОН). Таким образом, только одна из цепей может быть легко амплифицирована добавлением терминального нуклеотида к 3’-концу. Это называется токопроводящей или непрерывной жилой.
Производство осколков Окадзаки
Этот прерывистый синтез фрагментов заканчивается объединением каждой из частей, реакцией, катализируемой ДНК-лигазой. В честь первооткрывателя этого механизма Рейджи Окадзаки небольшие синтезированные сегменты называются фрагментами Окадзаки.
Ссылки
Классическая древность: периоды и их характеристики
Что такое ветвь репликации?
вилка репликации это точка, в которой происходит репликация ДНК, она также называется точкой роста. Он имеет форму Y, и в процессе репликации шпилька смещается молекулой ДНК..
Вилка репликации формируется как раз между соединением между недавно отделенными цепочками матриц или матриц и дуплексной ДНК, которая еще не была продублирована. При инициации репликации ДНК одна из цепей может быть легко продублирована, в то время как другая цепочка сталкивается с проблемой полярности.
Репликация ДНК и репликационная вилка
Этот огромный полимер, состоящий из четырех различных нуклеотидов (A, T, G и C), находится в ядре эукариот, в каждой из клеток, которые составляют ткани этих существ (за исключением зрелых эритроцитов млекопитающих, у которых отсутствует ядра).
Каждый раз, когда клетка делится, ДНК должна реплицироваться, чтобы создать дочернюю клетку с генетическим материалом..
Однонаправленная и двунаправленная репликация
Репликация может быть однонаправленной или двунаправленной, в зависимости от формирования форка репликации в точке происхождения.
Вовлеченные ферменты
Для этого процесса необходим сложный ферментативный механизм, который работает быстро и может точно копировать ДНК. Наиболее важными ферментами являются ДНК-полимераза, ДНК-примаза, ДНК-геликаза, ДНК-лигаза и топоизомераза..
Начало репликации и формирования форка
Репликация ДНК не начинается в любом случайном месте в молекуле. Есть определенные области в ДНК, которые отмечают начало репликации.
Когда ДНК начинает открываться, образуется Y-образная структура: репликационная вилка.
Удлинение и движение вилки
ДНК-полимераза не может начать синтез дочерних цепей с нуля. Вам нужна молекула с 3′-концом, чтобы у полимеразы было место, где начать полимеризацию..
Этот свободный 3 ‘конец представлен небольшой молекулой нуклеотидов, называемой праймером или праймером. Первый действует как своего рода крючок для полимеразы.
С ходом репликации репликационная вилка способна перемещаться по ДНК. Шаг вилки репликации оставляет две однозонные молекулы ДНК, которые управляют образованием двухзонных дочерних молекул.
Вилка может продвигаться благодаря действию ферментов геликазы, которые раскручивают молекулу ДНК. Этот фермент разрушает водородные связи между парами оснований и позволяет перемещать вилку.
завершение
Репликация прекращается, когда две вилки находятся при 180 ° C от источника.
В этом случае мы говорим о том, как протекает процесс репликации в бактериях, и необходимо выделить весь процесс кручения кольцевой молекулы, который включает репликацию. Топоизомеразы играют важную роль в раскручивании молекулы.
Репликация ДНК является полуконсервативной
Задумывались ли вы, как репликация происходит в ДНК? То есть из двойной спирали должна возникать еще одна двойная спираль, но как это происходит? В течение нескольких лет этот вопрос оставался открытым для биологов. Может быть несколько перестановок: две старые нити вместе и две новые вместе или новая нить и старая, чтобы сформировать двойную спираль.
В 1957 году на этот вопрос ответили исследователи Мэтью Мезельсон и Франклин Шталь. Модель репликации, предложенная авторами, была полуконсервативной.
Мезельсон и Шталь утверждают, что результатом репликации являются две двухцепочечные молекулы ДНК. Каждая из полученных молекул состоит из старой цепи (от матери или исходной молекулы) и новой синтезированной новой цепи..
Проблема полярности
Как работает полимераза?
Спираль ДНК образована двумя цепочками, которые идут антипараллельно: одна идет в направлении 5′-3 ‘, а другая 3′-5’.
Наиболее важным ферментом в процессе репликации является ДНК-полимераза, которая отвечает за катализирование связывания новых нуклеотидов, которые будут добавлены в цепь. ДНК-полимераза может удлинять цепь только в направлении 5′-3 ‘. Этот факт препятствует одновременному дублированию цепочек в ветке репликации.
Почему? Добавление нуклеотидов происходит на свободном конце 3 ‘, где обнаружена гидроксильная группа (-ОН). Таким образом, только одна из цепей может быть легко амплифицирована путем терминального добавления нуклеотида к 3 ‘концу. Это называется проводящей или непрерывной жилой.
Изготовление фрагментов Оказаки
Другая цепь не может удлиняться, потому что свободный конец 5 ‘, а не 3’, и никакая полимераза не катализирует добавление нуклеотидов к 5 ‘концу. Задача решается путем синтеза нескольких коротких фрагментов (от 130 до 200 нуклеотидов), каждый в нормальном направлении репликации от 5 ‘до 3’.
Этот прерывистый синтез фрагментов заканчивается объединением каждой из частей, реакцией, катализируемой ДНК-лигазой. В честь исследователя этого механизма, Рейджи Окадзаки, синтезированные небольшие сегменты называются фрагментами Окадзаки..
Репликация ДНК
Репликация — это механизм самокопирования и основное свойство наследственного материала, которым выступают молекулы ДНК.
Особенностью ДНК является то, что обычно ее молекулы состоит из двух комплементарных друг другу цепей, образующих двойную спираль. В процессе репликации цепи материнской молекулы ДНК расходятся, и на каждой строится новая комплементарная цепь. В результате из одной двойной спирали образуется две, идентичные исходной. Т. е. из одной молекулы ДНК образуются две, идентичные матричной и между собой.
Таким образом, репликация ДНК происходит полуконсервативным способом, когда каждая дочерняя молекула содержит одну материнскую цепь и одну вновь синтезированную.
У эукариот репликация происходит в S-фазе интерфазы клеточного цикла.
Описанный ниже механизм и основные ферменты характерны для подавляющего большинства организмов. Однако бывают исключения, в основном среди бактерий и вирусов.
Расхождение цепей исходной молекулы ДНК обеспечивает фермент геликаза, или хеликаза, который в определенных местах хромосом разрывает водородные связи между азотистыми основаниями ДНК. Хеликазы перемещаются по ДНК с затратой энергии АТФ.
Чтобы цепочки снова не соединились, они удерживаются на расстоянии друг от друга дестабилизирующими белками. Белки выстраиваются в ряд со стороны пентозо-фосфатного остова цепи. В результате образуются зоны репликации, называемые репликационными вилками.
Репликационные вилки образуются не в любых местах ДНК, а только в точках начала репликации, состоящих из определенной последовательности нуклеотидов (около 300 штук). Такие места распознаются специальными белками, после чего образуется так называемый репликационный глаз, в котором расходятся две цепи ДНК.
Из точки начала репликация может идти как в одном, так и в двух направлениях по длине хромосомы. В последнем случае цепи ДНК расходятся вперед и назад, и из одного репликационного глазка образуются две репликационные вилки.
Репликон — единица репликации ДНК, от точки ее начала и до точки ее окончания.
Поскольку в ДНК цепи спирально закручены относительно друг друга, то разделение их хеликазой вызывает появление дополнительных витков перед репликационной вилкой. Чтобы снять напряжение, молекула ДНК должна была бы проворачиваться вокруг своей оси один раз на каждые 10 пар разошедшихся нуклеодидов, именно столько образуют один виток спирали. В таком случае ДНК бы быстро вращалась с затратой энергии. Но этого не происходит, т. к. природа нашла более эффективный способ справится с возникающим при репликации напряжением спирали.
Фермент топоизомераза разрывает одну из цепей ДНК. Отсоединенный участок проворачивается на 360° вокруг второй целой цепи и снова соединяется со своей цепью. Этим снимается напряжение, т. е. устраняются супервитки.
Каждая отдельная цепь ДНК старой молекулы используется в качестве матрицы для синтеза новой комплементарной себе цепи. Добавление нуклеотидов к растущей дочерней цепи обеспечивает фермент ДНК-полимераза. Существует несколько разновидностей полимераз.
В репликационной вилке к освободившимся водородным связям цепей согласно принципу комплиментарности присоединяются свободные нуклеотиды, находящиеся в нуклеоплазме. Присоединяющиеся нуклеотиды представляют собой дезоксирибонуклеозидтрифосфаты (дНТФ), а конкретно дАТФ, дГТФ, дЦТФ, дТТФ.
После образования водородных связей фермент ДНК-полимераза связывает нуклеотид фосфоэфирной связью с последним нуклеотидом синтезируемой дочерней цепи. При этом отделяется пирофосфат, включающий два остатка фосфорной кислоты, который потом расщепляется на отдельные фосфаты. Реакция отщепления пирофосфата в результате гидролиза энергетически выгодна, так как связь между первым, который уходит в цепь, и вторым фосфатными остатками богата энергией. Эта энергия используется полимеразой.
Полимераза не только удлиняет растущую цепь, но и способна отсоединять ошибочные нуклеотиды, т. е. обладает корректирующей способностью. Если последний нуклеотид, который должен быть присоединен к новой цепи, не комплементарен матричному, то полимераза его удалит.
Поскольку цепи ДНК антипараллельны, а синтез новой цепи возможен только в направлении 5´→3´, то в репликационной вилке дочерние цепи будут синтезироваться в разных направлениях.
На матрице 3´→5´ сборка новой полинуклеотидной последовательности происходит по большей части непрерывно, так как эта цепь синтезируется в направлении 5´→3´. Антипараллельная матрица характеризуется 5´→3´ направлением, поэтому синтез дочерней цепи по ходу движения вилки здесь не возможен. Здесь он был бы 3´→5´, но ДНК-полимера не может присоединять к 5´-концу.
Поэтому синтез на матрице 5´→3´ выполняется небольшими участками — фрагментами Оказаки (названы в честь открывшего их ученого). Каждый фрагмент синтезируется в обратном ходу образования вилки направлении, что обеспечивает соблюдение правила сборки от 5´- к 3´-концу.
После удаления праймеров и застраивания брешей ДНК-полимеразой отдельные участки дочерней цепи ДНК сшиваются между собой ферментом ДНК-лигазой.
Непрерывная сборка идет быстрее, чем фрагментарная. Поэтому одна из дочерних цепей ДНК называется лидирующей, или ведущей, вторая — запаздывающей, или отстающей.
У прокариот репликация протекает быстрее: примерно 1000 нуклеотидов в секунду. В то время как у эукариот только около 100 нуклеотидов. Количество нуклеотидов в каждом фрагменте Оказаки у эукариот составляет примерно до 200, у прокариот — до 2000.
У прокариот кольцевые молекулы ДНК представляют собой один репликон. У эукариот каждая хромосома может содержать множество репликонов. Поэтому синтез начинается в нескольких точках, одновременно или нет.
Ферменты и другие белки репликации действуют совместно, образуя комплекс и двигаясь по ДНК. Всего в процессе участвует около 20 разных белков, здесь были перечислены лишь основные.
Спивак Ирина Михайловна
Репликация ДНК
Глава 1. Репликация – полимеразная реакция
1.1. Вилка репликации
Рис. 1. Схема репликативной вилки.
Длина таких коротких фрагментов у прокариот составляет 1000–2000 пн. По имени открывшего их ученого они были названы «фрагментами Оказаки». По мере движения репликативной вилки концы соседних фрагментов Оказаки соединяются с образованием непрерывной отстающей нити. Для того, чтобы процесс на обеих нитях шел синхронно, полимеразные комплексы лидирующей и отстающей нити связаны между собой, образуя сложную трехмерную структуру (рис. 1, б)
Вилка репликации может двигаться как в одну сторону от точки начала репликации, так и в обе стороны. В зависимости от этого процесс называется однонаправленной или двунаправленной репликацией. Как это выглядит схематически, показано на рис. 2. У эукариот репликация обычно двунапраленная. Также и у E.coli.
Механизмы инициации репликации в точке начала репликации и при образовании фрагментов Оказаки в отстающей цепи в принципе аналогичны, хотя имеются некоторые тонкие различия. В обоих случаях происходит образование коротких РНК-затравок (праймеров), комплементарных матричной ДНК, в виде продолжения которых синтезируется новая цепь ДНК. В дальнейшем короткие вставки РНК замещаются сегментами ДНК, отдельные фрагменты Оказаки затем объединяются с образованием непрерывной отстающей нити.
Все живые организмы на Земле обычно делят на прокариот и эукариот (от греч. карион – ядро). Главной особенностью прокариот является отсутствие у них в отличие от эукариот полноценного клеточного ядра, покрытого оболочкой. Генетический материал прокариот расположен в нуклеоиде – примитивном эквиваленте ядра эукариот. Клетки прокариот имеют очень небольшие размеры – около 1 мкм. Объем эукариотических клеток в 800-1000 раз больше объема клеток прокариот. К прокариотам относятся бактерии и археи (или архебактерии), предки которых возникли около 4 млрд лет назад. Эукариоты могут быть как одноклеточными, так и многоклеточными. Они появились на Земле примерно через 500 млн лет после прокариот.
По современным представлениям ДНК-метаболизм у прокариот имеет некоторые отличия от такового у эукариот. Описывая процессы репликации и рекомбинации, мы будем каждый раз подчеркивать эти отличия.
Глава 2. Начало репликации
Рис. 2. Возможное движение репликативной вилки.
У про– и эукариот можно встретить различные вариации на эту тему. Так, каждая из цепей родительской спирали митохондриальной ДНК животных (15тпн) имеет свою точку начала репликации. Синтез комплементарной цепи некоторых небольших однонитевых фаговых геномов начинается вблизи одной специфической последовательности, а репликация полученного дуплекса может инициироваться совсем в другой точке. Репликация линейных двунитевых ДНК также инициируется в особых сайтах. Например, ДНК бактериофага Т7 (40тпн) реплицируется в двух противоположных направлениях к разным концам молекулы, начиная от одной точки, а каждая из двух цепей ДНК аденовируса человека (30–38 тпн) реплицируется последовательно всегда от З’-конца.
Для геномов эукариотических клеток характерно наличие множественных точек начала репликации, разбросанных по хромосоме на расстоянии около 20тпн. После инициации репликация продолжается в двух направлениях от каждой точки до тех пор, пока репликативные вилки двух соседних точек начала репликации не сольются. Полноразмерные ДНК каждой дочерней хромосомы получаются путем соединения более коротких, независимо инициированных новосинтезированных нитей.
2.1. Понятие о репликоне и ориджине репликации
2.2. Ориждин репликации E.coIi oriC
Наиболее подробно изучены ориджины у Е. соli и Bacillus subtilis. Область начала репликации хромосомы, оriС (origin of chromosome), включает в себя участки со специфическими последовательностями, так называемыми ДНК-боксами, и расположенными между ними короткими последовательностями. ДНК-боксы со специфическим «мотивом» нуклеотидов, преимущественно в 9пн, перемежаются фрагментами в 12-1Зпн с высоким содержанием АТ. Сами девятичленные последовательности могут располагаться как в прямом, так и в инвертированном положении по отношению друг к другу. Например, у В. subtilis имеется один фрагмент ТТАТССАСА и два других девятичленных бокса, ориентированных в противоположном направлении, с заменой одной из пар нуклеотидов. Всего у В. subtilis на оriС расположено 15 ДНК-боксов. Область оriС очень консервативна: ДНК-боксы сходного состава имеются в соответствующем месте хромосомы у других бактерий (только у Mycoplаsma genitalium, несмотря на наличие общих для всех бактерий ферментов репликации, ДНК-боксов найдено не было). Сами ДНК-боксы не кодируют белок или РНК, хотя между ними располагаются отдельные гены. Продукты этих генов также большей частью вовлечены в «обслуживание» процесса репликации ДНК.
Порядок расположения ДНК-боксов, промежуточных областей и их количество позволяют думать, что эволюционная дивергенция oriС шла главным образом за счет дупликаций и трипликаций. Схема абстрактного «минимального ориджина» прокариот представлена на рис. 3.
2.3. Ориджины других организмов
Рис. 4. Схема ориджина вируса SV40.
У дрожжей Saccharomyces cerevisiae особые последовательности, способные обеспечивать репликацию фрагментов ДНК в дрожжевой клетке были выделены раньше, чем у других эукариот. Позднее такие последовательности были найдены и у многих других организмов. У S.cerevisiae АRS занимает 100—200пн и содержит специфическую консенсусную последовательность (АСS – ARS consensus sequence), размером в 11пн, необходимую для связывания с белком-инициатором, а также дополнительные элементы (В-элементы), усиливающие функцию ориджина. Например, АRS1 – первый подробно охарактеризованный ориджин – содержит три таких элемента – В1, В2, ВЗ. Последовательности АCS и В1 занимают приблизительно 50пн и представляют собой наименьшую функциональную область любого ориджина, которая требуется для связывания с белком-инициатором.
Элемент В2 обычно содержит генетически охарактеризованный участок DUE. Вспомогательный элемент ВЗ связывает фактор транскрипции Abf-1. Общая длина ARS-элемента составляет 100-200пн. Строение ориждина S.cerevisiae представлено на рис. 5.
Рис. 5. Схема ориджина Saccharomyces cereiseae
У другого вида дрожжей, Shizosaccharomyces pombe, ориджины состоят по крайней мере из одной ARS, которая значительно длиннее, чем у S. cerevisiae. В некоторых случаях несколько ARS-элементов формируют зону инициации репликации. (Рис. 6.)
Рис. 6. Схема ориджина Shizosaccharomyces pombe
У млекопитающих ориджины детально не охарактеризованы, некоторые из них располагаются в межгенных промежутках, имеют сайты связывания для транскрипционных факторов, часто содержат только районы инициации двунаправленной репликации – OBR.
2.4. Скорость репликации
Скорость репликации генома регулируется в основном частотой инициирующих событий. Так, у Е. соli скорость копирования в каждой репликативной вилке постоянна и равна примерно 1500пн в секунду: следовательно, полный геном длиной 4·10 6 пн реплицируется примерно за 40 мин. Если хромосома реплицируется быстрее, это значит, что увеличивается частота актов инициации в той же самой точке начала репликации при прежней скорости копирования. Клетки Е. соli делятся каждые 20 мин; это означает, что репликация ДНК инициируется в хромосомах, еще не закончивших предыдущий раунд репликации. Скорость движения репликативной вилки в эукариотических клетках значительно меньше (10-100пн в секунду), но завершение репликации хромосомы в разумное время обеспечивается одновременной инициацией во множестве точек. Итак, скорость репликации хромосом контролируется числом и расположением точек начала репликации. Например, в ранних эмбрионах дрозофилы репликация отдельной хромосомы осуществляется каждые 3 мин, благодаря почти одновременной инициации событий в точках, отстоящих друг от друга на 7000-8000пн. В тоже время известно, что у дрозофилы в ходе раннего эмбрионального развития, как скорость репликации, так и размеры и число репликонов тканеспецифичны. В культуре же соматических клеток той же дрозофилы скорость удвоения хромосом значительно более медленная, так как репликация начинается в гораздо меньшем числе точек, находящихся друг от друга на расстоянии 40000пн, при этом продолжительность S-фазы составляет 600 мин. Следовательно, при фиксированной скорости синтеза ДНК множественная инициация повышает скорость процесса репликации в целом и таким образом уменьшает время, необходимое для удвоения всего набора хромосом. Данные о числе репликонов и скорости репликации приведены в табл.1.
Различия в продолжительности S-фазы найдены и у других организмов. Например, у тритона S-фаза длится 1 ч в ядрах бластулы и 200 ч в предмейотической S-фазе сперматоцитов. Вероятно, длительность S-фазы определяется не скоростью синтеза ДНК, а числом задействованных ориджинов репликации. В ДНК клеток нейрулы тритона они находятся на расстоянии около 40 мкм друг от друга, а в соматических клетках – около 100 мкм.
Таблица 1
Число и длина репликонов у разных организмов.



