Рестриктазы
Эндонуклеазы рестрикции (или рестриктазы, лат. restrictio — ограничение) — группа ферментов, относящихся к классу гидролаз, катализируют реакцию гидролиза нуклеиновых кислот.
В отличие от экзонуклеаз, рестриктазы расщепляют нуклеиновые кислоты не с конца молекулы, а в середине. При этом каждая рестриктаза узнаёт определённый участок ДНК длиной от четырёх пар нуклеотидов и расщепляет нуклеотидную цепь внутри участка узнавания или вне его.
Содержание
Классификация
Выделают три класса эндонуклеаз рестрикции. К первому классу принадлежат ферменты (например, ЕсоК из Escherichia coli К12), узнающие специфическую последовательность сайта, но разрывающие нить ДНК в произвольной точке (по-видимому, после образования комплекса с ДНК фермент неспецифически взаимодействует с удаленной областью ДНК или передвигается вдоль нити ДНК). Ко второму классу принадлежат ферменты (например, EcoRI), расщепляющие ДНК в строго определенной точке по отношению к сайту узнавания. К третьему классу относят ферменты промежуточного типа (напр., EcoPI), разрывающие нить ДНК в нескольких точках на разном удалении от сайта узнавания. При этом образуются фрагменты ДНК либо с ровными (тупыми) концами, либо с выступающими (липкими) 5′- или З’-концами
Значение
Рестриктазы — часть сложной системы рестрикции-модификации, используемой бактериальными клетками для регуляции содержания и активности ДНК в клетке.
Открытие рестриктаз в 1970-х годах вместе с разработкой способов секвенирования ДНК послужило основным толчком для развития генетической инженерии.
Изошизомеры
Изошизомеры — это пары эндонуклеаз рестрикции, имеющих специфичность к распознаванию одинаковых последовательностей, но иногда отличающихся по наличию метилированных нуклеотидных остатков, и разрезающих эти последовательности в одинаковых местах. Например, изошизомерами являются рестриктазы Sph I (CGTAC^G) и Bbu I (CGTAC^G). Первый выделенный фермент для узнавания и специфического разрезания заданной последовательности, называют прототипом, а все остальные подобные рестриктазы называют изошизомерами.
Изошизомеры выделяют из разных штаммов бактерий и поэтому разные изошизомеры могут требовать разных условий реакции.
Гетерошизомеры (неошизомеры)
Фермент, узнающий такую же последовательность, но разрезающий ее по-другому, называют гетерошизомером (неошизомером). Изошизомеры, таким образом, являются частным случаем гетерошизомеров. Например, рестриктазы Sma I (GGG^CCC) и Xma I (G^GGCCC) являются гетерошизомерами, но не изошизомерами друг для друга.
Изокаудомеры
Рестриктазы, разпознающие совершенно разные последовательности, но образующие одинаковые концы, называют изокаудомерами.
См. также
Полезное
Смотреть что такое «Рестриктазы» в других словарях:
РЕСТРИКТАЗЫ — (ферменты рестрикции) группа бактериальных нуклеаз, специфически расщепляющих ДНК. Предохраняют бактериальные клетки от чужеродных ДНК (напр., вирусных). Широко используются при определении первичной структуры ДНК, для картирования генов и в… … Большой Энциклопедический словарь
рестриктазы — ферменты, разрезающие ДНК по определенным нуклеотидным последовательностям (сайтам рестрикции). Открыты в конце 60–х гг. XX в. как агенты, элиминирующие чужеродные нуклеиновые кислоты (вирусов) в клетках бактерий. В дальнейшем было показано, что… … Словарь микробиологии
Рестриктазы — * рэстрыктазы * restrictases ферменты рестрикции (см.), разрезающие ДНК по определенным нуклеотидным последовательностям, называемым сайтами рестрикции (см.). Р. кодируются бактериями, плазмидами и бактериофагами. Широко используются в генной… … Генетика. Энциклопедический словарь
рестриктазы — (ферменты, рестрикции), группа бактериальных нуклеаз, специфически расщепляющих ДНК. Предохраняют бактериальные клетки от чужеродных ДНК (например, вирусных). Широко используются при определении первичной структуры ДНК, для картирования генов и в … Энциклопедический словарь
РЕСТРИКТАЗЫ — (эндодезоксирибонуклеазы рестрикции), ферменты класса гидролаз, катализирующие гидролиз фос фодиэфирных связей чужеродных ДНК в большинстве про кариотических (бактерии и синезеленые водоросли) и нек рых др. организмах и выполняющие тем самым… … Химическая энциклопедия
РЕСТРИКТАЗЫ — (ферменты, рестрикции), группа бактериальных нуклеаз, специфически расщепляющих ДНК. Предохраняют бактериальные клетки от чужеродных ДНК (напр., вирусных). Широко используются при определении первичной структуры ДНК, для картирования генов и в… … Естествознание. Энциклопедический словарь
Рестриктазы — ферменты бактериального происхождения, распознающие специфические нуклеотидные последовательности длиной от 4 до 10 пар нуклеотидов и разрезающие молекулу ДНК в этом месте … Словарь по психогенетике
Рестрикционные эндонуклеазы рестриктазы р ферменты — Рестрикционные эндонуклеазы, рестриктазы, р. ферменты * рэстрыкцыйныя эндануклеазы, рэстрыктазы, р. ферменты * restriction endonucleases or restriction enzymes любой бактериальный фермент, узнающий определенные нуклеотидные последовательности… … Генетика. Энциклопедический словарь
Эндонуклеазы рестрикции рестриктазы — Эндонуклеазы рестрикции, рестриктазы * эндануклеазы рэстрыкцыі, рэстрыктазы * restriction еndonucleases ферменты, которые узнают специфические последовательности для разрезания (сайты узнавания, см.) и обычно состоят из 4 7 п. о. Бактериальная… … Генетика. Энциклопедический словарь
РЕСТРИКТИРУЮЩИЕ ФЕРМЕНТЫ (РЕСТРИКТАЗЫ) — Эндонуклеазы, распознающие специфическую последовательность нуклеотидов в ДНК и осуществляющие затем двухцепочный разрез ее в молекуле. Применяются для рестрикции ДНК с целью анализа полиморфизма отдельных участков ДНК … Термины и определения, используемые в селекции, генетике и воспроизводстве сельскохозяйственных животных
Рестриктазы




![]()
![]()
ЛПЗ № 1. Ферменты генетической инженерии.
Рестриктазы были открыты при изучении явления рестрикции и модификации у бактерий, расшифрованного в основном В. Арбером (1968). Рестриктазы узнают определенные последовательности нуклеотидов и разрезают двунитевую ДНК на фрагменты. Модификация заключается в метилировании определенных оснований в последовательности, узнаваемой сопряженной рестриктазой; тем самым обеспечивается защита данного участка ДНК от воздействия рестриктазы. Одновременное наличие в клетке этих двух ферментативных активностей (так называемая R-M система) препятствует гидролизу собственной нуклеиновой кислоты. Чужеродная же ДНК при проникновении в бактериальную клетку служит субстратом для обоих ферментов.
Первоначально многие считали, что единственной функцией R-M систем является защита клеток от инфицирования фагами. Однако дальнейшие исследования позволили сделать предположение о том, что R-M системы осуществляют функцию ограничения скрещивания между различными бактериальными видами и штаммами, которая, однако, не абсолютна и позволяет части чужеродной ДНК проникать в клетку, рекомбинационно встраиваться и поддерживаться в качестве генетического фонда для получения эволюционного преимущества. Уместно заметить, что у бактерий весьма проблематично определение вида. Существуют даже предположения об общем генофонде всех микроорганизмов, что должно было бы привести к бесконечному появлению новых видов бактерий во времени. Реально же мы видим, что бактерии проявляют определенное постоянство морфологических, генетических и биохимических характеристик: Достойными кандидатами для обеспечения в эволюции относительной стабильности генетического материала, т. е. для осуществления генетической изоляции, не отрицающей обмена определенными блоками, являются системы рестрикции-модификации.
Системы рестрикции и модификации найдены практически у всех исследованных бактерий. Недавно рестриктазы обнаружены и у некоторых видов дрожжей.
В 1968 г. М. Мезельсон и Р. Юань сообщили о выделении первой рестриктазы из штамма Е. coli К12. Подобный фермент был получен и из штамма Е. coli В. Данные эндонуклеазы ЕсоК и ЕсоВ отличались высокой специфичностью по отношению к узнаваемой последовательности нуклеотидов, yо расщепляли молекулы ДНК неспецифическив другом месте, отстоящем от участка (сайта) узнавания. В 1970 г. X. Смит иК. Вилькокс выделили из Haemophius influenzae рестриктазу HindII, не только специфически узнающую, но испецифически расщепляющую молекулы ДНК. При гидролизе вирусной или плазмидной ДНК рестриктазами такого типа образуется строго определенный набор фрагментов. Это наглядно выявляется при электрофоретическом разделении смеси получающихся фрагментов.
Принципиальное значение для разработки методологии генетической инженерии имело открытие в 1971 г. Р. Ёшимори рестриктаз EcoR I и EcoR II. Спомощью первой из них удалось выполнить пионерскую работу по направленной реконструкции генетического материала in vitro. В настоящее время рестриктазы используют практическиво всех генно-инженерных экспериментах. Такое широкое применение ферментов данного типа обусловлено их высокой специфичностью, а также особенностями структуры концов фрагментов ДНК, образуемых рестриктазами. Общепринято термины рестриктаза, эндонуклеаза рестрикции, сайтспецифическая эндодезоксирибонуклеаза считать синонимами.
X. Смит и Д. Натане в 1973 г. предложили номенклатуру рестриктаз, которая включает следующие пункты:
3.Различные системы рестрикции-модификации, кодируемые одной и той же бактериальной клеткой обозначаются римскими цифрами, например: HindI, HindII, HindIII.

4. Ферменты рестрикции-модификации в общем виде обозначаются как эндонуклеаза R или метилаза М с последующим определением
названия системы, например: эндонуклеаза R * HindII или метилаза М * HindII.
5. Если система генетически локализована в геноме фага или на плазмиде, то после родо-видового названия указывается символ внехромосомного элемента: EcoРI, EcoKII. Штаммовая принадлежность в этих случаях указывается в скобках: Есо(К)РI.
Открытие большого числа рестриктаз и изучение их свойств позволило выявить некоторые закономерности функционирования ферментов и разделить их на три класса. Основой классификации служат в первую очередь потребность фермента в кофакторах и характер расщепления ДНК.
Различают 3 основных класса рестриктаз: 1, 2 и 3. Все рестриктазы узнают на двуспиральной ДНК строго определенные последовательности, но рестриктазы 1-го класса осуществляют разрывы в произвольных точках молекулы ДНК, а рестриктазы 2-го и 3-го классов узнают и расщепляют ДНК в строго определенных точках внутри сайтов узнавания или на фиксированном от них расстоянии.
Ферменты второго класса состоят из 2 отдельных белков: рестрицирующей эндонуклеазы и модифицирующей метилазы, поэтому в генной инженерии используются исключительно ферменты 2-го класса. Они нуждаются в ионах магния в качестве кофакторов.
Рестриктазы I класса, к которым относятся, к примеру, ферменты ЕсоК, EcoB, EcoPl и другие, атакуют ДНК в произвольных местах и образуют сплошной спектр рестриктов. Эти рестриктазы в генетической инженерии не используются, поскольку с их помощью нельзя получить фрагменты ДНК строго детерминированных размеров.
Рестриктазы класса II. Системы рестрикции-модификации класса II состоят из отдельных белков рестрикционной эндонуклеазы и модификационной метилазы. Поэтому рестриктазы данного класса можно выделить в индивидуальном состоянии, свободном от метилазной активности, что в значительной мере упрощает их изучение и последующее использование для расщепления молекул ДНК.
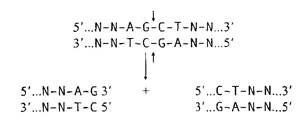
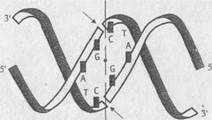
Рис. Строение участка расщепления молекулы ДНК рестриктазой AluI.
Кроме выступающих 5′-концов рестриктазы могут образовывать также липкие 3′-концы. К подобным ферментам относится рестриктаза PstI:
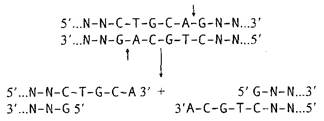
Ряд рестриктаз узнают частично вырожденные последовательности, поэтому такие ферменты не обладают столь высокой специфичностью, как рассмотренные выше.
Большой интерес для экспериментов по клонированию фрагментов ДНК представляют рестриктазы, которые, обладая разной специфичностью, дают при гидролизе ДНК одинаковые липкие концы.
Целенаправленный поиск эндонуклеаз рестрикции класса II, обусловленный значением этих ферментов, привел к открытию большого числа новых рестриктаз Новой считается любая рестриктаза найденная в не изученном ранее штамме. Дальнейшие исследования позволяют решить, действительно ли данный фермент узнает новую, последовательность нуклеотидов или же является аналогом уже известной рестриктазы. В том случае, если обнаруженная рестриктаза узнает ране неизвестную последовательность, она называется прототипам. Однако часто ферменты, выделенные из различных микроорганизмов, узнают одну и ту же последовательность и при гидролизеопределенной ДНК образуют одинаковый спектр фрагментов. Такие рестриктазы называют изошизомерами.В то же время, узнавая одну и ту же последовательность, рестриктазы могут по-разному расщеплять двухценочечную ДНК. Поэтому ферменты, имеющие одинаковыеузнаваемые последовательности и одинаково их расщепляющие, принято называть истинными изошизомерамн. Например, рестриктазы HapII, HpaII, MnoI являются истинными изошизомерами.Рестриктазы же SmaI и ХтаI, а также RsaI и CviIIимеют попарно одинаковые узнаваемые последовательности, но расщепляют ДНК различным образом. Такие ферменты принято называть ложными изошизомерами.
Распространенность рестриктаз.Рестриктазы необычайно широко распространены в мире микроорганизмов. Показано, что рестрикция и модификация не коррелируют с патогенностью. Не обнаружено и зависимости от потребления кислорода, т. е. R-M системы имеются и в аэробах, и в анаэробах. Строение клеточной стенки также не накладывает ограничений на присутствие систем рестрикции-модификации. Необходимо подчеркнуть, что рестриктазы и метилазы не являются обязательными компонентами клетки. Штаммы, не содержащие R-M систем, иногда называют «нулевыми».
Распространенность рестриктаз в царстве прокариот поставила вопрос о возможности существования R-M систем в эукариотических клетках. Попытки обнаружить у эукариот ферменты, аналогичные рестриктазам класса II, долгое время не давали положительных результатов. В середине 1980-х гг. появились сообщения о выделении специфических эндонуклеаз из Saccharomyces cerevisiae, Pichia membranafaciens, Chlamydomonas reinhardtii. Однако данные ферменты не давали исчерпывающего гидролиза ДНК, чем напоминали рестриктазы класса III.
Современные методы бионанотехнологии. Демонстрационный
Лекция 1.2 Основы генетической инженерии
Название раздела молекулярной генетики, именуемого генетической инженерией, указывает на то, что в результате такого рода исследований создаются искусственные генетические конструкции, в которых отдельные части генов или гены целиком объединяются в требуемой последовательности руками экспериментатора – генного инженера. Это позволяет определять их взаимное влияние и функциональное значение, а также проводить экспрессию генов в новом генетическом окружении. Таким образом, в экспериментальных условиях имеет место обмен генетической информацией как между отдельными генами организмов одного и того же вида, так и между организмами разных таксономических групп, что не происходит в природе из-за непреодолимых барьеров репродуктивной изоляции. Тем не менее, практически во всех методах современной генной инженерии фрагментарно (в адаптированном виде) используются элементы природных молекулярно-генетических механизмов.
Обмен генетической информацией между носителями этой информации – живыми организмами, а также составляющими их соматическими и половыми клетками является фундаментальным принципом существования всего живого. Половой процесс освобождает организмы от груза необратимых изменений в виде соматических мутаций и других многочисленных модификаций макромолекул, которые нарушают его нормальное функционирование в старости. Кроме того, в результате такого процесса геном нового организма воссоздается в новом сочетании аллелей, что, как правило, сопровождается расширением его адаптивных возможностей. Этот широко распространенный способ обмена генами между организмами с их передачей по вертикали, т.е. от поколения к поколению, не исчерпывает всего феномена обмена генами, непрерывно происходящего в биосфере.
Известна обширная группа генетических явлений, связанная с горизонтальной передачей генов в пределах одного поколения организмов, а также между клетками одного и того же многоклеточного организма. Характерными примерами такого рода являются специфическая и неспецифическая трансдукции, осуществляемые бактериофагами, в результате чего происходит перенос небольших частей генома микроорганизмов. Большое значение в эволюции бактерий играет и обмен генами с помощью конъюгативных плазмид и транспозонов, в частности распространение генов устойчивости к различным химическим веществам как в популяциях родственных бактерий, так и между представителями таксономически удаленных друг от друга групп. Ретровирусы, по-видимому, и в природных условиях способны осуществлять горизонтальный перенос генов у млекопитающих, а с помощью Ti-плазмид происходит горизонтальный обмен генами и у растений. Перемещение генетической информации и изменение характера ее экспрессии возможны и в пределах самих одноклеточных и многоклеточных организмов под действием разнообразных мобильных генетических элементов, а также при воздействии мутагенных факторов окружающей среды.
Приведенные примеры показывают, что в природе обмен блоками генов, отдельными генами и их фрагментами как в пределах геномов, так и между различными геномами и организмами – обычное явление. В результате этих событий переносимые гены не только сохраняют свою способность к экспрессии в новом генетическом окружении, но и могут значительно менять ее уровень, что часто сопровождается характерным изменением фенотипа организмов.
Образование новых сочетаний генов и их частей в природных условиях, по-видимому, не носит целенаправленного характера, и лишь жесткая проверка естественным отбором может оценить жизненную значимость таких преобразований геномов. Однако сознательное использование в лабораторных условиях основных генетических принципов, лежащих в основе природных перемещений генов, позволило разработать более эффективные системы передачи генетической информации между организмами и приступить к беспрецедентным по информативности исследованиям генетических явлений на молекулярном уровне. У нас на глазах произошло рождение нового направления в молекулярной биологии – генной инженерии, значение которого не ограничивается результатами тех или иных фундаментальных и прикладных исследований. Несомненно, именно с этого момента начался новый этап эволюции биосферы Земли, все последствия, которого мы в настоящее время не в состоянии предвидеть.
Необходимость манипулирования генами диктуется конкретными задачами фундаментальных и прикладных исследований. Для понимания молекулярных механизмов функционирования отдельных генов и взаимосвязанных генетических систем большое значение имеет работа с изолированными генами. Такие исследования позволяют определить границы генов, выделить их в чистом виде и идентифицировать элементы структуры, существенные для функционирования. Доказательством функциональной значимости выделенного участка генома может быть только его нормальная экспрессия в модельной генетической системе. Поэтому следующим этапом исследования выделенного гена всегда является перемещение его в такую генетическую систему, где экспрессия гена легко обнаруживается. Результаты экспрессии оценивают либо по появлению белкового продукта, кодируемого исследуемым геном, либо по изменению функций биологической системы вследствие появления в ней новой ферментативной или другой активности, например по компенсации присутствующей в этой системе мутации. Таким образом, в результате исследования структуры конкретного гена и моделирования его экспрессии в искусственной генетической системе можно понять особенности его функционирования в живом организме. Подобный подход может быть успешно применен как к известным генам, которые выделяются целенаправленно, так и к неидентифицированным ранее последовательностям нуклеотидов, функциональную значимость которых определяют лишь после выделения их в чистом виде. Последний подход реализуется в так называемой обратной генетике.
В настоящее время с помощью методов генной инженерии получены данные о структуре и функционировании генов разнообразных организмов, что дало возможность перейти на качественно новый уровень генетических исследований. Это, во-первых, возможность переноса гена в новое для него генетическое окружение с дальнейшей его экспрессией, что ведет к изменению свойств организма, в геном которого вводится ген (например создание продуцентов биологически активных веществ или трансгенных животных), а также осуществление генотерапии наследственных и приобретенных заболеваний путем искусственного замещения мутантных аллелей. Во-вторых, стало реальным конструирование новых генов путем объединения in vitro как известных, так и новых, искусственно синтезированных последовательностей нуклеотидов. Этот подход используется в белковой инженерии для исследования функциональной значимости отдельных аминокислот и доменов в полипептидных цепях ферментов, а также для создания новых белков. В-третьих, в современной биотехнологии появилась возможность применять изолированные гены в составе генно-инженерных конструкций для получения пищевых продуктов и биологически активных веществ белковой природы.
Поскольку в экспериментальных условиях невозможно работать с одной копией гена, получение необходимого числа идентичных копий гена или его частей является первой и одной из основных задач генной инженерии. Для ее решения используют метод молекулярного клонирования. Сущность метода заключается в том, что нуклеотидная последовательность, которую необходимо выделить или размножить, ковалентно встраивается в самореплицирующиеся молекулы нуклеиновой кислоты, называемые векторами. Далее такая последовательность нуклеотидов в составе вектора вводится в клетки про- или эукариотического организма, и эти гибридные клетки в селективных условиях, обеспечивающих сохранение вектора внутри клеток, выращивают на питательной среде. В результате образуется клон клеток, теоретически содержащих идентичные векторные молекулы с одной и той же вставкой чужеродной последовательности нуклеотидов. Поскольку объединение молекул клонируемой последовательности нуклеотидов и вектора является не чем иным, как рекомбинацией in vitro, такие гибридные молекулы называют рекомбинантными молекулами. В настоящее время разработаны многочисленные методы, позволяющие выделять определенные последовательности нуклеотидов из сложной смеси фрагментов хромосомной ДНК, а также осуществлять обмен между строго определенными фрагментами генов и другими последовательностями нуклеиновых кислот. Во всех этих реакциях, как правило, используются высокоочищенные препараты нуклеиновых кислот и ферментов нуклеинового обмена, особенности использования которых будут кратко рассмотрены ниже.
Большинство ферментов, применяемых для молекулярного клонирования нуклеиновых кислот, участвует в метаболизме нуклеиновых кислот in vivo. Это означает, что генная инженерия в своем развитии опирается на достижения исследований ферментных систем метаболизма нуклеиновых кислот и существует благодаря возможности получения таких ферментов в высокоочищенном состоянии. В то же время сам процесс клонирования и исследования клонированных последовательностей нуклеотидов сводится в основном к последовательному проведению in vitro определенных ферментативных реакций с использованием очищенных ферментов и их субстратов – нуклеиновых кислот.
При очистке ДНК и РНК в основном используются одни и те же приемы. Различия в методах их фракционирования определяются значительно большей чувствительностью молекул РНК к гидролитическому расщеплению из-за присутствия у рибонуклеотидов, входящих в состав РНК, свободных 2′-ОН групп, их более низкой молекулярной массой и наличием у одноцепочечных молекул характерной (компактной) пространственной структуры. Для выделения нуклеиновых кислот в нативном состоянии необходимо соблюдать, по крайней мере, две предосторожности: следует использовать мягкие условия разрушения тканей биологического объекта, содержащего изучаемые нуклеиновые кислоты, и инактивировать гидролитические ферменты (ДНКазы или РНКазы) до того, как они успеют гидролизовать молекулы нуклеиновых кислот. С учетом этого ингибиторы нуклеаз вводятся в буферные растворы до разрушения клеток или тканей. В качестве ингибиторов нуклеаз используют как неспецифические денатурирующие агенты (ионные детергенты, гидроокись ртути, гуанидинхлорид, гуанидинизотиоцианат, фенол, хлороформ и т.п.), так и специфические ингибиторы, например ингибитор РНКазы из плаценты человека или ванадиевые комплексы рибонуклеозидов.
Работу двухцепочечной геномной ДНК (особенно ДНК эукариотических клеток) сильно усложняют значительная молекулярная масса и высокая жесткость выделяемых молекул. Следствием этого являются большая вязкость растворов высокомолекулярной ДНК и легкость ее фрагментации в растворах. В связи с этим при выделении высокомолекулярной ДНК допускается лишь мягкое перемешивание ее раствора в процессе депротеинизации (освобождения от белков). Для этого на первых этапах очистки обычно применяют протеолитические ферменты (протеиназу К из Trirachium album или проназу из Streptomyces griseus), фенол и хлороформ. Поскольку при освобождении от белков происходит одновременная очистка ДНК и РНК, для освобождения от примесей РНК применяют высокоочищенные препараты РНКаз без примесей ДНКаз (чаще всего панкреатическую РНКазу), а в случае выделения РНК – ДНКаз, не загрязненных РНКазами.
Рестриктазы. Биологическая роль системы рестрикции-модификации бактерий. Виды рестриктаз. Рестриктазы II класса: особенности их строения и функций. Палиндромы. Классификация рестриктаз II класса: изошизомеры, неошизомеры, изокаудомеры. Образование фрагментов ДНК с «тупыми» и «липкими» концами.
Среди ферментов, используемых в генной инженерии для клонирования, большое значение имеют эндонуклеазы рестрикции – рестриктазы. Эти ферменты, впервые открытые как часть системы рестрикции–модификации ДНК у бактерий, специфически гидролизуют молекулы двухцепочечных ДНК при наличии в них определенных последовательностей нуклеотидов, называемых сайтами рестрикции. В то же время метилазы используют для ограничения числа сайтов рестрикции и получения более крупных фрагментов ДНК с помощью рестриктаз.

Формы разрывов двухцепочечных ДНК, образующихся под действием рестриктаз
а – 5’-выступающие (1), 3’-выступающие «липкие» (2) и «тупые» (3) концы ДНК, образующиеся под действием рестриктаз BamHI, KpnI и PvuII соответственно. Стрелками обозначены места разрывов цепей ДНК, пунктирной линией – ось симметрии сайтов рестрикции; б – лигирование с потерей сайта рестрикции
Названия рестриктаз складываются из первых букв видовых названий бактерий, в которых они обнаружены, например Eco – E. coli. В том случае, когда различные по специфичности действия рестриктазы присутствуют в клетках разных штаммов одного вида бактерий, в название рестриктазы вводят дополнительную букву, например рестриктазы Hinc и Hind выделены из бактериальных клеток Haemophilus influenzae, штаммы с и d. Цифры, следующие за буквенными обозначениями, отражают последовательность открытия соответствующих рестриктаз в клетках бактерий одного вида, например HaeI, HaeII и HaeIII из H. aegipticus.
Рестриктазы типа II – основной инструмент генной инженерии. Большинство рестриктаз типа II специфически узнают на ДНК тетра- и гексануклеотидные последовательности, а по крайней мере три из них – октануклеотиды. Чем короче олигонуклеотидная последовательность сайта рестрикции, узнаваемого рестриктазой, тем чаще он встречается в случайной последовательности нуклеотидов, в которой каждый из четырех нуклеотидов представлен с одинаковой частотой (50% А–Т-пар и 50% G–С-пар). Так, случайная тетрануклеотидная последовательность встречается в среднем через каждые 256 п.о. (4 4 ), а гексануклеотидная – через каждые 4096 п.о. (4 6 ). Однако в природных ДНК распределение нуклеотидов может заметно отличаться от случайного. Например, для эукариотических ДНК характерна низкая частота встречаемости динуклеотида CpG и соответственно сайтов рестрикции, содержащих эти динуклеотиды (рестриктазы HhaI, HpaII, TaqI, ThaI, AvaI, HaeII, HindII, SalI, SmaI, XhoI, XmaI). Существенное отклонение частоты встречаемости сайтов рестрикции от ожидаемого при случайном их распределении вдоль ДНК свойственно и хромосомам термофильных бактерий, которым, напротив, свойственно (хотя и не во всех случаях) обогащение по G–С-парам. Для большинства сайтов, узнаваемых рестриктазами типа II, характерно наличие в них симметрии второго порядка, т.е. узнаваемые ими последовательности представляют собой палиндромы, например у рестриктазы EcoRI – 5’-GAATTC-3’. Это означает, что нуклеотиды, расположенные в каждой из цепей на равном расстоянии от оси симметрии, комплементарны друг другу. Если точки расщепления противоположных цепей ДНК смещены друг относительно друга в сайте рестрикции, то образующиеся в результате рестрикции концы ДНК содержат выступающие одноцепочечные участки. Поскольку такие участки комплементарны сами себе и друг другу и могут между собой взаимодействовать, их часто называют «липкими» концами. В «липких» концах выступающим одноцепочечным участком может быть как 5’-, так и 3’-конец (рис. II.1,а). Формальным признаком образования 5’- или 3’-выступающих «липких» концов в сайтах рестрикции является расположение точки расщепления цепей ДНК в последовательности, используемой для обозначения сайта рестрикции, слева или справа от оси симметрии соответственно. У некоторых рестриктаз точки расщепления обеих цепей ДНК расположены непосредственно друг под другом в сайте рестрикции. В этом случае после расщепления ДНК «липких» концов не образуется, а получаются так называемые «тупые» концы, в которых нет выступающих одноцепочечных участков ДНК (см. рис. II.1,а). Имеется одно принципиальное функциональное различие между 5’- и 3’-выступающими «липкими» концами – последние невозможно пометить путем их достройки ДНК-полимеразой. Эту особенность следует иметь в виду при выборе рестриктаз для получения рестрикционных фрагментов ДНК, которые предполагается использовать в качестве зондов.
При конструировании рекомбинантных молекул полезно помнить, что, хотя рестриктазы BamHI, BclI, BglII и XhoII узнают разные сайты рестрикции, они образуют одни и те же «липкие» концы, GATC. То же характерно и для группы рестриктаз SalGI, XhoI и AvaI (NCGA). При лигировании (см. ниже) фрагментов ДНК, образованных рестриктазами одной из таких групп, происходит их объединение, но при этом исходные сайты рестрикции теряются, так как в результате образуется новая непрерывная последовательность нуклеотидов (см. рис. II.1,б). Сайты рестрикции для некоторых рестриктаз II типа не являются симметричными. Например, рестриктаза HgaI узнает асимметричную последовательность 5’-GACGC-3’, а одноцепочечные разрывы вносит в противоположные цепи ДНК, отступя вправо на 5 и 10 нуклеотидов соответственно:
Последовательности нуклеотидов образующихся «липких» концов являются уникальными для каждого такого сайта рестрикции. Вследствие этого рестрикционные фрагменты ДНК, образовавшиеся под действием данной рестриктазы, в смеси соединяются друг с другом лишь в строго определенной исходной последовательности, которая задается уникальными последовательностями нуклеотидов в «липких» концах рестрикционных фрагментов ДНК. Например, при расщеплении этой рестриктазой репликативной формы ДНК фага fX174 образуется 14 фрагментов, которые in vitro объединяются в правильную последовательность с образованием инфекционной fX174-ДНК.
Универсальные рестриктазы для одноцепочечных ДНК
На основе рестриктаз, узнающих асимметричные последовательности нуклеотидов, разработана система, позволяющая расщеплять молекулы одноцепочечной ДНК в любой заданной точке. С этой целью синтезируется олигонуклеотид, 5’-концевая часть которого содержит сайт, узнаваемый такой рестриктазой, а последовательность нуклеотидов 3’-концевой части комплементарна участку ДНК, в который необходимо внести эндонуклеазный разрыв. В результате гибридизации олигонуклеотида с одноцепочечной ДНК образуется структура, изображенная на рис. II.2. При этом фермент взаимодействует с сайтом узнавания (участок I), а разрывы вносятся по местам, обозначенным стрелками. Таким образом, положение места эндонуклеазного расщепления будет целиком зависеть от последовательности нуклеотидов участка II синтетического олигонуклеотида, комплементарного ДНК-субстрату.

Расщепление одноцепочечной ДНК универсальной рестриктазой
а – синтетический олигонуклеотид; б – одноцепочечная ДНК.
После образования шпильки и гибридизации с одноцепочечной ДНК-мишенью олигонуклеотид образует сайт связывания рестриктазы (участок I) и сайт расщепления ДНК-ДНК-гибрида (участок II). Стрелки указывают места эндонуклеазного расщепления ДНК и олигонуклеотида
Изошизомеры. В клетках разных видов бактерий могут содержаться рестриктазы, узнающие одни и те же сайты рестрикции. Такие рестриктазы называют изошизомерами. Изошизомеры некоторых рестриктаз с успехом используются для обнаружения метилированных участков ДНК в геноме. Так, рестриктазы HhaI и HpaII расщепляют неметилированные последовательности GCGC и CCGG соответственно и утрачивают способность к расщеплению, если хотя бы один из остатков цитозина в этих сайтах метилирован. В то же время фермент MspI (изошизомер HpaII) расщепляет последовательность CCGG независимо от того, метилированы или неметилированы остатки цитозина в таком сайте. N-Метилирование остатков аденозина в ДНК можно обнаружить с помощью изошизомеров Sau3A (расщепляет как метилированные, так и неметилированные последовательности GATC), DpnI (расщепляет только метилированные последовательности G Me ATC) и MboI (расщепляет только неметилированные последовательности).
Действие рестриктаз на необычные субстраты. Помимо двухцепочечных ДНК многие рестриктазы способны использовать ДНК-РНК-гибриды в качестве субстрата. Это относится, в частности, к рестриктазам EcoRI, HindII, SalI, MspI, HhaI, AluI, TaqI и HaeIII. Некоторые рестриктазы, например HaeIII, HhaI и SfaI, способны расщеплять одноцепочечную ДНК фага fX174, хотя и со значительно меньшей скоростью, чем соответствующую двухцепочечную RF-форму. Такая способность была продемонстрирована для некоторых других рестриктаз, а также ДНК-субстратов. Остается неясным, узнают ли эти рестриктазы истинные одноцепочечные сайты или же последовательности нуклеотидов, заключенные в элементы вторичной структуры.
С развитием метода полимеразной цепной реакции (ПЦР) (см. ниже) часто возникает необходимость расщепления рестриктазами амплифицированных олигонуклеотидов недалеко от их концов, т.е. в условиях, когда сайт рестрикции фланкирован с одного из своих концов одним или несколькими нуклеотидами. В этом случае установлена четкая зависимость способности определенных рестриктаз расщеплять сайты рестрикции от количества фланкирующих сайт нуклеотидов. Данное свойство рестриктаз объясняют, в частности тем, что на самих концах двухцепочечной молекулы ДНК происходит локальное плавление двойной спирали ДНК с образованием коротких одноцепочечных участков, захватывающих сайт узнавания рестриктазами. Частично избежать локальное плавление можно понижением температуры реакционной смеси во время проведения рестрикции таких олигонуклеотидов.
Определение единицы активности рестриктазы. Расчет реакционной смеси для проведения рестрикции. Оптимальные условия работы рестриктаз. Звездная активность рестриктаз. Метил-чувствительные и метил-нечувствительные рестриктазы. Применение рестриктаз в ДНК-фингерпринт анализе (RLFP).
Молекулярные биологи рутинно используют рестриктазы для различных целей, включая картирование генома, анализ полиморфизма длин рестрикционных фрагментов (RLFP-анализ), секвенирование ДНК, клонирование генов. Хотя рестрикция и не является незаменимым методом для проведения различных экспериментов, ее проведение и использование обусловлено удобством, простотой и относительной дешевизной.
В настоящее время ученые выделяют от 3 до 8 типов (классов, подклассов) эндонуклеаз рестрикции, отличающихся по своему строению, функциям, условиям работы, а также сайтам действия. Однако в генетической инженерии широко используются рестриктазы II типа, которых насчитывается более 700 видов. Также к их достоинствам относятся: коммерческая доступность, удобство в применении, а, главное, способность гидролизовать молекулу ДНК в специфическом месте, которое называется палиндромом. Палиндромом называют короткие олигонуклеотидные последовательности (обычно от 4 до 8), которые одинаково читаются по обеим цепям ДНК в одном направлении. Согласно последним данным, «уникальными» ферментами (рестриктазами с уникальными палиндромными сайтами рестрикции) являются порядка 250 видов. Однако, на практике можно также использовать изошизомеры. Обычно, фермент имеет несколько изошизомеров, и, таким образом, дефицита многообразия рестриктаз, на сегодняшний день, практически не существует. Стоит отметить, что свойства рестриктаз, наличие изошизомеров и коммерческую доступность можно узнать на сайте фирмы-производителя (New England Biolabs, ThermoScientific – Fermentas, Promega, SibEnzyme и др.) ферментов или в базе данных REBASE (http://rebase.neb.com).
Все рестриктазы II типа для эффективного расщепления требуют присутствия ионов магния (обычно 10 мМ), в качестве кофактора. Поэтому особое внимание уделите содержанию ЭДТА в препарате ДНК. Большинство реакций следует проводить при температуре +37 0 С, однако, некоторые ферменты требуют меньших (SmaI, +25 0 С) или больших (TaqI, +65 0 С) значений. Для большинства ферментов значение оптимума рН лежит в интервале 7.0 – 8.0, а оптимум концентрации NaCl 50 – 100 мМ. Однако, некоторые ферменты очень чувствительны к наличию специфических добавок. Например, SmaI для своей работы требует наличия в буферном растворе KCl вместо NaCl. Восстановительные агенты, такие так дитиотреитол или бета-меркаптоэтанол обычно не влияют на активность фермента, однако, предохраняют его от окисления (растворенного кислорода). Бычий сывороточный альбумин (БСА) часто добавляется в реакционную смесь в качестве стабилизирующего агента. Он повышает концентрацию белков в растворе, приближая данное значение к внутриклеточной концентрации, покрывает гидрофобную поверхность пластиковых пробирок, предохраняет от денатурации молекулу фермента. БСА положительно влияет на активность многих рестриктаз при концентрации 100 мкг/мл. В некоторых случаях молекулы ферментов (например, EcoRI, NotI) стабилизируют путем введения в реакционную смесь неионных детергентов, таких как Тритон Х-100 или Твин-20. Еще более редкий случай – это использование S-аденозилметионина для активации эндонуклеазы BsgI.
Какими критериями руководствуются для выбора той или иной эндонуклеазы рестрикции?
Обычно, для различных экспериментов необходимо и удобно добавлять 1 мкл фермента на реакцию. Вследствие этого, большинство фирм-производителей стандартизуют концентрацию фермента в фасовке (обычно от 2 до 20 ед./мкл). Однако многие производители очень часто производят ферменты в высокой концентрации (до 100 ед./мкл), которые подходят для гидролиза больших количеств ДНК. Таким образом, когда вы используете ферментные препараты с низкой концентрацией, возрастает объем фермента. В данном случае не забывайте, что концентрация глицерина в рестрикционной смеси не должна превышать 5 %, для того чтобы избежать звездной активности. Если же вы используете ферментный препарат с высокой концентрацией, то его необходимо предварительно развести до оптимальной концентрации. Аликвоту можно хранить при ‑20 0 С длительное время.
Что такое звездная активность?
Когда условия расщепления ДНК каким-либо ферментом не оптимальны, гидролиз макромолекулы может происходить не по канонической палиндромной последовательности. Такое изменение активности называется звездной активностью. Обычно в таких случаях, последовательность ДНК, по которой происходит гидролиз, отличается от палиндромной на 1 – 3 нуклеотида.
Впервые звездная активность была обнаружена при гидролизе молекулы ДНК ферментом EcoRI в буферном растворе с низкой ионной силой и высоким рН (Polisky et al, 1975). В таких условиях наряду с палиндромом данного фермента G/AATTC, фермент был способен узнавать и расщеплять ДНК по следующему сайту N/AATTC. Позднее было отмечено, что звездная активность фермента также проявляется при замене ионов Mg 2+ в буферном растворе на ионы Mn 2+ (Hsu и Berg, 1978). Длительное инкубирование раствора ДНК с ферментом, высокая концентрация ДНК и фермента, глицерина и органических растворителей в рестрикционной смеси также являются причинами появления звездной активности. Рекомендуется поддерживать концентрацию глицерина в растворе не более 5%. Для этого обычно следует помнить, что объем добавляемого фермента не должен превышать 10% от объема реакционной смеси. Не стоит забывать, что неспецифическое расщепление молекулы ДНК очень часто наблюдается, вследствие недостаточной чистоты фермента и присутствием других рестриктаз. Для установления данной причины, необходимо провести гидролиз ДНК в оптимальных рабочих условиях фермента. Также можно варьировать различные концентрации ферментного препарата, ДНК, время инкубации, температуру, то есть, подобрать неоптимальные условия работы примесного энзима.
Каким образом метилирование влияет на активность рестриктаз?
Многие эндонуклеазы рестрикции чувствительны к метилированию. Это является частью природного механизма защиты бактериальной клетки от экзогенной ДНК. Однако множество штаммов E. coli, предназначенных для клонирования, экспрессируют гены EcoKI, dam или dcm метилаз. Например, dam метилаза узнает специфическую последовательность GATC и метилирует аденин в N6-положении. Эндонуклеаза MboI также узнает GATC, поэтому ДНК будет гидролизоваться лишь в случае, когда препарат получен из штамма E. coli, дефицитного по метилазам, например, GM2163. DpnI – одна из немногих эндонуклеаз, способных к расщеплению метилированной ДНК. Она совместима с dam + штаммами. Dcm метилаза узнает специфическую последовательность CC(A/T)GG и метилирует второй цитозин в С5-положении. Она блокирует работу таких ферментов, как StuI и AatI (Song, Rueter, Geiner, 1988).
Интересно, что метилирование ДНК может повлиять на активность фермента, даже и без совпадения сайтов. Так, довольно популярный фермент XbaI имеет сайт узнавания следующего вида: 5’ TCTAGA 3’, что не является специфическим сайтом для метилаз. Однако если впереди или после данного палиндома будут находиться последовательности GA или TC соответственно, то рестриктный сайт будет подвержен dam-метилированию (GATCTAGATC).
Проблемы с метилированием также возникают при работе с препаратами ДНК, выделенными их клеток млекопитающих и растений. Молекулы ДНК млекопитающих могут метилироваться по цитозину С5-положении у последовательностей CG, а в ДНК растений обычно метилируются CG или CNG последовательности. Большинство штаммов E. coli имеют так называемую mcr систему рестрикции, которая обеспечивает расщепление метилированной ДНК. Поэтому, если целью вашей работы является клонирование метилированной ДНК, то необходимо использовать mcr-дефицитные штаммы.
Каким образом субстрат влияет на активность рестриктаз?
Эндонуклеазы рестрикции часто используются сразу же после проведения реакции ПЦР. При этом не стоит забывать, что на активность фермента значительно может влиять буферный раствор, применяемый для амплификации, а также праймеры. К примеру, было показано, что присутствие праймеров в концентрации 1 μМ ингибирует работу рестриктаз SmaI и NdeI (Ambrol, 1993). Однако в большинстве случаев присутствие праймеров, даже в концентрации 100 μМ, не влияет на эффективность расщепления. Если вы не удовлетворены результатами гидролиза молекулы ДНК, полученного после реакции ПЦР, в таком случае необходимо провести обессоливающую хроматографию, диафильтрацию на мембранном фильтре. Это более быстрая процедура, чем экстракция ДНК смесью «фенол-хлороформ» и последующее осаждение спиртом.
В зависимости от положения сайтов рестрикции в молекуле ДНК (концевые или внутренние) эффективность расщепления может различаться. Многие рестриктазы могут гидролизовать ДНК по палиндромным последовательностям, расположенным на концах молекулы. Однако перед сайтом должно находиться хотя бы 1 – 4 нуклеотида, для возможности эффективного узнавания палиндрома. Причем, в зависимости от того, какие нуклеотиды расположены перед сайтом, эффективность гидролиза может тоже быть различной. Некоторые примеры приведены в следующей таблице.
Фермент
Олигонуклеотидная последовательность
Длина
цепи



