Что такое следовые потенциалы
Следовые потенциалы. Последний участок фазы реполяризации для некоторых видов потенциалов действия бывает замедленным; хорошим примером служит потенциал действия мышцы на рис. 2.4. Приблизительно через 1 мс после начала потенциала действия наблюдается отчетливый перегиб кривой реполяризации; следующее за ним медленное изменение потенциала называется деполяриза-ционным следовым потенциалом. В других тканях, например в нейронах спинного мозга, кривая деполяризации быстро пересекает уровень потенциала покоя, так что на некоторое время потенциал становится более отрицательным, чем потенциал покоя. Это явление получило название гиперполяризационный следовой потенциал (рис. 2.5).
Природа потенциала действия
Порог и возбудимость. Каким образом потенциал покоя, обычно поддерживаемый на постоянном уровне посредством только что обсуждавшихся механизмов, нарушается до такой степени, что возникает потенциал действия? Потенциалы действия генерируются при деполяризации мембраны от потенциала покоя до примерно —50 мВ. Механизмы развития этой начальной деполяризации будут рассмотрены позднее. Уровень потенциала, при котором деполяризация приводит к потенциалу действия, называется порогом (рис. 2.5). При таком пороговом потенциале заряд мембраны становится нестабильным; он нарушается посредством внутреннего механизма, который ведет к реверсии полярности-быстрому нарастанию потенциала действия до пика. Это состояние автоматического прогрессирующего нарушения мембранного заряда называется возбуждением. Обычно возбуждение продолжается менее 1 мс. Оно подобно взрыву-протекает мощно, но быстро завершается. После фазы деполяризации наступает процесс восстановления заряда мембраны, присущего состоянию покоя.
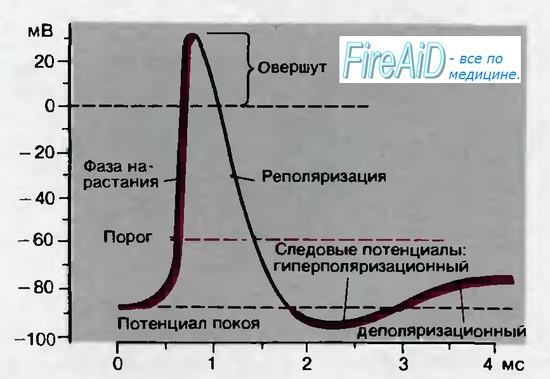 Рис. 2.5. Временной ход потенциала действия в нейроне; показаны последовательные фазы потенциала действия, описанные в тексте
Рис. 2.5. Временной ход потенциала действия в нейроне; показаны последовательные фазы потенциала действия, описанные в тексте
Клетки, в которых можно вызвать потенциалы действия, называются возбудимыми. Возбудимость является типичным свойством нервных и мышечных клеток. Клетки каждого типа характеризуются собственным постоянным временным ходом потенциала действия. Он практически не зависит от источника или частоты возбуждения клетки. Поскольку форма потенциала действия постоянна, говорят, что возбуждение протекает по закону «все или ничего».
Следовые потенциалы




![]()
![]()
Восстановление мембранного потенциала покоя после осуществления пика потенциала действия происходит не сразу, а после следовых процессов, выражающихся в характерных изменениях мембранного потенциала. Эти изменения, следующие за пиком потенциала действия, называют следовыми потенциалами. Различают два вида следовых потенциалов – следовой деполяризационный и следовой гиперполяризационный потенциал (рис. 62). Величина следовых потенциалов обычно не превышает нескольких милливольт, а длительность их у различных нервных волокон составляет от нескольких миллисекунд до нескольких сотен миллисекунд.
Следовой деполяризационный потенциал наблюдается в тот период, когда реполяризация, обусловленная выходом ионов К + из клетки, начинает происходить медленнее. Это вызвано тем, что по мере выхода катионов калия из клетки, ослабевает осмотическая сила выталкивания и усиливается электрическая сила отталкивания катионов от положительно заряженной наружной поверхности мембраны. Во время следового деполяризационного потенциала мембрана клетки имеет меньшую величину заряда, чем в покое, т.е. она слегка деполяризована. В тот период, когда величина деполяризационного потенциала находится между величиной потенциала покоя и величиной критического потенциала, клетка обладает повышенной возбудимостью. В это время она может ответить потенциалом действия и ответной реакцией даже на более слабые раздражители, на которые в обычных условиях она не отвечает.
В безмякотных нервных волокнах вслед за пиком потенциала действия развивается следовой гиперполяризационный потенциал. Он обусловлен более длительным сохранением повышенной проницаемости мембраны для ионов калия. Вследствие этого катионов калия выходит из клетки больше и наружная поверхность мембраны приобретает на какое-то время более положительный заряд, а внутренняя поверхность мембраны – более отрицательный заряд, чем в покое. Наблюдается период гиперполяризации мембраны, во время которого клетка менее возбудима, чем в покое.
В мякотных нервных волокнах следовые потенциалы имеют более сложный характер. Следовой деполяризационный потенциал может переходить в следовой гиперполяризационный потенциал и лишь после этого происходит полное восстановление потенциала покоя. Следовые потенциалы в значительно большей мере, чем пики потенциалов действия, чувствительны к изменениям ионного состава среды, кислородному снабжению клетки и т.д. Характерной особенностью следовых потенциалов является их способность изменяться в процессе ритмического раздражения.
На осуществление одного потенциала действия вместе со следовыми потенциалами расходуется очень мало ионов. Разницы концентрации ионов калия и натрия внутри и снаружи нервного волокна может хватить на 500 000 импульсов. Но для длительной активности нейронов необходимо восстанавливать градиенты ионных концентраций, что обеспечивается работой натрий-калиевого насоса мембраны клетки.
5.7. Проведение нервных импульсов
5.7.1. Распространение нервного импульса по безмякотным нервным волокнам
Распространение потенциалов действия по ткани, особенно нервного импульса по нервам, является самым быстрым и точно адресованным способом передачи информации в организме. Скорость передачи нервного импульса в быстропроводящих волокнах двигательных нервов (тип волокна Аα) достигает 120 м/с. Другие способы передачи информации гораздо медлительнее: гуморальный не превышает 0,5 м/с (скорость тока крови в аорте), аксонный транспорт веществ от тела нейрона к окончаниям аксона не превышает 40 см в сутки.
Распространение потенциалов действия, или проведение возбуждения, осуществляется по мембране нервных и мышечных клеток. Обязательным условием проведения нервного импульса является наличие на всем протяжении или в ограниченных, но повторяющихся участках волокна электровозбудимых (потенциал-зависимых) ионных каналов, ответственных за формирование потенциала действия. Проведение нервных импульсов по безмякотным и мякотным нервным волокнам имеет свои особенности.
При нанесении раздражения на безмякотное нервное волокно, не содержащее в своей оболочке электроизолирующего вещества миелина, в раздражаемом участке изменяется ионная проницаемость мембраны. Раскрываются активационные ворота натриевых каналов и ионы натрия поступают внутрь нервного волокна. Развивается потенциал действия и мембрана возбужденного участка заряжается снаружи отрицательно, а внутри положительно (рис. 63).

Входящий внутрь волокна положительный ток, создаваемый катионами натрия, должен выходить через мембрану наружу, чтобы замкнуть электрическую цепь. Соседние невозбужденные участки мембраны нервного волокна имеют на наружной поверхности положительный заряд, а на внутренней – отрицательный. Между возбужденным и невозбужденными участками мембраны возникают местные (круговые) электрические токи, которые, проходя через невозбужденные участки мембраны, раздражают их. В результате в них изменяется ионная проницаемость мембраны, эти участки возбуждаются и в них возникает потенциал действия. Процесс повторяется и таким образом возбуждение проводится в обе стороны по нервному волокну от первоначального места нанесения раздражения. Следовательно, потенциал действия распространяется по нервному волокну с помощью потенциал-зависимых натриевых каналов, которые открываются и закрываются по мере движения нервного импульса.
 Местные токи выходят из волокна наружу в области наименьшего сопротивления мембраны. Расстояние, на которое распространяется каждый «виток» местных токов по волокну, тем больше, чем ниже сопротивление цитоплазмы и выше сопротивление мембраны. В тонких безмякотных нервных волокнах это расстояние, т.е. протяженность возбужденного участка, тем больше, чем толще волокно, но не превышает 1 мм.
Местные токи выходят из волокна наружу в области наименьшего сопротивления мембраны. Расстояние, на которое распространяется каждый «виток» местных токов по волокну, тем больше, чем ниже сопротивление цитоплазмы и выше сопротивление мембраны. В тонких безмякотных нервных волокнах это расстояние, т.е. протяженность возбужденного участка, тем больше, чем толще волокно, но не превышает 1 мм.
Чем дальше от точки раздражения, тем слабее местные токи. Поэтому на расстоянии больше 1 мм они не могут проходить через мембрану и вызывать образование новых потенциалов действия. Если источник раздражения убрать, то в первоначальном месте нанесения раздражения благодаря действию натрий-калиевого насоса восстановится мембранный потенциал покоя.
В безмякотных нервных волокнах местные токи проходят последовательно практически через каждый соседний участок мембраны волокна, т.к. длина их «витка» не превышает 1 мм. Поэтому скорость распространения нервного импульса невелика (0,5–3 м/с) и его передача требует большего расхода энергии, чем при распространении импульсов по мякотным нервным волокнам.
5.7.2. Распространение нервного импульса по мякотным нервным волокнам. Особенности проведения возбуждения по нервным волокнам. Типы нервных волокон
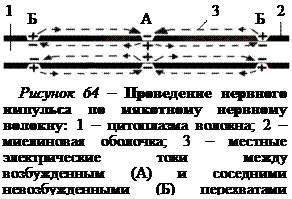 Величина потенциала действия, возникшего в одном перехвате Ранвье, в 5–10 раз больше той пороговой величины, которая необходима для возбуждения соседнего перехвата. Поэтому потенциал действия, возникший в одном перехвате, способен вызвать возбуждение не только в находящемся рядом, но и в соседних 2–3 перехватах. Это создает гарантию проведения импульсов по нервному волокну, даже если 1–2 ближайших перехвата Ранвье повреждены.
Величина потенциала действия, возникшего в одном перехвате Ранвье, в 5–10 раз больше той пороговой величины, которая необходима для возбуждения соседнего перехвата. Поэтому потенциал действия, возникший в одном перехвате, способен вызвать возбуждение не только в находящемся рядом, но и в соседних 2–3 перехватах. Это создает гарантию проведения импульсов по нервному волокну, даже если 1–2 ближайших перехвата Ранвье повреждены.
Проведение возбуждения по нервным волокнам характеризуется следующими закономерностями.
1. Двустороннее проведение возбуждения по нервному волокну. Если на любом участке нерва нанести раздражение, то возникший нервный импульс будет распространяться по нерву в обе стороны от участка, подвергшегося раздражению.
2. В каждом нервном волокне возбуждение проводится изолированно. Это обусловлено тем, что местные круговые токи, выходящие в межклеточную жидкость из возбужденного волокна, почти не проникают в соседние невозбужденные нервные волокна из-за большого электрического сопротивления их оболочек. Изолированное проведение импульсов по нервным волокнам обеспечивает точную адресную передачу информации.
3. Малая утомляемость нервного волокна. Расход энергии в нервном волокне на единицу массы при проведении нервных импульсов примерно в миллион раз меньше, чем в работающей мышце. При проведении одного импульса по нервному волокну используются лишь около одной миллионной части запасов ионов натрия и калия. Поэтому для восстановления градиентов ионных концентраций необходимо очень малое количество АТФ. При нормальном снабжении кислородом и питательными веществами нерв практически неутомляем и может проводить возбуждение в течение многих часов.
4. Необходимость анатомической и физиологической целостности нервного волокна для проведения возбуждения. Перерезка нервного волокна, травма его поверхностной мембраны нарушает проводимость. Проведение импульсов также прекращается при сохранении анатомической целостности нерва, но нарушении его физиологических свойств. Температурные и электрические воздействия на нервное волокно, блокада натриевых каналов мембраны, действие анестезирующих средств, механическое сдавливание нерва, прекращение кровоснабжения нарушают проведение возбуждения. После прекращения действия факторов, нарушающих физиологическую целостность нерва, проведение нервных импульсов по нервным волокнам, как правило, восстанавливается.
Типы нервных волокон.Скорость распространения импульсов зависит от диаметра нервных волокон, наличия и толщины миелиновой оболочки. Нервы у человека состоят из трех основных типов волокон: А, В и С. Тип А включает наиболее толстые хорошо миелинизированные афферентные и эфферентные нервные волокна и делится на 4 группы в зависимости от диаметра волокна и скорости проведения возбуждения. Волокна группы Аα имеют диаметр 13–22 мкм и скорость проведения импульсов 70–120 м/с; группы Аβ – диаметр 8–13 мкм и скорость 40–70 м/с; группы Аγ – диаметр 4–8 мкм, скорость 15–40 м/c; группы Аδ – диаметр 1–4 мкм и скорость проведения возбуждения 5–15 м/с.
Тип В включает в себя слабомиелинизированные волокна автономной нервной системы, имеющие диаметр 1–3 мкм и скорость проведения возбуждения 3–14 м/с. В тип С объединены безмякотные волокна автономной нервной системы диаметром 0,5–1 мкм и со скоростью проведения импульсов 0,5–2 м/с.
Следовые потенциалы
Различают отрицательные и положительные следовые потенциалы. Амплитуда как тех, так и других не превышает нескольких милливольт, а длительность варьирует от нескольких миллисекунд до нескольких десятков или даже сотен миллисекунд. Следовые потенциалы связаны с восстановительными процессами, медленно развивающимися в нервных и мышечных волокнах после окончания возбуждения.
Следовой положительный потенциал выражается в усилении нормальной поляризации — гиперполяризации — мембраны. Он особенно хорошо выражен в безмякотных нервных волокнах. Так, в безмякотном гигантском аксоне кальмара нисходящая фаза потенциала действия непосредственно переходит в положительный следовой потенциал, амплитуда которого достигает примерно 15 мв. и лишь затем мембранный потенциал возвращается к исходному уровню покоя ( рис. 119 ).
В миелинизированных нервных волокнах следовые изменения потенциала имеют более сложный характер: следовой отрицательный потенциал часто сменяется следовым положительным потенциалом, затем иногда развивается новая электроотрицательность и лишь после этого происходит полное восстановление потенциала покоя.
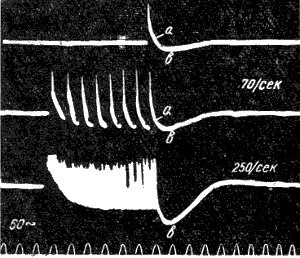
При ритмическом раздражении нерва следовые потенциалы суммируются, вследствие чего их амплитуда и длительность возрастают ( рис. 120 ).
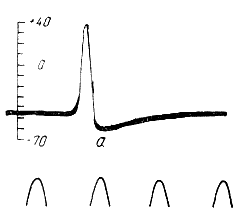 |  |
| Рис. 119. Потенциал действия гигантского аксона кальмара, записанный с помощью внутриклеточного микроэлектрода (по Ходжкину и Хаксли). Выражен следовой положительный потенциал (а). В нижней части рисунка отметка времени — 500 колебании в 1 секунду. |



