Эмбриональное развитие
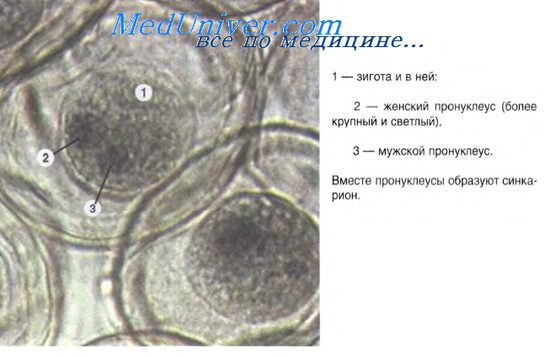
От момента образования зиготы и до выхода зародыша из яйцевых оболочек длится эмбриональный период развития.
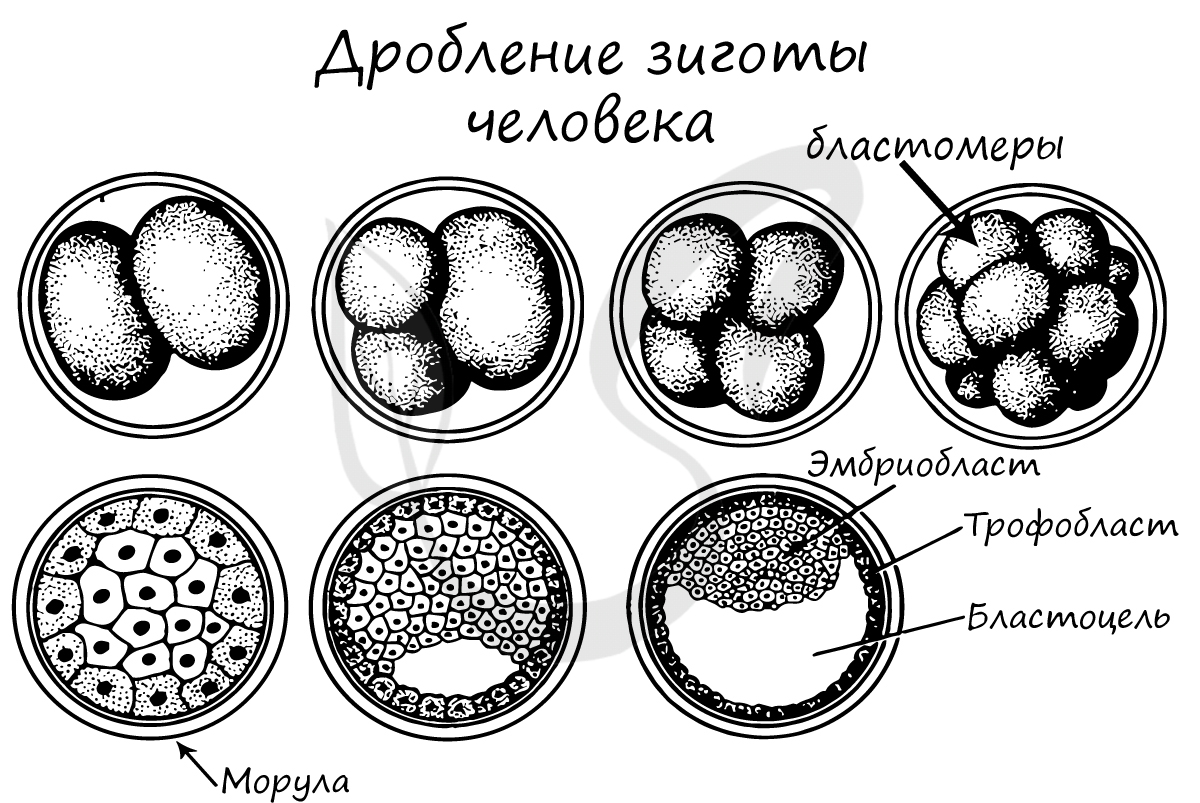
Дробление зиготы
Важная особенность дробления в том, что не происходит увеличение в размере зародыша: клетки дробятся (делятся) настолько быстро, что не успевают накопить цитоплазматическую массу. Дробление зиготы человека является полным неравномерным асинхронным.
Бластуляция

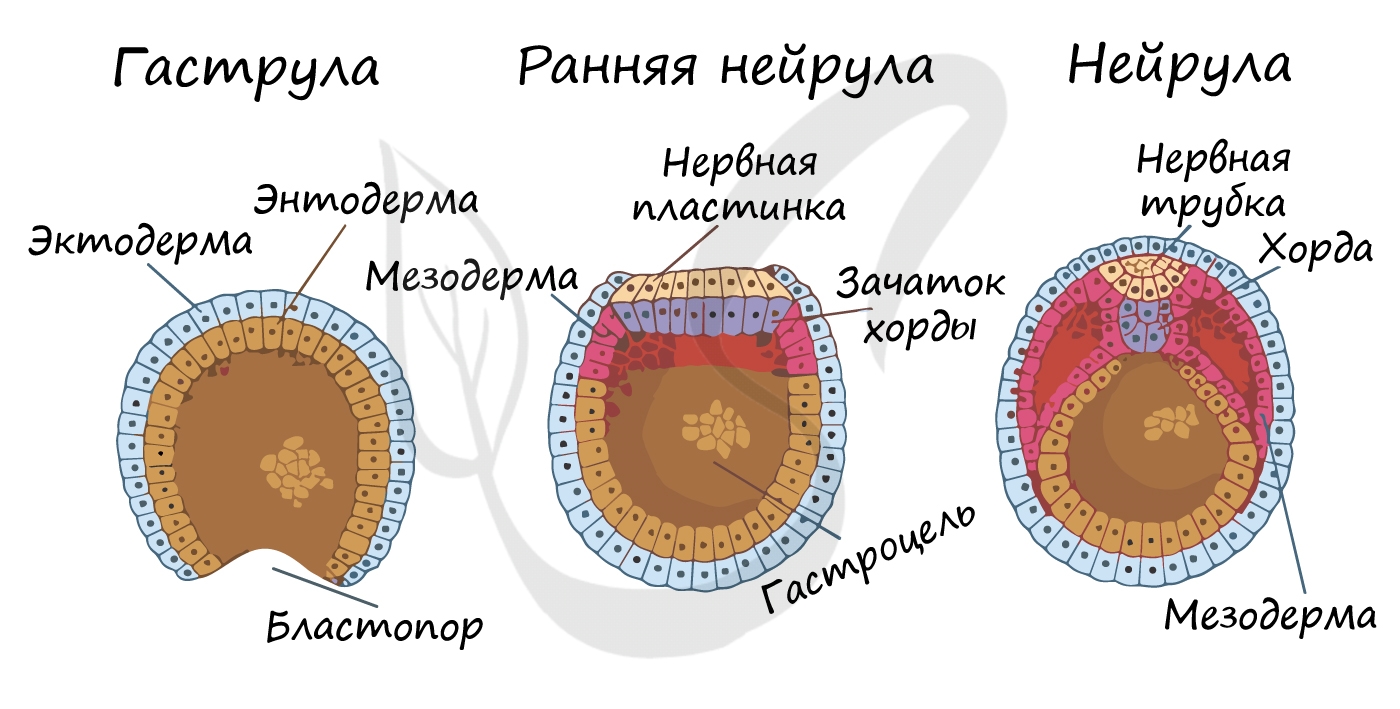
Гаструляция (греч. gaster — желудок, чрево)
Гаструляцией называют стадию эмбрионального развития, в ходе которой клетки, возникшие в результате дробления зиготы, формируют три зародышевых листка: эктодерму, мезодерму и энтодерму.

У первичноротых животных на месте первичного рта (бластопора) образуется ротовое отверстие. К первичноротым относятся: кишечнополостные, плоские, круглые и кольчатые черви, моллюски, членистоногие.
У вторичноротых на месте бластопора формируется анальное отверстие, а ротовое отверстие образуется на противоположном полюсе. К вторичноротым относят хордовых и иглокожих (морских звезд, морских ежей).


Нейрула
Эта стадия следует за гаструлой. Ранняя нейрула представляет собой трехслойный зародыш, состоящий из энто-, экто- и мезодермы. На этапе нейрулы происходит закладка отдельных органов.

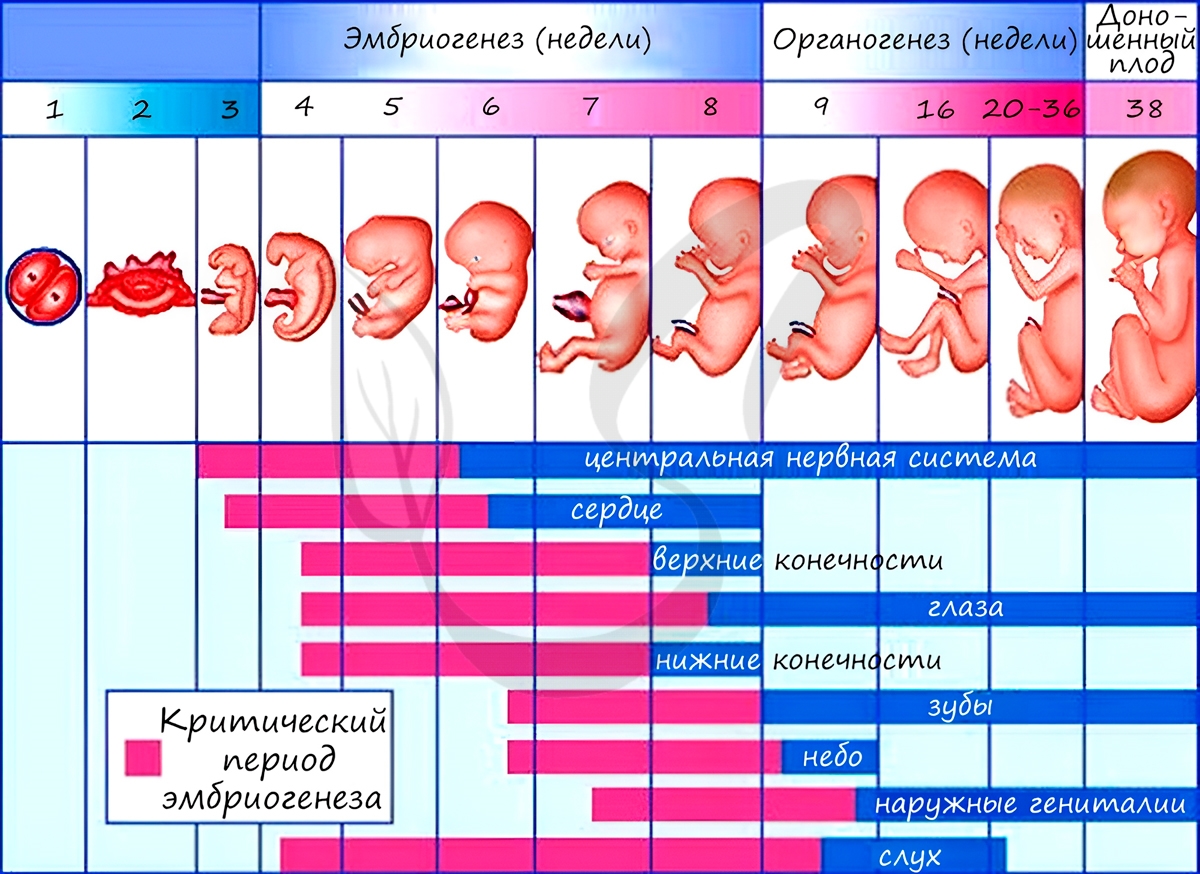
Все три зародышевых листка требуют нашего особого внимания, а также понимания того, какие органы и структуры из них образуются.

Из зародышевых листков образуются ткани, органы и системы органов. Такой процесс называется органогенезом. В период закладки органов важное значение имеет воздержание матери от вредных привычек (алкоголь, курение), которые могут нарушить процесс дифференцировки клеток и привести к тяжелейшим аномалиям, уродствам плода.
Некоторые лекарства также могут оказывать на плод тератогенный эффект (греч. τέρας — чудовище, урод), приводя к развитию уродств. Периоды закладки органов и система органов вследствие их большой важности носят название критических периодов эмбриогенеза.
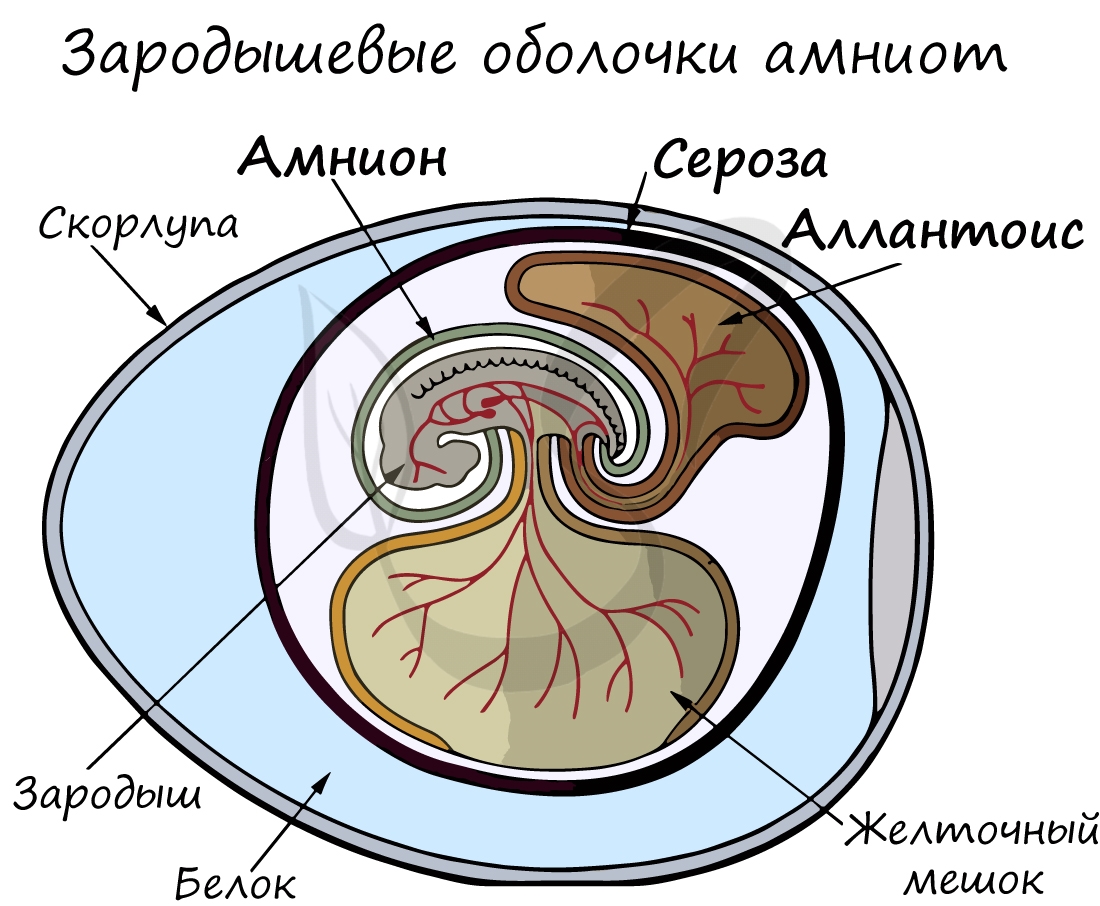
Анамнии и амниоты
К анамниям относятся рыбы, земноводные.

Зародышевый орган, аллантоис, является органом дыхания и выделения.
За счет особых оболочек, развивающихся в ходе эмбрионального развития, амниона и серозы, у амниот формируется амниотическая полость. В ней находится зародыш, окруженный околоплодными водами. Благодаря такому гениальному устройству, амниотам для размножения и развития более не нужно постоянное нахождение в водоеме, они «обрели независимость» от него.
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Что такое стадия дробления
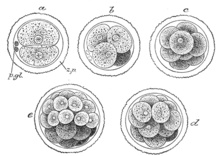
Дробление представляет собой серию митотических делений зиготы с образованием многих дочерних клеток (бластомеров) меньшего размера. Митотические деления зиготы, а в последующем — бластомеров происходят с увеличением числа клеток, но без увеличения их массы, поэтому именуются дроблением.
У человека дробление не имеет принципиальных отличий от такового у других представителей позвоночных, однако протекает гораздо медленнее. Дробление полное, или голобла-стическое (борозды дробления проходят через весь зародыш), неравномерное (в результате дробления образуются дочерние клетки — бластомеры неравной величины) и асинхронное (разные бластомеры дробятся с различной скоростью, поэтому зародыш на отдельных стадиях дробления содержит нечетное число клеток).
Первое деление дробления продолжается в среднем около 30 часов, последующие — более кратковременны (около 20-24 часов). В процессе дробления зародыш перемещается по маточной трубе и на 6-е сутки развития попадает в полость матки.
Бластомеры первой генерации у человека, как и зигота, тотипотентны (каждый бластомер способен развиться в полноценный организм). До стадии 8 бластомеров клетки зародыша формируют рыхлую неоформленную группу, и только после третьего деления устанавливают между собой плотные контакты, образуя компактный клеточный шар из 16 бластомеров, именуемый морулой. Компактизация создает условия для развития наружной клеточной массы и внутренней клеточной массы.
Последняя — это материал будущего тела зародыша (эмбриобласта) и внезародышевых органов. Бластомеры наружной клеточной массы — мелкие и многочисленные (их примерно в 10 раз больше, чем клеток внутренней клеточной массы), являются источником развития трофобласта.
Когда морула попадает в проксимальный отдел маточной трубы и далее — в полость матки, через ее прозрачную зону начинает проникать содержащаяся в маточной трубе и матке жидкость. Происходит кавитация морулы. Сначала жидкость накапливается между клетками и образует небольшие промежутки, которые затем сливаются в единую полость внутри морулы (бластоцель). В образовании жидкости и кавитации участвуют также клетки трофобласта, секретирующие жидкость.
С момента появления полости зародыш именуется бластоцистой. Клетки внутренней клеточной массы бластоцисты локализованы на одном из полюсов и обращены в полость. Клетки наружной клеточной массы уплощаются и, ограничивая полость, формируют оболочку бластоцисты — трофобласт. В период перемещения дробящегося зародыша по маточной трубе большое значение имеет тот факт, что сохраняющаяся прозрачная зона предотвращает прилипание бластоцисты к стенкам трубы и зародыш попадает в полость матки. Здесь он освобождается от прозрачной зоны и начинает имплантироваться (погружаться) в слизистую оболочку матки. Имплантация зародыша протекает параллельно с гаструляцией.
Дробление (эмбриология)

Дробле́ние — ряд последовательных митотических делений оплодотворенного или инициированного к развитию яйца. Дробление представляет собой первый период эмбрионального развития, который присутствует в онтогенезе всех многоклеточных животных. При этом масса зародыша и его объём не меняются, оставаясь такими же, как и в начале дробления. Яйцо разделяется на все более мелкие клетки — бластомеры. Характерная особенность дробления — ведущая регуляторная роль цитоплазмы в развитии. Характер дробления зависит от количества желтка и его расположения в яйце.
Содержание
Биологическое значение дробления
Характерные черты дробления
Дробление как особый этап онтогенеза животных имеет характерные черты, которые свойственны большинству животных, но могут отсутствовать у некоторых групп.
По всем этим характеристикам дробление млекопитающих резко отклоняется от типичного. Бластомеры делятся у них медленно, синхронность нарушается уже после 1—2 делений, в это же время активируется собственный геном зародыша.
Классификация типов дробления
На основе ряда существенных характеристик (степень детерминированности, полнота, равномерность и симметрия деления) выделяют ряд типов дробления. Типы дробления во многом определяются распределением веществ (в том числе, желтка) по цитоплазме яйца и характером межклеточных контактов, которые устанавливаются между бластомерами.
Дробление может быть: детерминированным и регулятивным; полным (голобластическим) или неполным (меробластическим); равномерным (бластомеры более-менее одинаковы по величине) и неравномерным (бластомеры не одинаковы по величине, выделяются две — три размерные группы, обычно называемые макро- и микромерами); наконец, по характеру симметрии различают радиальное, спиральное, различные варианты билатеризованных и анархическое дробление. В каждом из этих типов выделяют ряд вариантов.
По степени детерминированности
Детерминированное
Недетерминированное (регулятивное)
По степени полноты делений
Голобластическое дробление
Плоскости дробления разделяют яйцо полностью. Выделяют полное равномерное дробление, при котором бластомеры не различаются по размерам (такой тип дробления характерен для гомолецитальных и алецитальных яиц), и полное неравномерное дробление, при котором бластомеры могут существенно различаться по размерам. Такой тип дробления характерен для умеренно телолецитальных яиц.
Меробластическое дробление
Такой тип дробления типичен для телолецитальных яиц, богатых желтком (птицы, рептилии). Такое дробление называют также дискоидальным, так как в результате дробления на анимальном полюсе образуется небольшой диск клеток (бластодиск).
Такой тип дробления наблюдается у членистоногих.
По типу симметрии дробящегося яйца
Радиальное
Ось яйца является осью радиальной симметрии. Типично для ланцетника, осетровых, амфибий, иглокожих, круглоротых.
Спиральное
В анафазе бластомеры разворачиваются. Отличается лево-правой дисимметрией (энантиоморфизм) уже на стадии четырёх (иногда двух) бластомеров. Типично для некоторых моллюсков, кольчатых и ресничных червей.
Билатеральное
Имеется 1 плоскость симметрии. Типично для аскариды.
Анархическое
Бластомеры слабо связаны между собой, сначала образуют цепочки или бесформенную массу; часто у одного вида встречаются разные варианты расположения бластомеров. Типично для кишечнополостных.
Литература
Полезное
Смотреть что такое «Дробление (эмбриология)» в других словарях:
Дробление — Дробление: Дробление (технология) измельчение твёрдого тела до определенного размера; Дробление (полиграфия) воспроизведение на оттиске одного и того же печатающего элемента дважды, со смещением; Дробление (эмбриология) ряд… … Википедия
Дробление (значения) — Дробление: Дробление (технология) измельчение твёрдого тела до определенного размера; Дробление (полиграфия) воспроизведение на оттиске одного и того же печатающего элемента дважды, со смещением; Дробление (эмбриология) ряд последовательных… … Википедия
Эмбриология — (от древнегреческого ἔμβρυον, зародыш, «эмбрион»; и λογία, логия) это наука, изучающая развитие зародыша. Зародышем называют любой организм на ранних стадиях развития до рождения или вылупления, или, в случае растений, до момента прорастания.… … Википедия
Дробление яйца — Дробление: эмбрион млекопитающего. z.p. zona striata, p.gl полярные тельца, a. двуклеточная стадия, b. четырёхклеточная стадия, c. восьмиклеточная стадия, d, e. морула Дробление ряд последовательных митотических делений оплодотворенного… … Википедия
дробление радиальное — ЭМБРИОЛОГИЯ ЖИВОТНЫХ ДРОБЛЕНИЕ РАДИАЛЬНОЕ – первое дробление меридиональное, проходит в меридиональной плоскости яйца. Второе дробление также меридиональное, оно проходит через главную ось яйца, но под прямым углом к плоскости первого дробления.… … Общая эмбриология: Терминологический словарь
дробление анархическое — ЭМБРИОЛОГИЯ ЖИВОТНЫХ ДРОБЛЕНИЕ АНАРХИЧЕСКОЕ [БЕСПОРЯДОЧНОЕ, ХАОТИЧНОЕ] – дробление яиц метагенетических медуз – Oceania armata. Первая борозда дробления меридиональная, врезающаяся, появляется она на анимальном полюсе. Вторая борозда тоже… … Общая эмбриология: Терминологический словарь
дробление гетероквадратное — ЭМБРИОЛОГИЯ ЖИВОТНЫХ ДРОБЛЕНИЕ ГЕТЕРОКВАДРАТНОЕ – дробление аннелид, моллюсков, немертин, планарий. Неравномерное спиральное дробление, когда клетки основного квартета (первые четыре бластомера) неодинаковы по величине, то и их производные также… … Общая эмбриология: Терминологический словарь
дробление асинхронное — ЭМБРИОЛОГИЯ ЖИВОТНЫХ ДРОБЛЕНИЕ АСИНХРОННОЕ – дробление яиц телолецитальных (амфибия). Деление вегетативных бластомеров происходит медленней, по сравнению с бластомерами анимального полюса … Общая эмбриология: Терминологический словарь
дробление билатеральное — ЭМБРИОЛОГИЯ ЖИВОТНЫХ ДРОБЛЕНИЕ БИЛАТЕРАЛЬНОЕ [ДВУСТОРОННЕ СИММЕТРИЧНОЕ] – дробление яиц нематод, коловраток, асцидий. Характеризуется появлением билатеральной симметрии в расположении бластомеров уже на ранних этапах дробления. Каждый бластомер… … Общая эмбриология: Терминологический словарь
дробление голобластическое — ЭМБРИОЛОГИЯ ЖИВОТНЫХ ДРОБЛЕНИЕ ГОЛОБЛАСТИЧЕСКОЕ [ПОЛНОЕ] – дробление яиц алецитального (плоские черви), изолецитального типа (ланцетник) и некоторых яйцевых клеток телолецитального типа (амфибии). Дробятся все части зиготы. При дроблении вся… … Общая эмбриология: Терминологический словарь
Дробление
Дробление – это процесс разрушения кусков полезного ископаемого под действием внешних сил.
По технологическому назначению различают следующие виды дробления:
По различию использованных сил различают следующие типы дробления:
В обогащении полезных ископаемых применяется механическое дробление, остальные типы дробления относятся к другим сферам. Дробление полезного ископаемого осуществляется только в воздушной среде (сухое дробление), вода может подаваться в процесс лишь для пылеподавления.
По виду основной необратимой деформации различают следующие способы дробления:
Пределы крупности дробления и последующего измельчения определяются многими факторами, основными из которых являются дробимость, крупность вкрапленности полезного ископаемого и применяемые методы обогащения.
Основным показателем, характеризующим интенсивность процесса дробления, является степень дробления. Степень дробления показывает во сколько раз первоначальный размер куска полезного ископаемого уменьшился в результате дробления.
Степень дробления – это отношение размера максимальных кусков исходного материала к размеру максимальных кусков дроблёного продукта.
Степень дробления – всегда целая величина, при расчёте полученные результаты округляются в большую строну. Для определения степени дробления вычисляют максимальный среднеарифметический или среднегеометрический диаметр кусков исходного питания и дроблённого продукта. Максимальная величина степени дробления, которую можно получить в одной дробилке, не превышает 8-10, исключение составляют лишь конусные инерционные дробилки, где степень дробления может достигать 15-20. Поэтому дробление руды на обогатительных фабриках никогда не осуществляется в одну стадию, как правило, это 3-4 стадии дробления. При дроблении в несколько последовательных стадий общая степень дробления будет равна произведению стадиальных степеней дробления.
Стадия дробления – это часть общего процесса дробления, которую можно осуществить в одном аппарате. Стадии дробления в зависимости от верхнего предела крупности дроблённого продукта принято условно различать на:
Оборудование для дробления
Аппараты, в которых производится процесс дробления называются дробилками. Различают следующие типы дробилок:
В зависимости от производительности, условий работы, крупности питания, дробимости, наличия глинистых и илистых частиц в исходном питании, требуемого гранулометрического состава продукта дробления и других факторов на ОФ для крупного дробления применяют конусные, щековые дробилки и гирационные дробилки; для среднего дробления – конусные, валковые, молотковые; для мелкого дробления – конусные, валковые, ударного действия, с безударным ротором (центробежные однодисковые и центробежные многодисковые).
Дробилки могут работать в:
Если грохочение предшествует операции дробления, то оно называется предварительным, если осуществляется после дробления – поверочным (контрольным).
Схема дробления – графическое изображение последовательности операций при дроблении. Схемы дробления могут быть одно-, двух- и многостадиальными. Для предотвращения переизмельчения раскрытых рудных и нерудных минералов в схему дробления можно включать отдельные операции обогащения (межстадиальное обогащение).
Процесс дробления полезных ископаемых оценивается следующими показателями:
Перед дроблением угля необходимо предусмотреть устройство для удаления деревянных и металлических предметов, поступающих вместе с углём. Деревянные и другие предметы удаляются с ленточного конвейера. Для удаления из угля металлических предметов применяются электромагнитные шкивы и подвесные электромагнитные железоотделители.
Дробление негабаритов
Часто в рабочее пространство щековых дробилок попадают крупные куски округлой формы или куски, размер которых превышает ширину загрузочного отверстия. Крепкий кусок округлой формы не захватывается щеками и не дробится. Подвижная щека скользит по его поверхности и при приближении к неподвижной выталкивает его вверх. При отходе щеки кусок под тяжестью своей массы снова опускается вниз. Кусок большого размера может даже перекрыть загрузочное отверстие. В том и другом случае необходимо или принять меры, обеспечивающие прохождение куска через дробилку, т.е. раздробить его, или удалить этот кусок. Для этих целей изготавливают специальные устройства и инструменты:
При дроблении крепкого куска округлой формы при помощи клина, клин, подвешенный на длинный трос на крюк крана, опускается в щель между куском и неподвижной или подвижной щекой. В результате изменяется угол захвата куска между щеками, и кусок может быть раздроблен. Если использование клина не дает желаемых результатов или загрузочное отверстие дробилки перекрыто куском, имеющим размер больший размера загрузочного отверстия, то кусок поворачивают при помощи так называемых «кошек» или специально для этой цели изготовленных клещей. «Кошки» в виде крюка большого размера с зубьями или без них вырезают обычно из листа железа толщиной 30-50 мм и подвешивают на крюк крана при помощи троса. В том случае, когда раздробить кусок не удается, его удаляют из рабочего пространства, обвязав тросом, концы которого закрепляются на крюке крана.
Все работы по дроблению или удалению крупного куска могут производиться при работающей дробилке, но с обязательным соблюдением правил техники безопасности, предусмотренных в специально разработанной для выполнения этих работ инструкции. Поэтому в здании, где работает щековая дробилка, всегда должны быть в наличии исправный, готовый к работе кран и указанные приспособления.
В случае забивки рабочего пространства щековой дробилки мелким материалом очистка производится при остановленной дробилке с выполнением правил техники безопасности. Очистку производят сверху. Материал вручную нагружают в бадью с открывающимся днищем, которую при помощи крана поднимают из рабочего пространства и выгружают.
Очистка рабочего пространства конусной дробилки для среднего и мелкого дробления производится при помощи крючков и пик, специально изготовленных для этой цели. При спуске в рабочее пространство дробилки необходимо пользоваться предохранительными поясами. Загрузочные отверстия в этом случае перекрываются настилом для предотвращения падения на людей случайных предметов. Разбутовка, очистка и пуск дробилок после их аварийной остановки должны производиться по специальной инструкции, утвержденной главным инженером фабрики, а резка металла, застрявшего в рабочей зоне, – только под наблюдением лица технического надзора. Иногда удаление застрявших в рабочем пространстве дробилки кусков руды осуществляют с помощью взрыва, при этом необходимо соблюдать Правила безопасности при взрывных работах. Разбивать в рабочем пространстве дробилки куски руды молотками или кувалдами запрещается.
ЭМБРИОЛОГИЯ
ЭМБРИОЛОГИЯ, наука, изучающая развитие организма на самых ранних стадиях, предшествующих метаморфозу, вылуплению или рождению. Слияние гамет – яйца (яйцеклетки) и сперматозоида – с образованием зиготы дает начало новой особи, но прежде чем стать таким же существом, как родители, ей предстоит пройти определенные стадии развития: клеточное деление, образование первичных зародышевых листков и полостей, возникновение осей зародыша и осей симметрии, развитие целомических полостей и их производных, образование внезародышевых оболочек и, наконец, появление систем органов, функционально интегрированных и образующих тот или иной узнаваемый организм. Все это составляет предмет изучения эмбриологии.

Развитию предшествует гаметогенез, т.е. образование и созревание сперматозоида и яйца. Процесс развития всех яиц данного вида протекает в общем одинаково.
Гаметогенез.
Зрелые сперматозоид и яйцо различаются по своему строению, сходны у них только ядра; однако обе гаметы образуются из одинаковых на вид первичных половых клеток. У всех организмов, размножающихся половым путем, эти первичные половые клетки обособляются на ранних стадиях развития от других клеток и развиваются особым образом, готовясь к выполнению своей функции – продуцированию половых, или зародышевых, клеток. Поэтому их называют зародышевой плазмой – в отличие от всех других клеток, составляющих соматоплазму. Совершенно очевидно, однако, что и зародышевая плазма и соматоплазма происходят из оплодотворенного яйца – зиготы, давшей начало новому организму. Таким образом, в своей основе они одинаковы. Факторы, определяющие, какие клетки станут половыми, а какие – соматическими, до сих пор не установлены. Однако в конечном итоге половые клетки приобретают достаточно четкие отличия. Эти отличия возникают в процессе гаметогенеза.
У всех позвоночных и некоторых беспозвоночных первичные половые клетки возникают вдали от гонад и мигрируют к гонадам зародыша – яичнику или семеннику – с током крови, с пластами развивающихся тканей или посредством амебоидных движений. В гонадах из них образуются зрелые половые клетки. Ко времени развития гонад сома и зародышевая плазма функционально уже обособлены одна от другой, и, начиная с этого времени, на протяжении всей жизни организма половые клетки совершенно независимы от каких бы то ни было воздействий сомы. Именно поэтому признаки, приобретенные индивидом на протяжении его жизни, не оказывают влияния на его половые клетки.
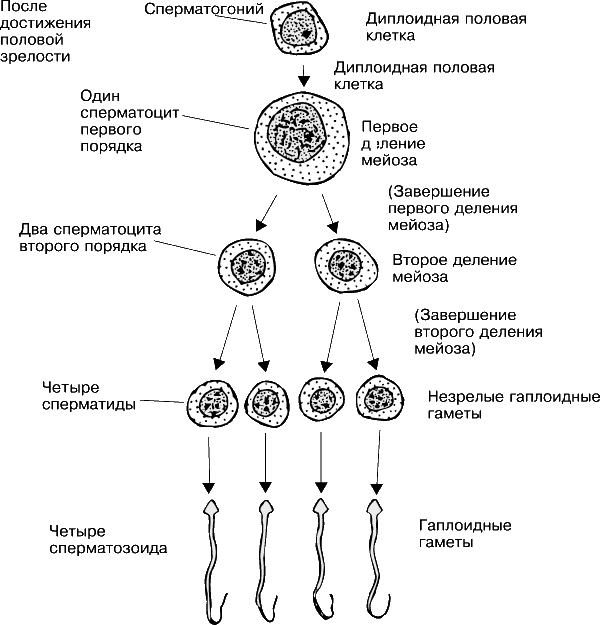
Первичные половые клетки, находясь в гонадах, делятся с образованием мелких клеток – сперматогониев в семенниках и оогониев в яичниках. Сперматогонии и оогонии продолжают многократно делиться, образуя клетки таких же размеров, что свидетельствует о компенсаторном росте как цитоплазмы, так и ядра. Сперматогонии и оогонии делятся митотически, и, следовательно, у них сохраняется первоначальное диплоидное число хромосом.
Спустя некоторое время эти клетки прекращают делиться и вступают в период роста, в течение которого в их ядрах происходят очень важные изменения. Хромосомы, полученные исходно от двух родителей, соединяются попарно (конъюгируют), вступая в очень тесное соприкосновение. Это делает возможным последующий кроссинговер (перекрест), в ходе которого гомологичные хромосомы разрываются и соединяются в новом порядке, обмениваясь эквивалентными участками; в результате кроссинговера в хромосомах оогониев и сперматогониев возникают новые комбинации генов. Предполагается, что стерильность мулов обусловлена несовместимостью хромосом, полученных от родителей – лошади и осла, из-за которой хромосомы не способны выжить при тесном соединении друг с другом. В результате созревание половых клеток в яичниках или семенниках мула прекращается на стадии конъюгации.
Когда ядро перестроилось и в клетке накопилось достаточное количество цитоплазмы, возобновляется процесс деления; вся клетка и ядро подвергаются двум разного типа делениям, определяющим собственно процесс созревания половых клеток. Одно из них – митоз – приводит к образованию клеток, аналогичных исходной; в результате другого – мейоза, или редукционного деления, в ходе которого клетки делятся дважды, – образуются клетки, каждая из которых содержит лишь половинное (гаплоидное) число хромосом по сравнению с исходным, а именно по одной из каждой пары (см. также КЛЕТКА). У некоторых видов эти клеточные деления происходят в обратном порядке. После роста и реорганизации ядер в оогониях и сперматогониях и непосредственно перед первым делением мейоза эти клетки получают названия ооцитов и сперматоцитов первого порядка, а после первого деления мейоза – ооцитов и сперматоцитов второго порядка. Наконец, после второго деления мейоза клетки, находящиеся в яичнике, называют яйцами (яйцеклетками), а находящиеся в семеннике – сперматидами. Теперь яйцо окончательно созрело, а сперматиде предстоит еще проделать метаморфоз и превратиться в сперматозоид.
Здесь необходимо подчеркнуть одно важное различие между оогенезом и сперматогенезом. Из одного ооцита первого порядка в результате созревания получается только одно зрелое яйцо; остальные три ядра и небольшое количество цитоплазмы превращаются в полярные тельца, которые не функционируют как половые клетки и в дальнейшем дегенерируют. Вся цитоплазма и желток, которые могли бы распределиться по четырем клеткам, концентрируются в одной – в зрелом яйце. В отличие от этого один сперматоцит первого порядка дает начало четырем сперматидам и такому же числу зрелых сперматозоидов, не теряя ни одного ядра. При оплодотворении восстанавливается диплоидное, или нормальное, число хромосом.
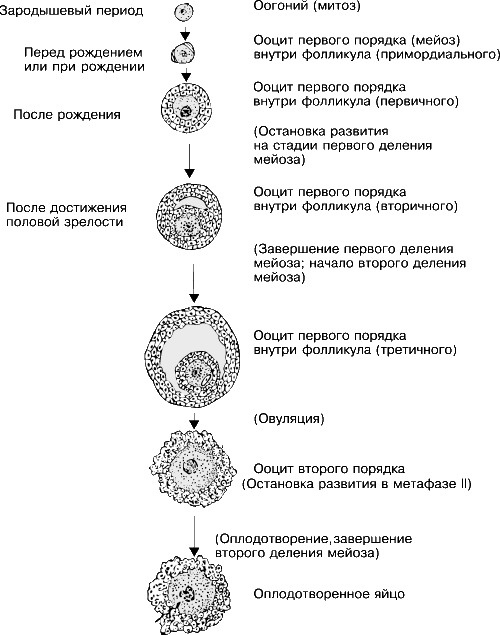
Яйцеклетка инертна и обычно крупнее соматических клеток данного организма. Яйцеклетка мыши составляет примерно 0,06 мм в диаметре, тогда как диаметр страусиного яйца бывает более 15 см. Яйца обычно имеют шаровидную или овальную форму, но бывают также продолговатыми, как у насекомых, миксины или ильной рыбы. Размеры и другие признаки яйца зависят от количества и распределения в нем питательного желтка, накапливающегося в виде гранул или, реже, в виде сплошной массы. Поэтому яйца делят на разные типы в зависимости от содержания в них желтка.
Гомолецитальные яйца
(от греч. homós – равный, однородный, lékithos – желток). В гомолецитальных яйцах, называемых также изолецитальными или олиголецитальными, желтка очень мало и он равномерно распределен в цитоплазме. Такие яйца типичны для губок, кишечнополостных, иглокожих, морских гребешков, нематод, оболочников и большинства млекопитающих.
Телолецитальные яйца
(от греч. télos – конец) содержат значительное количество желтка, а цитоплазма сконцентрирована у них на одном конце, обозначаемом обычно как анимальный полюс. Противоположный полюс, на котором сконцентрирован желток, называют вегетативным. Такие яйца типичны для кольчатых червей, головоногих моллюсков, бесчерепных (ланцетник), рыб, земноводных, пресмыкающихся, птиц и однопроходных млекопитающих. У них хорошо выражена анимально-вегетативная ось, определяемая градиентом распределения желтка; ядро обычно располагается эксцентрически; в яйцах, содержащих пигмент, он также распределяется по градиенту, но, в отличие от желтка, его больше на анимальном полюсе.
Центролецитальные яйца.
В них желток расположен в центре, так что цитоплазма сдвинута к периферии и дробление поверхностное. Такие яйца типичны для некоторых кишечнополостных и членистоногих.
Сперматозоид.
В отличие от крупной и инертной яйцеклетки, сперматозоиды мелкие, от 0,02 до 2,0 мм в длину, они активны и способны проплыть большое расстояние, чтобы добраться до яйца. Цитоплазмы в них мало, а желтка нет вообще.
Форма сперматозоидов разнообразна, однако среди них можно выделить два главных типа – жгутиковые и безжгутиковые. Безжгутиковые формы сравнительно редки. У большинства животных активная роль в оплодотворении принадлежит сперматозоиду. См. также СПЕРМАТОЗОИД.
Оплодотворение.
Оплодотворение – сложный процесс, в ходе которого сперматозоид проникает в яйцо и их ядра сливаются. В результате слияния гамет образуется зигота – по существу уже новая особь, способная развиваться при наличии необходимых для этого условий. Оплодотворение вызывает активацию яйца, стимулируя его к последовательным изменениям, приводящим к развитию сформированного организма. При оплодотворении происходит также амфимиксис, т.е. смешение наследственных факторов в результате слияния ядер яйца и сперматозоида. Яйцо обеспечивает половину необходимых хромосом и обычно все питательные вещества, необходимые для ранних стадий развития.
При соприкосновении сперматозоида с поверхностью яйца желточная оболочка яйца изменяется, превращаясь в оболочку оплодотворения. Это изменение считается доказательством того, что произошла активация яйца. Одновременно на поверхности яиц, содержащих мало желтка или не содержащих его вовсе, возникает т.н. кортикальная реакция, не допускающая проникновения в яйцо других сперматозоидов. У яиц, содержащих очень много желтка, кортикальная реакция возникает позднее, так что в них обычно проникает несколько сперматозоидов. Но даже в таких случаях оплодотворение совершает только один сперматозоид, первым дошедший до ядра яйца.
У некоторых яиц в месте соприкосновения сперматозоида с плазматической мембраной яйца образуется выпячивание мембраны – т.н. бугорок оплодотворения; он облегчает проникновение сперматозоида. Обычно в яйцо проникают головка сперматозоида и центриоли, находящиеся в его средней части, а хвост остается снаружи. Центриоли способствуют образованию веретена при первом делении оплодотворенного яйца. Процесс оплодотворения можно считать завершенным, когда два гаплоидных ядра – яйцеклетки и сперматозоида – сливаются и их хромосомы конъюгируют, готовясь к первому дроблению оплодотворенного яйца. См. также ЯЙЦО.
Дробление.
Если возникновение оболочки оплодотворения считается показателем активации яйца, то деление (дробление) служит первым признаком действительной активности оплодотворенного яйца. Характер дробления зависит от количества и распределения желтка в яйце, а также от наследственных свойств ядра зиготы и особенностей цитоплазмы яйца (последние целиком определяются генотипом материнского организма). Выделяют три типа дробления оплодотворенного яйца.
Голобластическое дробление
характерно для гомолецитальных яиц. Плоскости дробления разделяют яйцо полностью. Они могут делить его на равные части, как у морской звезды или морского ежа, или же на неравные части, как у брюхоногого моллюска Crepidula. Дробление умеренно телолецитального яйца ланцетника происходит по голобластическому типу, однако неравномерность деления проявляется только после стадии четырех бластомеров. У некоторых клеток после этой стадии дробление становится крайне неравномерным; образующиеся при этом мелкие клетки называют микромерами, а крупные клетки, содержащие желток, – макромерами. У моллюсков плоскости дробления проходят таким образом, что начиная со стадии восьми клеток бластомеры располагаются по спирали; этот процесс регулируется ядром.
Меробластическое дробление
типично для телолецитальных яиц, богатых желтком; оно ограничено относительно небольшим участком у анимального полюса. Плоскости дробления не проходят через все яйцо и не захватывают желток, так что в результате деления на анимальном полюсе образуется небольшой диск клеток (бластодиск). Такое дробление, называемое также дискоидальным, свойственно пресмыкающимся и птицам.
Поверхностное дробление
типично для центролецитальных яиц. Ядро зиготы делится в центральном островке цитоплазмы, и получающиеся при этом клетки перемещаются на поверхность яйца, образуя поверхностный слой клеток вокруг лежащего в центре желтка. Этот тип дробления наблюдается у членистоногих.
Правила дробления.
Установлено, что дробление подчиняется определенным правилам, названным именами исследователей, которые их впервые сформулировали. Правило Пфлюгера: веретено всегда тянется в направлении наименьшего сопротивления. Правило Бальфура: скорость голобластического дробления обратно пропорциональна количеству желтка (желток затрудняет деление как ядра, так и цитоплазмы). Правило Сакса: клетки обычно делятся на равные части, и плоскость каждого нового деления пересекает плоскость предшествующего деления под прямым углом. Правило Гертвига: ядро и веретено обычно располагаются в центре активной протоплазмы. Ось каждого веретена деления располагается по длинной оси массы протоплазмы. Плоскости деления обычно пересекают массу протоплазмы под прямым углом к ее осям.
В результате дробления оплодотворенных яиц любого типа образуются клетки, называемые бластомерами. Когда бластомеров становится много (у земноводных, например, от 16 до 64 клеток), они образуют структуру, напоминающую ягоду малины и названную морулой.
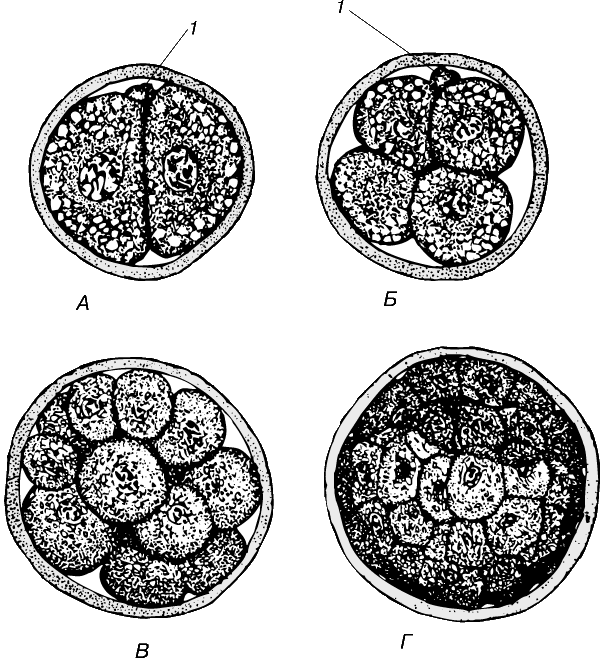
Бластула.
По мере продолжения дробления бластомеры становятся все мельче и все плотнее прилегают друг к другу, приобретая гексагональную форму. Такая форма повышает структурную жесткость клеток и плотность слоя. Продолжая делиться, клетки раздвигают друг друга и в итоге, когда их число достигает нескольких сотен или тысяч, формируют замкнутую полость – бластоцель, в который поступает жидкость из окружающих клеток. В целом это образование носит название бластулы. Ее формированием (в котором клеточные движения не участвуют) завершается период дробления яйца.
В гомолецитальных яйцах бластоцель может располагаться в центре, но в телолецитальных яйцах он обычно бывает сдвинут желтком и располагается эксцентрически, ближе к анимальному полюсу и прямо под бластодиском. Итак, бластула обычно представляет собой полый шарик, полость которого (бластоцель) заполнена жидкостью, но в телолецитальных яйцах с дискоидальным дроблением бластула представлена уплощенной структурой.
При голобластическом дроблении стадия бластулы считается завершенной, когда в результате деления клеток соотношение между объемами их цитоплазмы и ядра становится таким же, как в соматических клетках. В оплодотворенном яйце объемы желтка и цитоплазмы совершенно не соответствуют размерам ядра. Однако в процессе дробления количество ядерного материала несколько увеличивается, тогда как цитоплазма и желток только делятся. В некоторых яйцах отношение объема ядра к объему цитоплазмы в момент оплодотворения составляет примерно 1:400, а к концу стадии бластулы – примерно 1:7. Последнее близко к соотношению, характерному и для первичной половой и для соматической клетки.
Поверхности поздней бластулы оболочников и земноводных можно картировать; для этого на разные ее участки наносят прижизненные (не наносящие вреда клеткам) красители – сделанные цветные метки сохраняются в ходе дальнейшего развития и позволяют установить, какие органы возникают из каждого участка. Эти участки называют презумптивными, т.е. такими, судьбу которых при нормальных условиях развития можно предсказать. Если, однако, на стадии поздней бластулы или ранней гаструлы переместить эти участки или поменять местами, их судьба изменится. Подобные эксперименты показывают, что до какой-то определенной стадии развития каждый бластомер способен превратиться в любую из множества разнообразных клеток, составляющих организм.
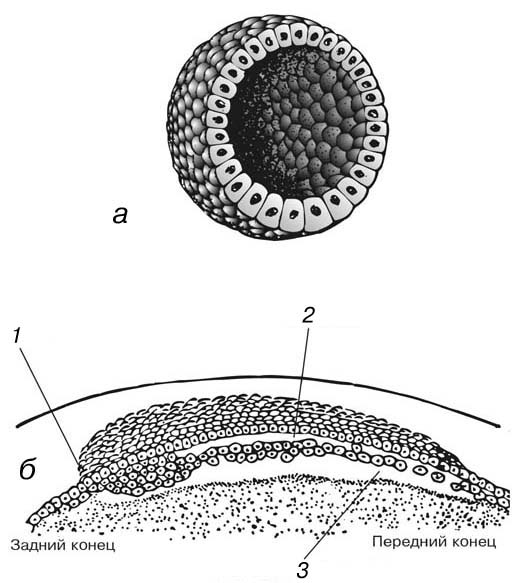
Гаструла.
Гаструлой называют стадию эмбрионального развития, на которой зародыш состоит из двух слоев: наружного – эктодермы, и внутреннего – энтодермы. У разных животных эта двуслойная стадия достигается разными способами, поскольку яйца разных видов содержат разное количество желтка. Однако в любом случае главную роль в этом играют перемещения клеток, а не клеточные деления.
Инвагинация.
В гомолецитальных яйцах, для которых типично голобластическое дробление, гаструляция обычно происходит путем инвагинации (впячивания) клеток вегетативного полюса, что приводит к образованию двуслойного зародыша, имеющего форму чаши. Первоначальный бластоцель сокращается, но при этом образуется новая полость – гастроцель. Отверстие, ведущее в этот новый гастроцель, называется бластопором (название неудачное, поскольку оно открывается не в бластоцель, а в гастроцель). Бластопор расположен в области будущего анального отверстия, на заднем конце зародыша, и в этой области развивается большая часть мезодермы – третьего, или среднего, зародышевого листка. Гастроцель называют также архентероном, или первичной кишкой, и он служит зачатком пищеварительной системы.
Инволюция.
У пресмыкающихся и птиц, телолецитальные яйца которых содержат большое количество желтка и дробятся меробластически, клетки бластулы на очень небольшом участке приподнимаются над желтком и затем начинают вворачиваться внутрь, под клетки верхнего слоя, образуя второй (нижний) слой. Этот процесс вворачивания клеточного пласта называют инволюцией. Верхний слой клеток становится наружным зародышевым листком, или эктодермой, а нижний – внутренним, или энтодермой. Эти слои переходят один в другой, а место, где происходит переход, известно под названием губы бластопора. Крыша первичной кишки у зародышей этих животных состоит из вполне сформировавшихся энтодермальных клеток, а дно – из желтка; дно из клеток образуется позднее.
Деламинация.
У высших млекопитающих, в том числе у человека, гаструляция происходит несколько иначе, а именно путем деламинации, но приводит к тому же результату – образованию двуслойного зародыша. Деламинация – это расслоение исходного наружного слоя клеток, приводящее к возникновению внутреннего слоя клеток, т.е. энтодермы.
Вспомогательные процессы.
Результаты гаструляции.
Конечный результат гаструляции заключается в образовании двуслойного зародыша. Наружный слой зародыша (эктодерма) образован мелкими, часто – пигментированными клетками, не содержащими желтка; из эктодермы в дальнейшем развиваются такие ткани, как, например, нервная, и верхние слои кожи. Внутренний слой (энтодерма) состоит из почти не пигментированных клеток, сохраняющих некоторое количество желтка; они дают начало главным образом тканям, выстилающим пищеварительный тракт и его производные. Следует, однако, подчеркнуть, что глубоких различий между этими двумя зародышевыми листками не существует. Эктодерма дает начало энтодерме, и если у некоторых форм границу между ними в области губы бластопора можно определить, то у других она практически неразличима. В экспериментах по трансплантации было показано, что различие между этими тканями определяется только их местоположением. Если участки, которые в норме оставались бы эктодермальными и дали бы начало производным кожи, пересадить на губу бластопора, они вворачиваются внутрь и становятся энтодермой, которая может превратиться в выстилку пищеварительного тракта, легкие или щитовидную железу.
Часто с появлением первичной кишки центр тяжести зародыша смещается, он начинает поворачиваться в своих оболочках, и в нем впервые устанавливаются передне-задняя (голова – хвост) и дорсо-вентральная (спина – живот) оси симметрии будущего организма.
Зародышевые листки.
Эктодерму, энтодерму и мезодерму различают на основании двух критериев. Во-первых, по их местоположению в зародыше на ранних стадиях его развития: в этот период эктодерма всегда расположена снаружи, энтодерма – внутри, а мезодерма, появляющаяся последней, – между ними. Во-вторых, по их будущей роли: каждый из этих листков дает начало определенным органам и тканям, и их нередко идентифицируют по их дальнейшей судьбе в процессе развития. Однако напомним, что в период возникновения этих листков никаких принципиальных различий между ними не существует. В опытах по пересадке зародышевых листков было показано, что первоначально каждый из них обладает потенциями любого из двух других. Таким образом, их разграничение искусственно, но им очень удобно пользоваться при изучении эмбрионального развития.
Мезодерма, т.е. средний зародышевый листок, образуется несколькими способами. Она может возникать непосредственно из энтодермы путем образования целомических мешков, как у ланцетника; одновременно с энтодермой, как у лягушки; или путем деламинации, из эктодермы, как у некоторых млекопитающих. В любом случае вначале мезодерма представляет собой слой клеток, лежащих в пространстве, которое первоначально занимал бластоцель, т.е. между эктодермой с наружной и энтодермой с внутренней стороны.
Мезодерма вскоре расщепляется на два клеточных слоя, между которыми образуется полость, называемая целомом. Из этой полости в последующем образуются полость перикарда, окружающая сердце, плевральная полость, окружающая легкие, и брюшная полость, в которой лежат органы пищеварения. Наружный слой мезодермы – соматическая мезодерма – образует вместе с эктодермой т.н. соматоплевру. Из наружной мезодермы развиваются поперечнополосатые мышцы туловища и конечностей, соединительная ткань и сосудистые элементы кожи. Внутренний слой мезодермальных клеток называется спланхнической мезодермой и вместе с энтодермой образует спланхноплевру. Из этого слоя мезодермы развиваются гладкие мышцы и сосудистые элементы пищеварительного тракта и его производных. В развивающемся зародыше много рыхлой мезенхимы (эмбриональной мезодермы), заполняющей пространство между эктодермой и энтодермой.
У хордовых в процессе развития образуется продольный столбик плоских клеток – хорда, основной отличительный признак этого типа. Клетки хорды происходят из эктодермы у одних животных, из энтодермы у других и из мезодермы у третьих. В любом случае эти клетки уже на очень ранней стадии развития можно отличить от остальных, и расположены они в виде продольного столбика над первичной кишкой. У зародышей позвоночных хорда служит центральной осью, вокруг которой развивается осевой скелет, а над ней – центральная нервная система. У большинства хордовых это чисто эмбриональная структура, и только у ланцетника, круглоротых и пластиножаберных она сохраняется в течение всей жизни. Почти у всех других позвоночных клетки хорды замещаются костными клетками, образующими тело развивающихся позвонков; из этого следует, что наличие хорды облегчает формирование позвоночного столба.
Производные зародышевых листков.
Дальнейшая судьба трех зародышевых листков различна.
Из эктодермы развиваются: вся нервная ткань; наружные слои кожи и ее производные (волосы, ногти, зубная эмаль) и частично слизистая ротовой полости, полостей носа и анального отверстия.
Энтодерма дает начало выстилке всего пищеварительного тракта – от ротовой полости до анального отверстия – и всем ее производным, т.е. тимусу, щитовидной железе, паращитовидным железам, трахее, легким, печени и поджелудочной железе.
Из мезодермы образуются: все виды соединительной ткани, костная и хрящевая ткани, кровь и сосудистая система; все типы мышечной ткани; выделительная и репродуктивная системы, дермальный слой кожи.
У взрослого животного очень мало таких органов энтодермального происхождения, которые не содержали бы нервных клеток, происходящих из эктодермы. В каждом важном органе содержатся и производные мезодермы – кровеносные сосуды, кровь, часто и мышцы, так что структурная обособленность зародышевых листков сохраняется только на стадии их образования. Уже в самом начале своего развития все органы приобретают сложное строение, и в них входят производные всех зародышевых листков.
ОБЩИЙ ПЛАН СТРОЕНИЯ ТЕЛА
Симметрия.
На ранних стадиях развития организм приобретает определенный тип симметрии, характерный для данного вида. Один из представителей колониальных протистов, вольвокс, обладает центральной симметрией: любая плоскость, проходящая через центр вольвокса, делит его на две равноценные половины. Среди многоклеточных нет ни одного животного, обладающего симметрией такого типа. Для кишечнополостных и иглокожих характерна радиальная симметрия, т.е. части их тела расположены вокруг главной оси, образуя как бы цилиндр. Некоторые, но не все плоскости, проходящие через эту ось, делят такое животное на две равноценные половинки. Все иглокожие на личиночной стадии обладают двусторонней симметрией, но в процессе развития приобретают радиальную симметрию, характерную для взрослой стадии.
Для всех высокоорганизованных животных типична двусторонняя симметрия, т.е. их можно разделить на две симметричные половины только в одной плоскости. Поскольку такое расположение органов наблюдается у большинства животных, его считают оптимальным для выживания. Плоскость, проходящая по продольной оси от вентральной (брюшной) к дорсальной (спинной) поверхности, делит животное на две половины, правую и левую, являющиеся зеркальными отображениями друг друга.
Почти все неоплодотворенные яйца обладают радиальной симметрией, но некоторые теряют ее в момент оплодотворения. Например, в яйце лягушки место проникновения сперматозоида всегда сдвинуто к переднему, или головному, концу будущего зародыша. Эта симметрия определяется только одним фактором – градиентом распределения желтка в цитоплазме.
Двусторонняя симметрия становится очевидной, как только в ходе эмбрионального развития начинается формирование органов. У высших животных практически все органы закладываются попарно. Это относится к глазам, ушам, ноздрям, легким, конечностям, большинству мышц, частей скелета, кровеносных сосудов и нервов. Даже сердце закладывается в виде парной структуры, а затем ее части сливаются, образуя один трубчатый орган, который впоследствии перекручивается, превращаясь в сердце взрослой особи с его сложной структурой. Неполное слияние правой и левой половинок органов проявляется, например, в случаях расщелины нёба или заячьей губы, изредка встречающихся у человека.
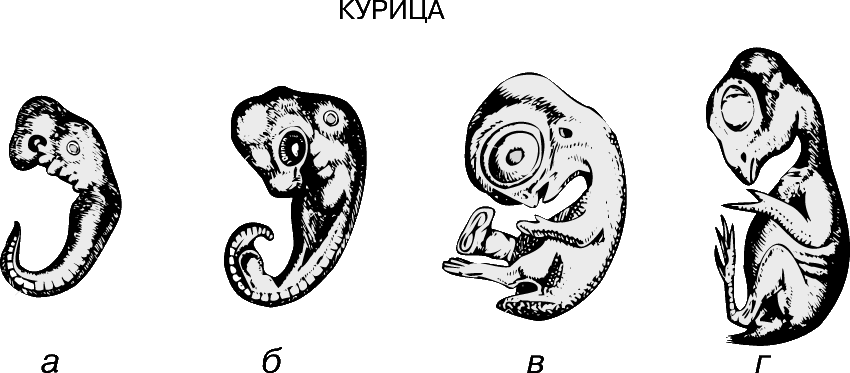
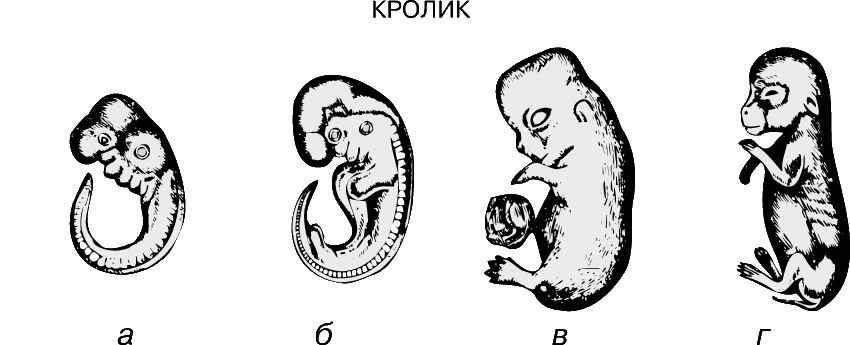
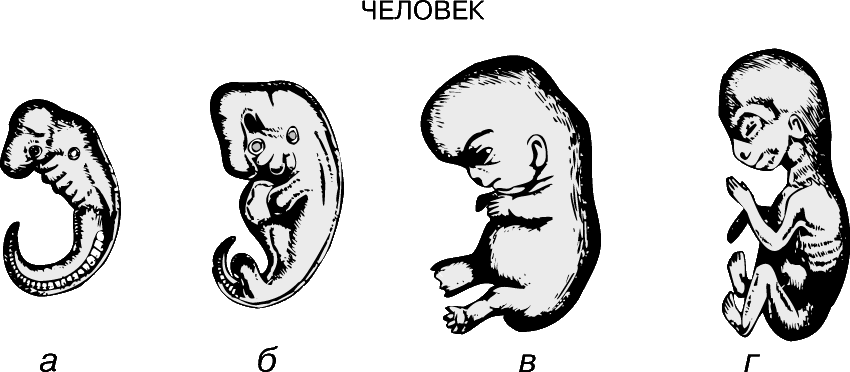
Метамерия (расчленение тела на сходные сегменты).
Наибольшего успеха в длительном процессе эволюции достигли животные с сегментированным телом. Метамерное строение кольчатых червей и членистоногих отчетливо видно на протяжении всей их жизни. У большинства позвоночных первоначально сегментированное строение в дальнейшем становится мало различимым, однако на эмбриональных стадиях метамерия у них ясно выражена.
У ланцетника метамерия проявляется в строении целома, мышц и гонад. Для позвоночных характерно сегментарное расположение некоторых частей нервной, выделительной, сосудистой и опорной систем; однако уже на ранних стадиях эмбрионального развития на эту метамерию накладывается опережающее развитие переднего конца тела – т.н. цефализация. Если рассмотреть выращенного в инкубаторе 48-часового зародыша цыпленка, то можно выявить у него одновременно и двустороннюю симметрию и метамерию, наиболее отчетливо выраженную на переднем конце тела. Например, группы мышц, или сомиты, сначала появляются в области головы и образуются последовательно, так что наименее развитыми сегментированными сомитами оказываются задние.
Органогенез.
У большинства животных одним из первых дифференцируется пищеварительный канал. В сущности, зародыши большинства животных представляют собой трубку, вставленную в другую трубку; внутренняя трубка – это кишка, от ротового до анального отверстия. Другие органы, входящие в систему пищеварения, и органы дыхания закладываются в виде выростов этой первичной кишки. Присутствие крыши архентерона, или первичной кишки, под дорсальной эктодермой вызывает (индуцирует), возможно совместно с хордой, образование на спинной стороне зародыша второй важнейшей системы организма, а именно центральной нервной системы. Это происходит следующим образом: сначала утолщается дорсальная эктодерма и образуется нервная пластинка; затем края нервной пластинки приподнимаются, образуя нервные валики, которые растут навстречу друг другу и в конечном счете смыкаются, – в результате возникает нервная трубка, зачаток центральной нервной системы. Из передней части нервной трубки развивается головной мозг, а остальная ее часть превращается в спинной мозг. Полость нервной трубки по мере разрастания нервной ткани почти исчезает – от нее остается лишь узкий центральный канал. Головной мозг формируется в результате выпячиваний, впячиваний, утолщений и утоньшений передней части нервной трубки зародыша. От образовавшегося головного и спинного мозга берут начало парные нервы – черепные, спинномозговые и симпатические.
Мезодерма тоже претерпевает изменения сразу после своего возникновения. Она образует парные и метамерные сомиты (блоки мышц), позвонки, нефротомы (зачатки органов выделения) и части репродуктивной системы.
Таким образом, развитие систем органов начинается сразу после образования зародышевых листков. Все процессы развития (при нормальных условиях) происходят с точностью самых совершенных технических устройств.
МЕТАБОЛИЗМ ЗАРОДЫШЕЙ
Зародышам, развивающимся в водной среде, не требуется иных покровов, кроме студнеобразных оболочек, покрывающих яйцо. Эти яйца содержат достаточное количество желтка, чтобы обеспечить зародыш питанием; оболочки до некоторой степени защищают его и помогают сохранять метаболическое тепло и вместе с тем достаточно проницаемы, чтобы не препятствовать свободному газообмену (т.е. поступлению кислорода и выходу диоксида углерода) между зародышем и средой.
Внезародышевые оболочки.
У животных, откладывающих яйца на суше или живородящих, зародышу необходимы дополнительные оболочки, защищающие его от обезвоживания (если яйца откладываются на суше) и обеспечивающие питание, удаление конечных продуктов обмена и газообмен.
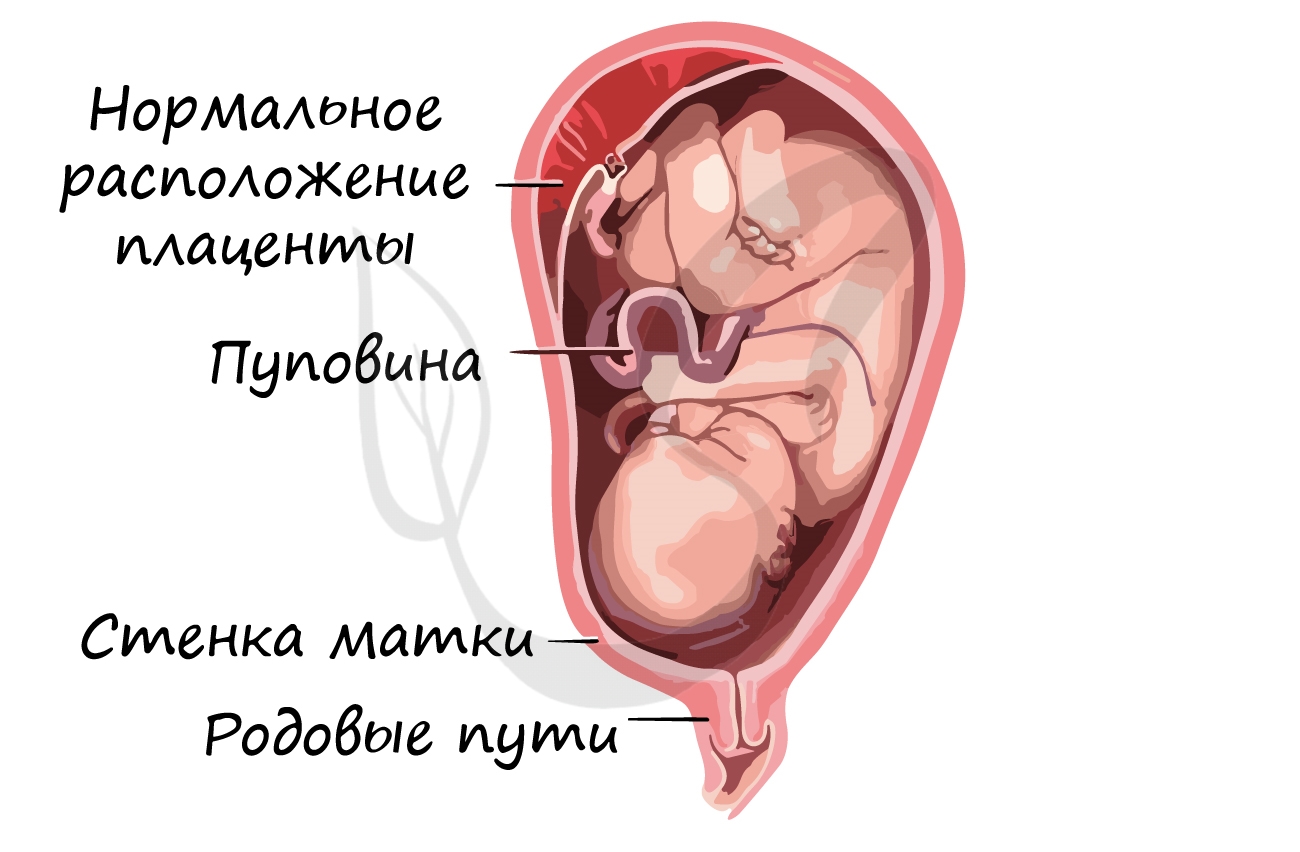
Эти функции выполняют внезародышевые оболочки – амнион, хорион, желточный мешок и аллантоис, образующиеся в процессе развития у всех пресмыкающихся, птиц и млекопитающих. Хорион и амнион тесно связаны между собой по происхождению; они развиваются из соматической мезодермы и эктодермы. Хорион – самая наружная оболочка, окружающая зародыш и три другие оболочки; эта оболочка проницаема для газов и через нее происходит газообмен. Амнион предохраняет клетки зародыша от высыхания благодаря амниотической жидкости, секретируемой его клетками. Желточный мешок, наполненный желтком, вместе с желточным стебельком поставляет зародышу подвергшиеся перевариванию питательные вещества; эта оболочка содержит густую сеть кровеносных сосудов и клетки, вырабатывающие пищеварительные ферменты. Желточный мешок, как и аллантоис, образуется из спланхнической мезодермы и энтодермы: энтодерма и мезодерма распространяются по всей поверхности желтка, обрастая его, так что в конце концов весь желток оказывается в желточном мешке. У пресмыкающихся и птиц аллантоис служит резервуаром для конечных продуктов обмена, поступающих из почек зародыша, а также обеспечивает газообмен. У млекопитающих эти важные функции выполняет плацента – сложный орган, образуемый ворсинками хориона, которые, разрастаясь, входят в углубления (крипты) слизистой оболочки матки, где вступают в тесный контакт с ее кровеносными сосудами и железами.
У человека плацента полностью обеспечивает дыхание зародыша, питание и выделение продуктов обмена в кровоток матери.
Внезародышевые оболочки не сохраняются в постэмбриональном периоде. У пресмыкающихся и птиц при вылуплении высохшие оболочки остаются в скорлупе яйца. У млекопитающих плацента и остальные внезародышевые оболочки выбрасываются из матки (отторгаются) после рождения плода. Эти оболочки обеспечили высшим позвоночным независимость от водной среды и, несомненно, сыграли важную роль в эволюции позвоночных, особенно в возникновении млекопитающих.
БИОГЕНЕТИЧЕСКИЙ ЗАКОН
В 1828 К.фон Бэр сформулировал следующие положения: 1) наиболее общие признаки любой крупной группы животных появляются у зародыша раньше, чем менее общие признаки; 2) после формирования самых общих признаков появляются менее общие и так до появления особых признаков, свойственных данной группе; 3) зародыш любого вида животных по мере развития становится все менее похожим на зародышей других видов и не проходит через поздние стадии их развития; 4) зародыш высокоорганизованного вида может обладать сходством с зародышем более примитивного вида, но никогда не бывает похож на взрослую форму этого вида.
Биогенетический закон, сформулированный в этих четырех положениях, часто истолковывают неверно. Закон этот просто утверждает, что некоторые стадии развития высокоорганизованных форм обладают явным сходством с некоторыми стадиями развития нижестоящих на эволюционной лестнице форм. Предполагается, что это сходство можно объяснить происхождением от общего предка. О взрослых стадиях низших форм ничего не говорится. В данной статье сходство между зародышевыми стадиями подразумевается; в противном случае развитие каждого вида пришлось бы описывать отдельно.
По-видимому, в длительной истории жизни на Земле среда играла главную роль в отборе зародышей и взрослых организмов, наиболее приспособленных для выживания. Узкие рамки, создаваемые средой в отношении возможных колебаний температуры, влажности и снабжения кислородом, сокращали разнообразие форм, приводя их к относительно общему типу. В результате возникло то сходство строения, которое лежит в основе биогенетического закона, если речь идет о зародышевых стадиях. Разумеется, у ныне существующих форм в процессе зародышевого развития проявляются особенности, соответствующие времени, месту и способам размножения данного вида.
Онтогенез, т.е. развитие отдельной особи, предваряет филогенез, т.е. развитие группы, потому что мутации обычно возникают в половых клетках до оплодотворения. Изменения в эмбрионе естественно предшествуют изменениям взрослой особи, имеющим эволюционное значение, а часто и вызывают их. Новая особь «закладывается» в момент оплодотворения, а зародышевое развитие только подготавливает его к превратностям взрослого существования и созданию будущих зародышей. См. также ЦИТОЛОГИЯ; НАСЛЕДСТВЕННОСТЬ; СИСТЕМАТИКА ЖИВОТНЫХ.



