ДЫХАТЕЛЬНАЯ ФУНКЦИЯ КРОВИ. Перенос кислорода кровью
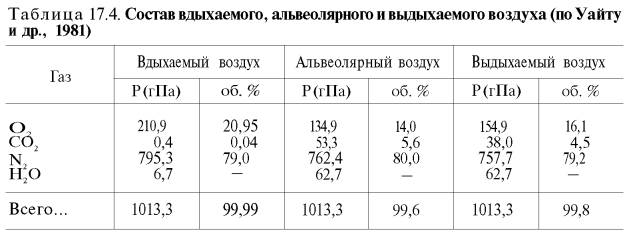
Сущность дыхательной функции крови состоит в доставке кислорода от легких к тканям и углекислого газа от тканей к легким (табл. 17.4).
Кровь осуществляет дыхательную функцию прежде всего благодаря наличию в ней гемоглобина. Физиологическая функция гемоглобина как переносчика кислорода основана на способности обратимо связывать кислород. Поэтому в легочных капиллярах происходит насыщение крови кислородом, а в тканевых капиллярах, где парциальное давление кислорода резко снижено, осуществляется отдача кислорода тканям.
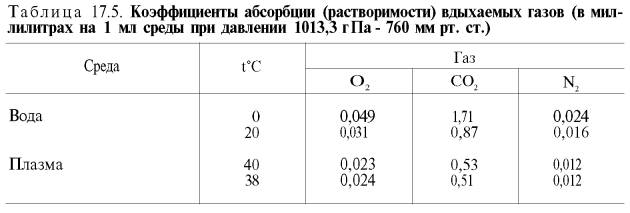
В состоянии покоя ткани и органы человека потребляют около 200 мл кислорода в минуту. При тяжелой физической работе количество потребляемого тканями кислорода возрастает в 10 раз и более (до 2–3 л/мин). Доставка от легких к тканям такого количества кислорода в виде газа, физически растворенного в плазме, невозможна вследствие малой растворимости кислорода в воде и плазме крови (табл. 17.5).
Исходя из приведенных в табл. 17.5 данных, а также зная РO2 в артериальной крови – 107–120 гПа (80–90 мм рт. ст.), нетрудно видеть, что количество физически растворенного кислорода в плазме крови не может превышать 0,3 об. %. При расчете кислородной емкости крови этой величиной можно пренебречь.
Итак, функцию переносчика кислорода в организме выполняет гемоглобин. Напомним, что молекула гемоглобина построена из 4 субъединиц (полипептидных цепей), каждая из которых связана с гемом (см. главу 2). Следовательно, молекула гемоглобина имеет 4 гема, к которым может присоединяться кислород, при этом гемоглобин переходит в оксигемо-глобин.
Гемоглобин человека содержит 0,335% железа. Каждый грамм-атом железа (55,84 г) в составе гемоглобина при полном насыщении кислородом связывает 1 грамм-молекулу кислорода (22400 мл). Таким образом, 100 г гемоглобина могут связывать
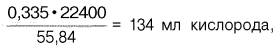
а каждый грамм гемоглобина – 1,34 мл кислорода. Содержание гемоглобина в крови здорового человека составляет 13–16%, т.е. в 100 мл крови 13–16 г гемоглобина. При РО2 в артериальной крови 107–120 гПа гемоглобин насыщен кислородом на 96%. Следовательно, в этих условиях 100 мл крови содержит 19–20 об. % кислорода:

В венозной крови в состоянии покоя РО2 = 53,3 гПа, и в этих условиях гемоглобин насыщен кислородом лишь на 70–72%, т.е. содержание кислорода в 100 мл венозной крови не превышает
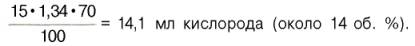
Артериовенозная разница по кислороду будет около 6 об. %. Таким образом, за 1 мин ткани в состоянии покоя получают 200–240 мл кислорода (при условии, что минутный объем сердца в покое составляет 4 л).
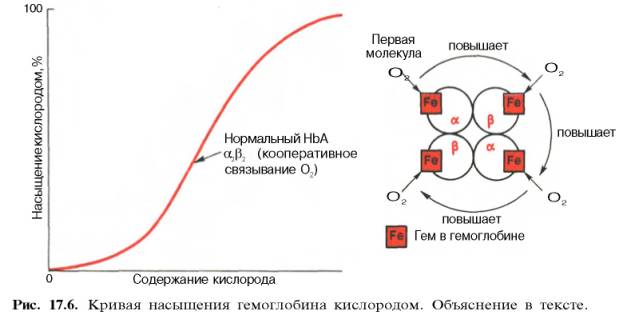
Возрастание интенсивности окислительных процессов в тканях, например при усиленной мышечной работе всегда связано с более полным извлечением кислорода из крови. Кроме того, при физической работе резко увеличивается скорость кровотока. Зависимость между степенью насыщения гемоглобина кислородом и РО2, можно выразить в виде кривой насыщения гемоглобина кислородом, или кривой диссоциации оксигемоглобина, которая имеет S-образную форму и характеризует сродство гемоглобина к кислороду (рис. 17.6).
Характерная для гемоглобина S-образная кривая насыщения кислородом свидетельствует, что связывание первой молекулы кислорода одним из
гемов гемоглобина облегчает связывание последующих молекул кислорода тремя другими оставшимися гемами. Долгое время механизм, лежащий в основе этого эффекта, оставался загадкой, так как, по данным рентгено-структурного анализа, 4 гема в молекуле гемоглобина довольно далеко отстоят друг от друга и вряд ли могут оказывать взаимное влияние. В последнее время принято следующее объяснение происхождения S-образ-ной кривой. Считают, что тетрамерная молекула гемоглобина способна обратимо распадаться на две половинки, каждая из которых содержит одну α-цепь и одну β-цепь:
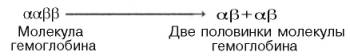
При взаимодействии молекулы кислорода с одним из четырех гемов гемоглобина кислород присоединяется к одной из половинок молекулы гемоглобина (допустим, к α-цепи этой половинки). Как только такое присоединение произойдет, α-полипептидная цепь претерпевает конформа-ционные изменения, которые передаются на тесно связанную с ней β-цепь; последняя также подвергается конформационным сдвигам. β-Цепь присоединяет кислород, имея уже большее сродство к нему. Таким путем связывание одной молекулы кислорода благоприятствует связыванию второй молекулы (так называемое кооперативное взаимодействие).
После насыщения кислородом одной половины молекулы гемоглобина возникает новое, внутреннее, напряженное состояние молекулы гемоглобина, которое вынуждает и вторую половину гемоглобина изменить конфор-мацию. Теперь еще две молекулы кислорода, по-видимому, по очереди связываются со второй половинкой молекулы гемоглобина, образуя оксигемоглобин.
S-образная форма кривой насыщения гемоглобина кислородом имеет большое физиологическое значение. При такой форме кривой обеспечивается возможность насыщения крови кислородом при изменении РО2 в довольно широких пределах. Например, дыхательная функция крови существенно не нарушается при снижении РО2 в альвеолярном воздухе со 133,3 до 80–93,3 гПа. Поэтому подъем на высоту до 3,0–3,5 км над уровнем моря не сопровождается развитием выраженной гипоксемии.
Численно сродство гемоглобина к кислороду принято выражать величиной Р50 – парциальное напряжение кислорода, при котором 50% гемоглобина связано с кислородом (рН 7,4 температура 37°С). Нормальная величина Р50 около 34,67 гПа (см. рис. 17.6). Смещение кривой насыщения гемоглобина кислородом вправо означает уменьшение способности гемоглобина связывать кислород и, следовательно, сопровождается повышением Р50. Напротив, смещение кривой влево свидетельствует о повышенном сродстве гемоглобина к кислороду, величина Р50 снижена.
Ход кривой насыщения гемоглобина кислородом или диссоциации оксигемоглобина зависит от ряда факторов. Сродство гемоглобина к кислороду в первую очередь связано с рН. Чем ниже рН, тем меньше способность гемоглобина связывать кислород и тем выше Р50. В тканевых капиллярах рН ниже (поступает большое количество СО2), в связи с чем гемоглобин легко отдает кислород. В легких СО2 выделяется, рН повышается и гемоглобин активно присоединяет кислород.
Способность гемоглобина связывать кислород зависит также от температуры. Чем выше температура (в тканях температура выше, чем в легких), тем меньше сродство гемоглобина к кислороду. Напротив, снижение температуры вызывает обратные явления.
Количество гемоглобина в крови, а также в какой-то мере его способность связывать кислород (характер кривой диссоциации оксигемоглобина) несколько меняются с возрастом. Например, у новорожденных содержание гемоглобина доходит до 20–21% (вместо обычных для взрослого 13–16%). У человека имеется несколько гемоглобинов, которые образуются в различном количестве в разные стадии онтогенеза и различаются по своему сродству к кислороду.
Рассмотрим нарушения дыхательной функции крови при некоторых патологических состояниях.
Анемия. Кровь и бескровие
Введение
Кровь представляет собой ткань, которая находится в постоянном движении. Даже когда человек абсолютно неподвижен, сердце его продолжает ритмично сокращаться, легкие совершают дыхательные циклы, в нескольких местах осуществляется волнообразная перистальтика – но все эти органы находятся на своих фиксированных анатомических позициях. А вот кровь непрерывно струится по… впрочем, не будем повторять банальности вслед за лириками: кровь, конечно, струится не по просторечным и устаревшим «жилам», а по артериям, артериолам, венам, венулам, артериоло-венулярным анастомозам и капиллярам, да и не так чтоб она прямо уж «струилась», «закипала» или, скажем, «стыла» – нет, кровь дискретными толчками перемещается под действием перепадов давления, создаваемых насосной функцией миокарда. Подвижные среды (кровь, лимфа) внутри неподвижной структуры – поистине удивительное изобретение природы, позже скопированное человеком в бесчисленных машинах с механизмом циркуляции горюче-смазочных, абсорбирующих, охлаждающих материалов.
О составе, биологическом значении и функциях крови можно говорить очень долго, тем более что мы, по-видимому, не всё еще об этом знаем. Уже выяснили, однако, что кровь более чем наполовину состоит из жидкой плазмы (которая, в свою очередь, содержит 90% воды), а сухой остаток – это т.н. форменные элементы, разноцветные клетки эритроциты, лейкоциты и тромбоциты; знаем, что в крови растворен сложнейший коктейль из протеинов, витаминов, гормонов, нейромедиаторов, иммуноглобулинов, ферментов, газов, аминокислот, электролитов, сахаров, солей и бог знает чего еще. Мы понимаем, что кровь одновременно является тканью и органом иммунной, дыхательной, энергообменной, метаболической, эндокринной, экскреторной, репаративной, репродуктивной, терморегуляторной и, в целом, гомеостатической систем (пусть даже некоторые из этих названий не академичны, а придуманы ради связности изложения). Именно благодаря кровообращению мы имеем возможность травить сразу весь организм этанолом, усилителями вкуса и другими ядами, а затем эффективно лечиться пилюлями и инъекциями; благодаря гемодинамике и гидравлическому шедевру пещеристых тел мы можем рассчитывать на то, что некоторые органы изменят конфигурацию и размеры в соответствии с обстоятельствами.
Однако человек давно знает и другое: чем больше функций, тем больше дисфункций. Чем сложнее состав, тем он нестабильнее. Как и любая другая ткань, кровь подвержена многочисленным болезням; почти все они клинически тяжелы, почти все трудны в изучении, понимании и лечении. Но все же они поддаются, и можно надеяться, что медицинская наука гематология в своем быстром развитии принесет нам качественно новые, революционные знания о том, что такое кровь и как это работает. Во всяком случае, тяжелобольным кровь больше не «отворяют», а вот спасительная гемотрансфузия применяется повсеместно.
Ниже речь пойдет о самой обширной и распространенной группе заболеваний крови – об анемиях. Сегодня даже за пределами медицины анемию «малокровием» называют все реже, – потому что это не совсем и не всегда малокровие; так можно было бы назвать, скорее, гиповолемию (сокращение объема циркулирующей крови), а «анемия» с древнегреческого должна переводиться, по идее, как полное отсутствие крови, бескровие, обескровленность.
Однако анемия – и не бескровие тоже. Если уж впадать в грех неологизма, то это «малоэритроцитье» или «безгемоглобинница».
Но обо всем по порядку..
Оксигенация, аноксия, гипоксия
Органы внешнего дыхания, – дыхательная мускулатура, воздухоносные пути и легкие со своей газообменной паренхимой, – в норме обеспечивают бесперебойное поступление в организм кислородсодержащей газовой смеси (которую мы привыкли называть воздухом) и обратный выброс в атмосферу отработанной ее порции, для дыхания уже не пригодной. Это, так сказать, механическая часть процесса оксигенации, – насыщения крови и, затем, тканей кислородом.
С конечными, самыми тонкими ответвлениями бронхиального древа, – бронхиолами, – сопряжены т.н. легочные ацинусы, каждый из которых представляет собой гроздь альвеолярных ходов и мешочков. В свою очередь, стенка каждого мешочка образована множеством единичных микроскопических пузырьков-альвеол, через мембраны которых, собственно, и осуществляется диффузия кислорода в кровь, одновременно с ее очищением от углекислого газа.
Кислород для нас, аэробов – основа существования, топливо; углекислота – выхлопной газ. Можно возразить: а как же, мол, пища и вода, а как же продукты их переработки?…
Одним воздухом, и то правда, сыт не будешь. Но до пищи и воды дело попросту не дойдет, если вы попробуете пообедать в вакууме. В печке может быть сколько угодно дров, но без огня вы не согреетесь. Огня же без кислорода, как известно, не бывает; горение – это не что иное, как окислительно-восстановительная реакция с интенсивным выделением тепла. Только что вы прошли краткий курс катаболизма аэробных живых организмов.
Рассмотрение конкретных окислительно-восстановительных каскадов (начиная со знаменитого цикла Кребса, или цикла трикарбоновых кислот), обеспечивающих жизнедеятельность организма энергией, здесь вполне можно опустить: слишком сложно, долго, да и не важно в данном контексте. Важно то, что кислород для человека, – как электричество для компьютера. Это энергоноситель, в отсутствие которого останавливается и отключается буквально всё, а в нашем случае еще и всё погибает, поскольку сохраняться на диске мы пока не научились.
Мы понимаем, что этот вариант является предельным, фатальным, – хотя и он, при всем трагизме, отнюдь не уникален в нашем несовершенном мире: летальность острой дыхательной недостаточности составляет примерно 35-40% (лет двести назад этот показатель практически не отличался от стопроцентного). Гораздо чаще встречается более или менее выраженная гипоксия, частичный «недобор» кислорода тканями. Человек об этом зачастую даже не знает, продолжая, например, курить табачные палочки. Но это до поры, до первой вынужденной остановки на лестнице, по которой раньше мы взлетали на пятый этаж бегом.
Если сложное электронное устройство работает «от розетки», а напряжение в сети по каким-то причинам постепенно снижается, то с определенного момента электроника не просто начинает сбоить, – она может самопроизвольно отключиться и/или серьезно повредиться.
Примерно то же самое происходит с тканями в условиях хронической гипоксии, нехватки кислорода, недостаточности тканевого дыхания. Но отчего возникает сама гипоксия?
Дыхательная функция крови
Тканевое (клеточное) дыхание обеспечивается не трахеей или, скажем, плеврой, а кровообращением. Это одна из функций крови, и без особой натяжки эту функцию можно назвать важнейшей, витальной, жизнеобеспечивающей.
Мы с детства знаем, что разрезанное яблоко на срезе темнеет и рыжеет, что кровь и планета Марс – красные, а беспризорный металл покрывается ржавчиной. Можем даже припомнить, что все перечисленные феномены объединены одним общим свойством: присутствием окислов железа, в частности, оксида Fe2O3 и/или его комбинаций с различными радикалами.
Кислород попадает к тканям в связанном виде, в составе сложных соединений с железом, из которых потом этакой птицей феникс восстанавливается внутри клеток, – дабы прореагировать с чем надо и исполнить свою энерговысвобождающую роль. Железо же входит в замысловатую формулу гемоглобина – сложного окрашенного белка (хромопротеина), состоящего как из органических, так и неорганических звеньев. Гемоглобин является транспортным веществом-челноком: он емко связывает не только кислород в альвеолах, но и углекислый газ на обратном пути.
Содержится гемоглобин в одном из трех форменных элементов крови – эритроцитах, дискообразных двояковогнутых красных кровяных тельцах. На самом деле они не настолько красные, как их рисуют, но тут все дело в количестве, расстоянии до наблюдателя и присутствии различных примесей. То, что содержит окислы железа в значительных концентрациях, издалека человеческому глазу действительно кажется красным или красновато-коричневым, ржавым.
Живая кровь – красная, а коагулированная на воздухе, – ржаво-темно-коричневая. В артериях, проложенных от легких к тканям, течет насыщенная кислородом ярко-алая кровь, а в отводящих венах – темная, деоксигенированная, нуждающаяся в очередном очищении и «заправке». Оттенок зависит от баланса между кислородом и углеродом в их сложных соединениях с гемоглобином.
Анемия
Теперь попробуем придумать логичные и правдоподобные причины кислородного голодания тканей – гипоксии. Вариантов на самом деле не так уж много.
Таким образом, анемия – группа заболеваний и синдромов, обусловленных кислородным голоданием тканей в связи с нарушениями дыхательной функции крови.
Виды анемии. Основные симптомы
Основных, принципиальных видов анемии тоже много не придумаешь, и, «вычислив» их логически, мы самостоятельно получим наиболее общую классификацию (для нужд реальной клинической практики разработаны и применяются другие, значительно более обширные, разветвленные и дифференцированные классификации, построенные на патогенетическом, этиологическом и других критериях). Итак:
В ряде источников в отдельную группу выделяются пернициозные анемии, обусловленные гиповитаминозом В, точнее, дефицитом фолиевой кислоты В9 и кобальтсодержащих витаминов В12.
Нередко рассматриваются отдельно анемии, обусловленные генетически, – например, серповидноклеточная анемия или талассемия, – а также псевдо- и истинные анемии беременных. Часть этих особых заболеваний и состояний уже освещена на сайте Лахта Клиники, другие будут непременно рассмотрены в будущем.
Вообще, многие специалисты резонно замечают, что считать анемию единой болезнью или даже группой болезней, – неправомерно. Анемия встречается и как симптом более общего заболевания, и как его патогенетическая почва, и как следствие одномоментного поражения, и как сравнительно самостоятельный наследственный синдром, и т.д. Слишком существенно, по мнению сторонников данной точки зрения, различаются этиопатогенез, клиника, подходы к терапии (к слову, по этой причине вопросы лечения в данном материале не затрагиваются).
Что касается симптоматики, попробуем из нашей «топливно-энергетической теории» вывести хотя бы наиболее общие следствия анемии. Ими должны быть (да и есть на самом деле) следующие симптомы:
Как указано выше, каждая анемия имеет свою клиническую специфику, и перечислить все многообразие симптомов совершенно немыслимо: в литературе описывается свыше 400 различных (различных!) вариантов анемии. В качестве заключения отметим лишь, что любая анемия, даже малосимптомная (не говоря уже о жизнеугрожающих) относится к самым глубинным, фундаментальным проблемам здоровья: она подрывает его энергетический базис, и, образно говоря, перекрывает организму кислород. Впрочем, почему «образно», – перекрывает вполне буквально. Поэтому, если какому бы то ни было врачу в рамках какого бы ни было обследования в вашем клиническом анализе крови «не понравятся» эритроциты, или гемоглобин, или гематокритное число, или цветовой показатель, или присутствие ретикулоцитов, – отнеситесь к этому серьезно и отреагируйте адекватно (в данном случае это означает выявить и устранить причину). Сделайте это, чтобы жить полнокровной жизнью.
Что такое дыхательная функция крови
Дыхательная функция крови
| Таблица 49. Состав вдыхаемого, альвеолярного и выдыхаемого воздуха (по Уайту и др., 1981) | ||||||
| Газ | Вдыхаемый воздух | Альвеолярный воздух | Выдыхаемый воздух | |||
| Р (гПа) | об.% | Р (гПа) | об.% | Р (гПа) | об.% | |
| О2 | 210,9 | 20,95 | 134,9 | 14,0 | 154,9 | 16,1 |
| CO2 | 4,0 | 0,04 | 53,3 | 5,6 | 38,0 | 4,5 |
| N2 | 795,3 | 79,0 | 762,4 | 80,0 | 757,7 | 79,2 |
| H2O | 6,67 | — | 62,7 | — | 62,7 | — |
| 1013,3 | 99,99 | 1013,3 | 99,6 | 1013,3 | 99,8 | |
Сущность дыхательной функции крови состоит в доставке кислорода от легких к тканям и углекислоты от тканей к легким (табл. 49).
Перенос кислорода кровью
Кровь осуществляет свою дыхательную функцию прежде всего благодаря наличию в ней гемоглобина. Физиологическая функция гемоглобина как переносчика кислорода основана на способности обратимо связывать кислород в зависимости от его напряжения в крови. Поэтому в легочных капиллярах происходит насыщение крови кислородом, а в тканевых капиллярах, где парциальное давление кислорода резко снижено, осуществляется отдача кислорода тканям.
В состоянии покоя ткани и органы человека потребляют около 200 мл кислорода в минуту. При тяжелой физической работе количество потребляемого тканями кислорода возрастает в 10 и более раз (до 2-3 л O2 в 1 мин). Доставка от легких к тканям такого количества кислорода в виде газа, физически растворенного в плазме, невозможна вследствие малой растворимости кислорода в воде и плазме крови (табл. 50).
Исходя из приведенных в табл. 50 данных, а также зная парциальное давление кислорода в артериальной крови 107-120 гПа (80-90 мм рт. ст.), нетрудно видеть, что количество физически раствoренного кислорода в плазме крови не может превышать 0,3 об%. При расчете кислородной емкости крови этой величиной можно пренебречь.
Гемоглобин человека содержит 0,335% железа. Каждый грамм-атом железа (55,84 г) в составе гемоглобина при полном насыщении кислoродом связывает 1 грамм-молекулу O2 (22 400 мл). Таким образом, 100 г гемоглобина может связать
0,335 x 22 400/ 55,84 = 134 мл 02,
15 х 1,34 х 96/ 100 = 19,3 мл О2 (в среднем 19-20 об. %).
В венозной крови при покое РO2 = 53,3 гПа, и в этих условиях гемоглобин насыщен кислородом лишь на 70-72%, т. е. содержание кислорода в 100 мл венозной крови не превышает
15 x 1,34 x 70 / 100 = 14,1 O2 (
Артериовенозная разница по кислороду будет равна

Таким образом, за 1 мин ткани в состоянии покоя получают 200-240 мл 02 (при условии, что минутный объем крови, протекающей через сердце, в покое составляет 4 л).
Возрастание интенсивности окислительных процессов в тканях, например при усиленной мышечной работе, всегда связано с более полным извлечением кислорода из крови. Кроме того, при физической работе резко увеличивается скорость кровотока. Зависимость между степенью насыщения гемоглобина кислородом и РO2 можно выразить в виде кривой насыщения гемоглобина кислородом, или кривой диссоциации оксигемоглобина, которая имеет S-образную форму и характеризует сродство гемоглобина к кислороду (рис. 129).
Характерная для гемоглобина S-образная кривая насыщения кислородом свидетельствует о том, что связывание первой молекулы кислорода одним из гемов гемоглобина облегчает связывание последующих молекул кислорода тремя другими оставшимися темами. Долгое время механизм, лежащий в основе этого эффекта, оставался загадкой, ибо, по данным рентгеноструктурного анализа, четыре гема в молекуле гемоглобина довольно далеко отстоят друг от друга и вряд ли могут оказывать взаимное влияние. В последнее время принято следующее объяснение происхождения S-образной кривой. Считают, что молекула гемоглобина способна обратимо распадаться на две половинки, каждая из которых содержит одну α-цепь и одну β-цепь:

При взаимодействии молекулы кислорода с одним из четырех гемов гемоглобина происходит присоединение кислорода (обозначим звездочкой) к одной из половинок молекулы гемоглобина (допустим, к α-цепи этой половинки). Как только такое присоединение произойдет, α-полипептидная цепь претерпевает конформационные изменения (условно обозначим эту форму О):

После этого конформационные изменения α-цепи механически передаются на тесно связанную с ней β-цепь, которая также подвергается конформационным сдвигам:

β-Цепь присоединяет кислород, имея уже большее сродство к нему. Таким путем связывание одной молекулы кислорода благоприятствует связыванию второй молекулы:

После насыщения кислородом одной половины молекулы гемоглобина возникает новое, внутреннее, напряженное состояние молекулы гемоглобина, которое вынуждает и вторую половинку гемоглобина изменить конформацию:

Теперь еще две молекулы кислорода, по-видимому, по очереди связываются со второй половинкой1 молекулы гемоглобина, образуя оксигемоглобин:

S-образная форма кривой насыщения гемоглобина кислородом имеет большое физиологическое значение. При такой форме кривой обеспечивается возможность насыщения крови кислородом при изменении РO2 в довольно широких предела. Например, дыхательная функция крови существенно не нарушается при снижении РO2 во вдыхаемом воздухе с 133,3 до 8-93,3 гПа.
Численно сродство гемоглобина к кислороду принято выражать через величину P50-парциальное напряжение О2, при котором 50% гемоглобина связано с кислородом (pH 7,4, температура 37°С). Нормальная величина Р50 около 34,67 гПа. Смещение кривой насыщения гемоглобина кислородом вправо означает уменьшение способности гемоглобина связывать кислород и, следовательно, сопровождается повышением Р50. Напротив, смещение кривой влево свидетельствует о повышенном сродстве гемоглобина к кислороду, величина Р50 снижена.
Ход кривой насыщения гемоглобина кислородом или диссоциации оксигемоглобина зависит от ряда факторов. Сродство гемоглобина к кислороду в первую очередь связано о pH. Чем ниже pH, тем меньше способность гемоглобина связывать кислород и тем больше Р50. В тканевых капиллярах pH ниже (поступает большое количество СO2), в связи с чем гемоглобин легко отдает кислород. В легких СO2 выделяется, pH повышается и гемоглобин жадно присоединяет кислород.
Способность гемоглобина связывать кислород зависит также от температуры. Чем выше температура (в тканях температура выше, чем в легких), тем меньше сродство гемоглобина к кислороду. Напротив, снижение температуры вызывает обратные явления.
Количество гемоглобина в крови, а также в какой-то мере способность связывать кислород (характер кривой диссоциации оксигемоглобина) несколько меняются с возрастом. Например, у новорожденных содержание гемоглобина доходит до 20-21% (вместо обычных для взрослого 13-16%). У человека имеется несколько гемоглобинов, которые образуются в различном количестве в разные стадии онтогенеза и отличаются по своему сродству к кислороду.
Рассмотрим нарушения дыхательной функции крови при некоторых патологических состояниях.
Различные формы гипоксии
Гипоксия вследствие понижения парциального давления кислорода во вдыхаемом воздухе.
Этот вид гипоксии возникает главным образом при подъеме на высоту. Может наблюдаться и в тех случаях, когда общее барометрическое давление нормально, но РO2 понижено, например при авариях в шахтах, неполадках в системе кислородообеспечения кабины летательного аппарата, в подводных лодках и т. п., а также во время операций при неисправности наркозно-дыхательной аппаратуры.
При экзогенной гипоксии развивается гипоксемия, т. е. уменьшается напряжение кислорода в артериальной крови и снижается насыщение гемоглобина кислородом.
Дыхательный (легочный) тип гипоксии возникает в связи с альвеолярной гиповентиляцией, что может быть обусловлено нарушением проходимости дыхательных путей (воспалительный процесс, инородные тела, спазм), уменьшением дыхательной поверхности легких (отек легкого, пневмония и т. д.). В подобных случаях снижаются РO2 в альвеолярном воздухе и напряжение кислорода в крови, в результате чего уменьшается насыщение гемоглобина кислородом. Обычно нарушается также выведение из организма углекислоты и к гипоксии присоединяется гиперкапния.
Сердечно-сосудистый (циркуляторный) тип гипоксии наблюдается при нарушениях кровообращения, приводящих к недостаточному кровоснабжению органов и тканей. Для газового состава крови в типичных случаях циркуляторной гипоксии характерны нормальные напряжение и содержание кислорода в артериальной крови, снижение этих показателей в венозной крови и высокая артериовенозная разница по кислороду.
Кровяной (гемический) тип гипоксии возникает в результате уменьшения кислородной емкости крови при анемиях, обусловленных значительным уменьшением эритроцитной массы или резким понижением содержания гемоглобина в эритроцитах. В этих случаях РO2 в венозной крови резко снижено.
Гемическая гипоксия наблюдается также при отравлений окисью углерода (образование карбоксигемоглобина) и метгемоглобинообразователями (метгемоглобинемия), а также пни некоторых генетически обусловленных аномалиях гемоглобина. При образовании карбоксигемоглобина и метгемоглобина напряжение кислорода в венозной крови и тканях оказывается значительно пониженным при одновременном уменьшении артериовенозной разницы содержания кислорода.
Перенос углекислоты кровью от тканей к легким
В организме человека, не выполняющего физической работы (состояние покоя), от тканей к легким каждую минуту переносится примерно 180 мл СO2.
Некоторое количество СO2 может переноситься в виде карбаминовой формы. Оказалось, что СO2 может присоединяться к гемоглобину посредством карбаминовой связи, образуя карбгемоглобин, или карбаминогемоглобин (впервые мысль о наличии углекислоты, непосредственно связанной с гемоглобином, была высказана И. М. Сеченовым):


В легочных капиллярах, опять таки в эритроцитах, происходит процесс вытеснения угольной кислоты из бикарбоната калия оксигемоглобином:
Образующаяся Н2СO3 быстро расщепляется при участии карбоангидразы на углекислый газ и воду. Низкое РO2 в просвете альвеол способствует диффузии углекислого газа из эритроцитов в легкие.
Итак, в форме бикарбоната при участии гемоглобина эритроцитов транспортируется с кровью к легким более 80% всего количества углекислоты.



