Что такое гиперполяризация мембраны
В нервных волокнах сигналы передаются с помощью потенциалов действия, которые представляют собой быстрые изменения мембранного потенциала, быстро распространяющиеся вдоль мембраны нервного волокна. Каждый потенциал действия начинается со стремительного сдвига потенциала покоя от нормального отрицательного значения до положительной величины, затем он почти так же быстро возвращается к отрицательному потенциалу. При проведении нервного сигнала потенциал действия движется вдоль нервного волокна вплоть до его окончания.
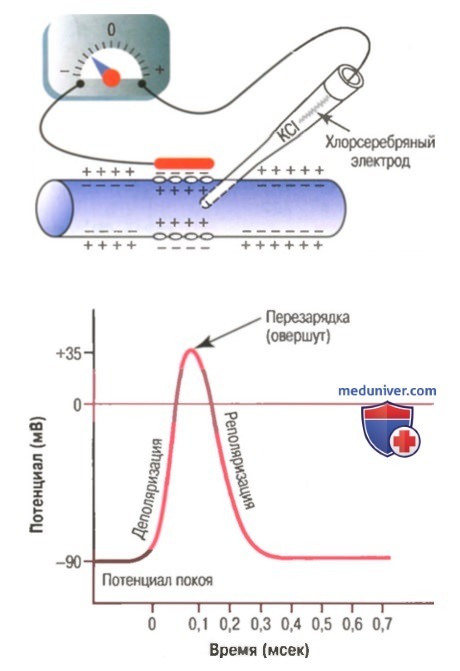 Типичный потенциал действия, зарегистрированный с помощью метода, представленного в верхней части рисунка.
Типичный потенциал действия, зарегистрированный с помощью метода, представленного в верхней части рисунка.
Фаза реполяризации. В течение нескольких долей миллисекунды после резкого повышения проницаемости мембраны для ионов натрия, натриевые каналы начинают закрываться, а калиевые — открываться. В результате быстрая диффузия ионов калия наружу восстанавливает нормальный отрицательный мембранный потенциал покоя. Этот процесс называют реполя-ризацией мембраны.
Для более полного понимания факторов, являющихся причиной деполяризации и реполяризации, необходимо изучить особенности двух других типов транспортных каналов в мембране нервного волокна: электроуправляемых натриевых и калиевых каналов.
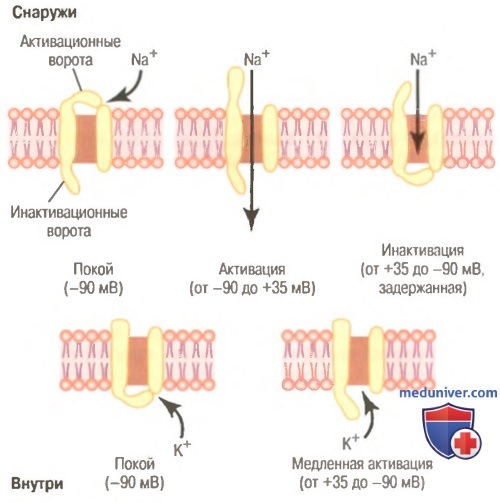 Типичные изменения состояния электроуправляемых натриевых (вверху) и калиевых (внизу) каналов при изменении мембранного потенциала от нормального отрицательного значения до положительных величин.
Типичные изменения состояния электроуправляемых натриевых (вверху) и калиевых (внизу) каналов при изменении мембранного потенциала от нормального отрицательного значения до положительных величин.
Показана последовательная активация и инактивация натриевых каналов и задержанная активация калиевых каналов.
Инактивация натриевого канала. В верхней правой части рисунке показано третье состояние натриевого канала. Увеличение потенциала, открывающее активационные ворота, закрывает инактивационные ворота. Однако инактивационные ворота закрываются в течение нескольких десятых долей миллисекунды после открытия активационных ворот. Это значит, что конформационное изменение, приводящее к закрытию инактивационных ворот, — процесс более медленный, чем конформационное изменение, открывающее активационные ворота. В результате через несколько десятых долей миллисекунды после открытия натриевого канала инактивационные ворота закрываются, и ионы натрия не могут более проникать внутрь волокна. С этого момента мембранный потенциал начинает возвращаться к уровню покоя, т.е. начинается процесс реполяризации.
Существует другая важная характеристикая процесса инактивации натриевого канала: инактивационные ворота не открываются повторно до тех пор, пока мембранный потенциал не вернется к значению, равному или близкому к уровню исходного потенциала покоя. В связи с этим повторное открытие натриевых каналов обычно невозможно без предварительной реполяризации нервного волокна.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в оглавление раздела «Физиология человека.»
Гиперполяризация ядерных спинов и её применение в биомедицине
Лекция профессора РАН, ассистент-профессора Университета Вандербильт (США) Эдуарда Юрьевича Чекменева посвящена гиперполяризации ядерных спинов — это явление позволяет в миллионы раз раз увеличить чувствительность МРТ. Молекулярное томографирование методом МРТ дает возможность визуализировать многие процессы метаболизма и функции органов, поскольку гиперполяризованные элементы и соединения можно наблюдать внутри живых организмов при миллимолярных концентрациях. Уже прошли первые клинические испытания по визуализации процессов гликолиза рака и функциональной томографии легких.
Фундаментальная основа ядерно-магнитного резонанса (ЯМР) и магнитно-резонансной томографии (МРТ) — энергетические переходы между энергетическими уровнями. Еще эти последние называют Зеемановскими уровнями, в честь их первооткрывателя Питера Зеемана, нидерладского физика и лауреата Нобелевской премии по физике 1902 года.
Далее, известно, что многие ядра атомов обладают магнитным спином. Если одновременно присутствует сильное постоянное магнитное поле, то происходит расщепление энергетических уровней в атоме.
Процесс гиперполяризации позволяет достичь (искусственно и временно) уровня поляризации ядерных спинов вплоть до теоретического уровня единицы или 100%, позволяя увеличить чувствительность МРТ в миллионы раз. Гиперполяризованные элементы и соединения успешно детектируются внутри живых организмов при миллимолярных концентрациях, позволяя молекулярное томографирование методом МРТ многих процессов метаболизма и функции органов. Например, не только прошли клинические испытания по визуализации процессов гликолиза рака и функциональной томографии легких в животных, но первые клинические испытания уже прошли и в волонтерах.
Помимо возможности получения уникальной информации по визуализации процессов на молекулярном уровне, МРТ сканирование с помощью гиперполяризованных соединений (также называемых контрастными агентами) может быть произведено за секунды — например, на одном вдохе пациента. Более того, чувствительность процесса сканирования не зависит от силы магнитного поля МРТ сканера, что дает потенциальную возможность использовать недорогие низкопольные МРТ сканеры, что сделает процедуру молекулярного МРТ существенно дешевле.
Вот каковы основные способы приготовления гиперполяризованных контрастных агентов:
Например, путем внутривенной инъекции в организм доставляется субстрат гиперполярированного [13С1]-пирувата. Это позволяет визуализировать раковые опухоли путем молекулярного обмена с лактатом, который значительно повышен в раковых опухолях по сравнению с нормальными тканями.
Во-вторых, спин-обменная оптическая накачка (СООН) или Spin Exchange Optical Pumping (SEOP). Здесь мощный (>100 Ватт) узкополосный (
Формирование мембранного потенциала покоя
Автор
Редакторы
Статья на конкурс «био/мол/текст»: Потенциал покоя — это важное явление в жизни всех клеток организма, и важно знать, как он формируется. Однако это сложный динамический процесс, трудный для восприятия целиком, особенно для студентов младших курсов (биологических, медицинских и психологических специальностей) и неподготовленных читателей. Впрочем, при рассмотрении по пунктам, вполне возможно понять его основные детали и этапы. В работе вводится понятие потенциала покоя и выделяются основные этапы его формирования с использованием образных метафор, помогающих понять и запомнить молекулярные механизмы формирования потенциала покоя.

«Био/мол/текст»-2011
Эта статья представлена на конкурс научно-популярных работ «био/мол/текст»-2011 в номинации «Лучшая обзорная статья».
Мембранные транспортные структуры — натрий-калиевые насосы — создают предпосылки для возникновения потенциала покоя. Предпосылки эти — разность в концентрации ионов на внутренней и наружной сторонах клеточной мембраны. Отдельно проявляет себя разность концентрации по натрию и разность концентрации по калию. Попытка ионов калия (K + ) выровнять свою концентрацию по обе стороны мембраны приводит к его утечке из клетки и потере вместе с ними положительных электрических зарядов, за счёт чего значительно усиливается общий отрицательный заряд внутренней поверхности клетки. Эта «калиевая» отрицательность составляет бóльшую часть потенциала покоя (−60 мВ в среднем), а меньшую его часть (−10 мВ) составляет «обменная» отрицательность, вызванная электрогенностью самого ионного насоса-обменника.
Давайте разбираться подробнее.
Зачем нам нужно знать, что такое потенциал покоя и как он возникает?
Вы знаете, что такое «животное электричество»? Откуда в организме берутся «биотоки»? Как живая клетка, находящаяся в водной среде, может превратиться в «электрическую батарейку» и почему она моментально не разряжается?
На эти вопросы можно ответить только в том случае, если узнать, как клетка создаёт себе разность электрических потенциалов (потенциал покоя) на мембране.
Совершенно очевидно, что для понимания того, как работает нервная система, необходимо вначале разобраться, как работает её отдельная нервная клетка — нейрон. Главное, что лежит в основе работы нейрона — это перемещение электрических зарядов через его мембрану и появление вследствие этого на мембране электрических потенциалов. Можно сказать, что нейрон, готовясь к своей нервной работе, вначале запасает энергию в электрической форме, а затем использует ее в процессе проведения и передачи нервного возбуждения.
Таким образом, наш самый первый шаг к изучению работы нервной системы — это понять, каким образом появляется электрический потенциал на мембране нервных клеток. Этим мы и займёмся, и назовём этот процесс формированием потенциала покоя.
Определение понятия «потенциал покоя»
В норме, когда нервная клетка находится в физиологическом покое и готова к работе, у неё уже произошло перераспределение электрических зарядов между внутренней и наружной сторонами мембраны. За счёт этого возникло электрическое поле, и на мембране появился электрический потенциал — мембранный потенциал покоя.
Как известно из физики, электрические заряды (движущиеся и неподвижные) формируют в пространстве электромагнитное поле, которое влияет на тела, обладающие электрическим зарядом. С точки зрения электромагнетизма клеточную мембрану можно представить как плоский конденсатор, заполненный однородным диэлектриком из неполярных молекул. Если конденсатор заряжен, то внутри него возникает электрическое поле, обусловленное поверхностной плотностью заряда. На поверхности мембраны возникают некомпенсированные заряды: положительные у «отрицательной» поверхности и отрицательные — у «положительной» [6].
Таким образом, мембрана оказывается поляризованной. Это означает, что она имеет разный электрический потенциал наружной и внутренней поверхностей. Разность между этими потенциалами вполне возможно зарегистрировать.
В этом можно убедиться, если ввести внутрь клетки микроэлектрод, соединённый с регистрирующей установкой. Как только электрод попадает внутрь клетки, он мгновенно приобретает некоторый постоянный электроотрицательный потенциал по отношению к электроду, расположенному в окружающей клетку жидкости. Величина внутриклеточного электрического потенциала у нервных клеток и волокон, например, гигантских нервных волокон кальмара, в покое составляет около −70 мВ. Эту величину называют мембранным потенциалом покоя (МПП). Во всех точках аксоплазмы этот потенциал практически одинаков.
Ноздрачёв А.Д. и др. Начала физиологии [5].
Ещё немного физики. Макроскопические физические тела, как правило, электрически нейтральны, т.е. в них в равных количествах содержатся как положительные, так и отрицательные заряды. Зарядить тело можно, создав в нем избыток заряженных частиц одного вида, например, трением о другое тело, в котором при этом образуется избыток зарядов противоположного вида. Учитывая наличие элементарного заряда (e), полный электрический заряд любого тела можно представить как q = ±N×e, где N — целое число.
Потенциал электростатического поля φ определяется как отношение потенциальной энергии W пробного заряда q к величине этого заряда: φ = W/q, откуда следует, что потенциал численно равен потенциальной энергии, которой обладает в данной точке поля единичный положительный заряд. Единицей измерения потенциала служит вольт (1 В) [4]. В некоторых случаях потенциал электрического поля нагляднее определяется как физическая величина, численно равная работе внешних сил против сил электрического поля E при перемещении единичного положительного точечного заряда из бесконечности в данную точку. Последнее определение удобно записать следующим образом:
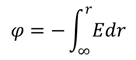
В электрофизиологии кроме потенциала покоя рассматриваются и другие электрические потенциалы: локальные постсинаптические и рецепторные потенциалы (возбуждающие и тормозные), электротонические и следовые потенциалы, миниатюрные потенциалы концевой пластинки, концентрационный потенциал и потенциал действия [5].
Потенциал покоя — это разность электрических потенциалов, имеющихся на внутренней и наружной сторонах мембраны, когда клетка находится в состоянии физиологического покоя. Его величина измеряется изнутри клетки, она отрицательна и составляет в среднем −70 мВ (милливольт), хотя в разных клетках может быть различной: от −35 мВ до −90 мВ.
Важно учитывать, что в нервной системе электрические заряды представлены не электронами, как в обычных металлических проводах, а ионами — химическими частицами, имеющими электрический заряд. И вообще в водных растворах в виде электрического тока перемещаются не электроны, а ионы. Поэтому все электрические токи в клетках и окружающей их среде — это ионные токи.
Итак, изнутри клетка в покое заряжена отрицательно, а снаружи — положительно. Это свойственно всем живым клеткам, за исключением, разве что, эритроцитов, которые, наоборот, заряжены отрицательно снаружи. Если говорить конкретнее, то получается, что снаружи вокруг клетки будут преобладать положительные ионы (катионы Na + и K + ), а внутри — отрицательные ионы (анионы органических кислот, не способные свободно перемещаться через мембрану, как Na + и K + ).
Теперь нам всего лишь осталось объяснить, каким же образом всё получилось именно так. Хотя, конечно, неприятно сознавать, что все наши клетки кроме эритроцитов только снаружи выглядят положительными, а внутри они — отрицательные.
Термин «отрицательность», который мы будем применять для характеристики электрического потенциала внутри клетки, пригодится нам для простоты объяснения изменений уровня потенциала покоя. В этом термине ценно то, что интуитивно понятно следующее: чем больше отрицательность внутри клетки — тем ниже в отрицательную сторону от нуля смещён потенциал, а чем меньше отрицательность — тем ближе отрицательный потенциал к нулю. Это намного проще понять, чем каждый раз разбираться в том, что же именно означает выражение «потенциал возрастает» — возрастание по абсолютному значению (или «по модулю») будет означать смещение потенциала покоя вниз от нуля, а просто «возрастание» — смещение потенциала вверх к нулю. Термин «отрицательность» не создаёт подобных проблем неоднозначности понимания.
Сущность формирования потенциала покоя
Попробуем разобраться, откуда берётся электрический заряд нервных клеток, хотя их никто не трёт, как это делают физики в своих опытах с электрическими зарядами.
Здесь исследователя и студента поджидает одна из логических ловушек: внутренняя отрицательность клетки возникает не из-за появления лишних отрицательных частиц (анионов), а, наоборот, из-за потери некоторого количества положительных частиц (катионов)!
Главный секрет появления отрицательности внутри клетки
Сразу откроем этот секрет и скажем, что клетка лишается части своих положительных частиц и заряжается отрицательно за счёт двух процессов:
Эти два процесса нам и надо объяснить.
Первый этап создания внутренней отрицательности: обмен Na + на K +
Но ведь при обмене одного положительного заряда (Na + ) на другой такой же положительный заряд (K + ) никакого дефицита положительных зарядов в клетке возникать не может! Правильно. Но, тем не менее, из-за этого обмена в клетке остаётся очень мало ионов натрия, потому что они почти все ушли наружу. И в то же время клетка переполняется ионами калия, которые в неё накачали молекулярные насосы. Если бы мы могли попробовать на вкус цитоплазму клетки, мы бы заметили, что в результате работы насосов-обменников она превратилась из солёной в горько-солёно-кислую, потому что солёный вкус хлорида натрия сменился сложным вкусом довольно-таки концентрированного раствора хлорида калия. В клетке концентрация калия достигает 0,4 моль/л. Растворы хлорида калия в пределах 0,009–0,02 моль/л имеют сладкий вкус, 0,03–0,04 — горький, 0,05–0,1 — горько-солёный, а начиная с 0,2 и выше — сложный вкус, состоящий из солёного, горького и кислого [8].
Важно здесь то, что обмен натрия на калий — неравный. За каждые отданные клеткой три иона натрия она получает всего два иона калия. Это приводит к потере одного положительного заряда при каждом акте ионного обмена. Так что уже на этом этапе за счёт неравноценного обмена клетка теряет больше «плюсов», чем получает взамен. В электрическом выражении это составляет примерно −10 мВ отрицательности внутри клетки. (Но помните, что нам надо ещё найти объяснение для оставшихся −60 мВ!)
Чтобы легче было запомнить работу насосов-обменников, образно можно выразиться так: «Клетка любит калий!» Поэтому клетка и затаскивает калий к себе, несмотря на то, что его и так в ней полно. И поэтому она невыгодно обменивает его на натрий, отдавая 3 иона натрия за 2 иона калия. И поэтому она тратит на этот обмен энергию АТФ. И как тратит! До 70% всех энергозатрат нейрона может уходить на работу натрий-калиевых насосов. (Вот что делает любовь, пусть она даже и не настоящая!)
Кстати, интересно, что клетка не рождается с готовым потенциалом покоя. Ей его ещё надо создать. Например, при дифференцировке и слиянии миобластов потенциал их мембраны изменяется от −10 до −70 мВ, т.е. их мембрана становится более отрицательной — поляризуется в процессе дифференцировки. А в экспериментах на мультипотентных мезенхимальных стромальных клетках костного мозга человека искусственная деполяризация, противодействующая потенциалу покоя и уменьшающая отрицательность клеток, даже ингибировала (угнетала) дифференцировку клеток [1].
Образно говоря, можно выразиться так: Создавая потенциал покоя, клетка «заряжается любовью». Это любовь к двум вещам:
Механизм насыщения клетки калием мы уже объяснили (это работа насосов-обменников), а механизм ухода калия из клетки объясним ниже, когда перейдём к описанию второго этапа создания внутриклеточной отрицательности. Итак, результат деятельности мембранных ионных насосов-обменников на первом этапе формирования потенциала покоя таков:
Можно сказать так: на первом этапе ионные насосы мембраны создают разность концентраций ионов, или градиент (перепад) концентрации, между внутриклеточной и внеклеточной средой.
Второй этап создания отрицательности: утечка ионов K + из клетки
Итак, что начинается в клетке после того, как с ионами поработают её мембранные натрий-калиевые насосы-обменники?
Из-за образовавшегося дефицита натрия внутри клетки этот ион при каждом удобном случае норовит устремиться внутрь: растворённые вещества всегда стремятся выровнять свою концентрацию во всём объёме раствора. Но это у натрия получается плохо, поскольку ионные натриевые каналы обычно закрыты и открываются только при определённых условиях: под воздействием специальных веществ (трансмиттеров) или при уменьшении отрицательности в клетке (деполяризации мембраны).
В то же время в клетке имеется избыток ионов калия по сравнению с наружной средой — потому что насосы мембраны насильно накачали его в клетку. И он, тоже стремясь уравнять свою концентрацию внутри и снаружи, норовит, напротив, выйти из клетки. И это у него получается!
Тут ещё важно понять то, что ионы натрия и калия как бы «не замечают» друг друга, они реагируют только «на самих себя». Т.е. натрий реагирует на концентрацию натрия же, но «не обращает внимания» на то, сколько вокруг калия. И наоборот, калий реагирует только на концентрацию калия и «не замечает» натрий. Получается, что для понимания поведения ионов надо по отдельности рассматривать концентрации ионов натрия и калия. Т.е. надо отдельно сравнить концентрацию по натрию внутри и снаружи клетки и отдельно — концентрацию по калию внутри и снаружи клетки, но не имеет смысла сравнивать натрий с калием, как это, бывает, делается в учебниках.
По закону выравнивания химических концентраций, который действует в растворах, натрий «хочет» снаружи войти в клетку; туда же его влечёт и электрическая сила (как мы помним, цитоплазма заряжена отрицательно). Хотеть-то он хочет, но не может, так как мембрана в обычном состоянии плохо его пропускает. Натриевые ионные каналы, имеющиеся в мембране, в норме закрыты. Если все же его заходит немножко, то клетка сразу же обменивает его на наружный калий с помощью своих натрий-калиевых насосов-обменников. Получается, что ионы натрия проходят через клетку как бы транзитом и не задерживаются в ней. Поэтому натрий в нейронах всегда в дефиците.
А вот калий как раз может легко выходить из клетки наружу! В клетке его полно, и она его удержать не может. Он выходит наружу через особые каналы в мембране — «калиевые каналы утечки», которые в норме открыты и выпускают калий [5, 7].
Зефиров А.Л. и Ситдикова Г.Ф. Ионные каналы возбудимой клетки (структура, функция, патология) [7].
От химического — к электрическому
А теперь — ещё раз самое главное. Мы должны осознанно перейти от движения химических частиц к движению электрических зарядов.
Калий (K + ) положительно заряжен, и поэтому он, когда выходит из клетки, выносит из неё не только самого себя, но и положительный заряд. За ним изнутри клетки к мембране тянутся «минусы» — отрицательные заряды. Но они не могут просочиться через мембрану — в отличие от ионов калия — т.к. для них нет подходящих ионных каналов, и мембрана их не пропускает. Помните про оставшиеся необъяснёнными нами −60 мВ отрицательности? Это и есть та самая часть мембранного потенциала покоя, которую создаёт утечка ионов калия из клетки! И это — большая часть потенциала покоя.
Для этой составной части потенциала покоя есть даже специальное название — концентрационный потенциал [5]. Концентрационный потенциал — это часть потенциала покоя, созданная дефицитом положительных зарядов внутри клетки, образовавшимся за счёт утечки из неё положительных ионов калия.
Ну, а теперь немного физики, химии и математики для любителей точности.
Электрические силы связаны с химическими по уравнению Гольдмана. Его частным случаем является более простое уравнение Нернста, по формуле которого можно рассчитать трансмембранную диффузионную разность потенциалов на основе различной концентрации ионов одного вида по разные стороны мембраны. Так, зная концентрацию ионов калия снаружи и внутри клетки, можно рассчитать калиевый равновесный потенциал EK:
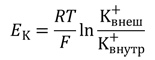
Более точно итоговая величина суммарного диффузионного потенциала, который создаётся утечкой нескольких видов ионов, рассчитывается по формуле Гольдмана-Ходжкина-Катца. В ней учтено, что потенциал покоя зависит от трех факторов: (1) полярности электрического заряда каждого иона; (2) проницаемости мембраны Р для каждого иона; (3) [концентраций соответствующих ионов] внутри (внутр) и снаружи мембраны (внеш). Для мембраны аксона кальмара в покое отношение проводимостей РK : PNa :PCl = 1 : 0,04 : 0,45 [5].
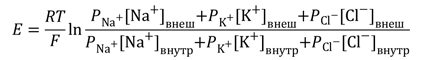
Заключение
Итак, поте нциал покоя состоит из двух частей:
Что интересно, калий перестанет выходить из клетки (точнее, его вход и выход уравниваются) только при уровне отрицательности клетки −90 мВ. В этом случае сравняются химические и электрические силы, проталкивающие калий через мембрану, но направляющие его в противоположные стороны. Но этому мешает постоянно подтекающий в клетку натрий, который несёт с собой положительные заряды и уменьшает отрицательность, за которую «борется» калий. И в итоге в клетке поддерживается равновесное состояние на уровне −70 мВ.
Вот теперь мембранный потенциал покоя окончательно сформирован.
Что такое гиперполяризация мембраны
Комплексная терапия при ишемических поражениях мозга, миокарда и других органов направлена на нормализацию кровотока и соответственно усиление оксигенации тканей.
Однако накопленные данные клинических и экспериментальных исследований указывают, что примерно в 30% подобных наблюдений развивается так называемый «синдром реперфузии», обусловленный неспособностью энергетической системы кардиомиоцитов утилизировать поступающий кислород и субстраты синтеза АТФ. При этом на начальных этапах постокклюзионной реперфузии возникают тяжелые осложнения, в частности, в ранее ишемизированном миокарде возможны нарушения ритма, включая фибрилляцию желудочков.
Установлено также развитие «синдрома реперфузии» в структурах ранее ишемизированного мозга в постокклюзионный период в виде отека мозга, гемморагий.
Несмотря на то, что гипоксия и гипероксия тканей являются диаметрально противоположными процессами, механизмы метаболических расстройств в ранний постокклюзионный период, по существу, во многом аналогичны таковым в условиях спастической ишемии тканей или гипоксии другого генеза и являются их логическим продолжением и усугублением.
В основе реперфузионных осложнений лежит избыточное поступление в условиях реканализации сосудов электролитов – кальция, натрия, а также воды, глюкозы, кислорода и других субстратов к альтерированным или некротизированным тканям, потерявшим способность их метаболизировать в типовых окислительно-восстановительных реакциях, а также в реакциях гликолиза, липолиза, протеолиза.
Повышение пассивной проницаемости цитоплазматических, митохондриальных, лизосомальных и других мембран, а также подавление активного энергозависимого транспорта электролитов, формирующихся в условиях гипоксии, и обеспечивают беспрепятственное проникновение в альтерированные клетки воды и растворимых в ней осмотически активных соединений, поступающих в ишемизированную ткань в условиях ее реперфузии.
При избыточном поступлении в клетку натрия, возникает смена частичной деполяризации клеток стойкой деполяризацией и вместо повышения возбудимости и функциональной активности, формируется резкое снижение возбудимости и соответственно функции клеток.
Избыточное накопление кальция в тканях в ранний постокклюзионный период приводит к дальнейшей активации мембранных фосфолипаз и дезинтеграции клеточных мембран, а активация под влиянием ионов кальция циклооксигеназы и липооксигеназы обеспечивает избыточное образование простагландинов и лейкотриенов, индуцирующих развитие перифокальной воспалительной реакции.
Развитие набухания митохондрий в период гипоксии, обусловленное чрезмерным поступлением в них кальция, калия, фосфатов и воды, приводит к нарушению утилизации субстратов в окислительно-восстановительных pеакциях, разобщению процессов окислительного фосфорилировання и дыхания, дефициту АТФ и ограничению всех энергозависимых реакций. В связи с этим усиление притока к ранее ишемизированным клеткам различных субстратов синтеза макроэргов не является фактором срочной восстановительной регенерации субклеточных фракций, в частности, митохондрий, так как проникающие с кровью субстраты не используются в должной мере или вообще не подвергаются метаболизму в аэробных реакциях. В то же время возможная активация анаэробного метаболизма усугубляет метаболический ацидоз и приводит к дальнейшей дестабилизации биологических мембран.
Развитие дезинтеграции, дезорганизации цитоплазматических, лизосомальных, митохондриальных и др. биологических мембран обеспечивается и активацией процессов липопероксидации, усугубляющейся в процессе реперфузии ишемизированных тканей.
Как указывалось выше, избыточное накопление кальция в ишемизированных структурах в период их реперфузии индуцирует активацию прокоагулянтных механизмов в окружающих, неповрежденных тканях, приводит к развитию тромбоза, эмболии сосудов с дальнейшим нарушением васкуляризации, трофики, оксигенации альтерированных структур.
Вышеизложенное делает очевидным большую значимость дестабилизации биологических мембран, нарушения электролитного баланса, энергообеспечения, коагуляционного гемостаза в механизмах развития гипоксического некробиоза и осложнений постокклюзионной реперфузии тканей.
14.2. Возможности медикаментозной коррекции метаболических расстройств при гипоксиях различного генеза (эффективность применения цитофлавина)
Гипоксия как типовой патологический процесс является основным патогенетическим фактором метаболических и функциональных расстройств, возникающих как при системных нарушениях гемодинамики, в частности, при сердечной недостаточности, различных видах шока, коллаптоидных состояниях, при ДВС – синдроме, так и при локальных расстройствах регионарного кровотока и микроциркуляции в зоне развития воспаления, тромбоза, эмболии артериальных сосудов, длительного спазма и т.д.
Одним из наиболее значимых механизмов повреждения клеток в условиях гипоксии и реперфузии ишемизированных тканей является активация процессов липопероксидации, на подавление которых должна быть направлена комплексная терапия заболеваний, связанных с развитием гипоксии.
В клинической практике постоянно предпринимаются попытки использования в комплексной терапии заболеваний, сопровождающихся развитием гипоксии, антигипоксантов и антиоксидантов.
Как известно, цитофлавин – комплексный препарат, разработанный научно-технологической фармацевтической фирмой «Полисан», включающий рибоксин, рибофлавин мононуклеотид, никотинамид, янтарную кислоту.
Останавливаясь на биологических эффектах отдельных компонентов цитофлавина, в частности, рибоксина, следует отметить, что указанное соединение является производным пурина, предшественником АТф, обладает способностью активировать ферменты цикла Кребса, стимулирует синтез нуклеотидов. В то же время рибоксин является агонистом пуринэргических рецепторов, которые широко представлены не только в ЦНС, но и в органах ЖКТ, миокарде, в эндотелии коронарных артерий и других сосудов.
Установлено, что пуринэргические рецепторы являются частью лиганд-контролируемых йоных каналов и оказывают метаболическое действие через ГТФ-связанные белки, что приводит к образованию дополнительного количества энергии за счет активации гликолиза.
Показано, что цитофлавин обеспечивает интенсификацию гексокиназной системы, а также фермента глюкозо-6-фосфат-дегидрогеназы и реакции карбоксилирования ПВК.
Таким образом, рибоксин, являясь одним из компонентов цитофлавина, обеспечивает ряд системных биологических эффектов, характеризующихся, в частности:
1. Индуцированной гиперполяризацией мембран клеток;
2. Неконкурентным антагонизмом НМДА к рецепторам;
3. Вазо- и коронародилятирующим действием;
4. Хроно-инотропным эффектом;
5. Седативным, анксиолитическим действиями;
6. Метаболотропным эффектами;
7. Моделированием поведенческих актов.
Другим компонентом цитофлавина является мононуклеотид рибофлавина, обеспечивающий сохранение и поддержание окислительно-восстановительных реакций, регулируемых флавиновыми коферментами. Среди последних особого внимания заслуживает глютатиоредуктаза, восстанавливающая пул глютатиона – важнейший компонент антиоксидантной системы клеток. Рибофлавин входит в состав дыхательных ферментов митохондрий (глютарил – КоА-дегидрогеназы, саркозиндегидрогеназы, электронпереносящих флавопротеинов, НАДН – дегидрогеназы и других), которые способны обеспечивать регенерацию НАД. Это свойство имеет отношение к ЦТК, так как сдвиг отношения НАДН/НАД в левую сторону является одним из важнейших факторов в ингибировании некоторых из его реакций. Отметим, что флавиновые ферменты участвуют и в реализации активности сукцинатдегидрогеназы и потреблении сукцината через метаболические шунты.
Биологическая активность цитофлавина в значительной мере определяются наличием в его составе никотинамида – амидного метаболита никотиновой кислоты – прекурсора коферментов дегидрогеназ(НАД и НАДФ). Соотношение НАДН/НАД является главным регуляторным механизмом ЦТК и отчасти – окислительного фосфорилирования.
Важным компонентом цитофлавина является янтарная кислота – эндогенный субстрат клетки. В условиях гипоксии ее действие реализуется в ЦТК и окислительном фосфорилировании.
В то же время введение экзогенного сукцината обеспечивает усиление фосфорилирования белков вследствие субстратной активации тропного фермента. Янтарная кислота усиливает потребление кислорода тканями и улучшает тканевое дыхание за счет активации транспорта электронов в митохондриях, воссоздания протоннового градиента на их мембранах и смещения кривой диссоциации оксигемоглобина вправо.
Как известно, интенсивность окисления сукцината зависит от его концентрации а клетке, а также от присутствия активаторов биотрансформации янтарной кислоты, т.е. от наличия предшествующих сукцинату и следующих после него биохимических субстратов. Это очень важное положение для практического применения цитофлавина в сочетании с препаратами других фармакологических групп. При низких и средних концентрациях сукцината восстанавливается пул НАД, при высоких – возникает сукцинатоксидазное окисление, возрастает антиоксидантная функция системы глютатиона.
В условиях гипоксии сукцинат (входящий в состав цитофлавина) может поглощаться через альтернативный метаболический путь сукцинатоксидазной системой с последующим потреблением янтарной кислоты в дыхательной цепи митохондрий.
Установлено участие сукцината в ресинтезе эндогенной ГАМК через α – кетоглютаровую кислоту и янтарный полуальдегид (в нервной ткани). ГАМК – ергические системы относятся к трофотропным (тормозным) системам ЦНС, противодействующим эрготропным. В совокупности с рибоксином и никотинамидом это свойство янтарной кислоты расширяет возможности применения цитофлавина в качестве неконкурентного антагониста НМДА – рецепторов и создает основу для его назначения в терапии не только острых состояний, но и хронических дегенеративно – дистрофических неврологических и сердечно – сосудистых заболеваний, астенических и абстинентных синдромов, в основе которых лежит эксайтотоксичность.
Вышеуказанные фармакологические эффекты цитофлавина, сочетающего в себе свойства антиоксиданта, антигипоксанта и мембранопротектора, обеспечили его широкое применение в последние годы в клинической практике, в частности, при лечении ишемического поражения миокарда и структур головного мозга.
Описана достаточно высокая эффективность использования цитофлавина при ишемическом инсульте, при тяжелых формах гипоксии в случае отравления нейротропными ядами, при хронических цереброваскулярных заболеваниях, при дисциркуляторной энцефалопатии, в постинсультном периоде ишемического нарушения мозгового кровообращения, при ишемическом поражении миокарда, а также при интраоперационной защите миокарда в коронарной хирургии на работающем сердце.



