Нервное волокно
Не́рвные воло́кна — отростки нейронов, покрытые глиальными оболочками.
В различных отделах нервной системы оболочки нервных волокон значительно отличаются по своему строению, что лежит в основе деления всех волокон на миелиновые и безмиелиновые. Те и другие состоят из отростка нервной клетки, лежащего в центре волокна, и поэтому называемого осевым цилиндром (аксоном), и окружающей его глиальной оболочки.
В зависимости от интенсивности функциональной нагрузки нейроны формируют тот или иной тип волокна. Для соматического отдела нервной системы, иннервирующей скелетную мускулатуру, обладающую высокой степенью функциональной нагрузки, характерен миелиновый тип нервных волокон, а для вегетативного отдела, иннервирующего внутренние органы — безмиелиновый тип.
Содержание
Классификация Эрлангера-Гассера
Является наиболее полной классификацией нервных волокон по скорости проведения нервного импульса.
| Тип волокна | Функция | Диаметр, мкм | Скорость проведения, м/с | Миелинизация |
|---|---|---|---|---|
| Aα | Афферентные — мышечные веретёна, сухожильные органы; эфферентные — скелетные мышцы | 10-20 | 60-120 | + |
| Aβ | Афферентные — тактильное чувство; коллатерали Aα волокон к интрафузальным мышечным волокнам | 7-15 | 40-90 | + |
| Aγ | Эфферентные — мышечные веретёна | 4-8 | 15-30 | + |
| Aδ | Афферентные — температура, быстрое проведение боли | 3-5 | 5-25 | + |
| B | Симпатические, преганглионарные; постганглионарные волокна цилиарного ганглия | 1-3 | 3-15 | прерывистая |
| C | Симпатические, постганглионарные; афферентные — медленное проведение боли | 0,3-1 | 0,5-2 | — |
Классификация по Ллойду
Классифицирует только афферентные нейроны.
| Тип волокна | Функция | Диаметр, мкм | Скорость проведения, м/с | Миелинизация |
|---|---|---|---|---|
| Ia | Мышечные веретёна | 18-22 | 90-120 | + |
| Ib | Сухожильные рецепторы | 15-18 | 60-90 | + |
| II | Механорецепторы кожи, вторичные мышечные веретёна | 7-15 | 40-90 | + |
| III | Рецепторы связок | 1-5 | 3-25 | прерывистая |
| IV | Болевые рецепторы, рецепторы соединительной ткани | 0,1-1 | 0,5-2 | — |
Миелинизация нервных волокон
При формировании безмиелинового нервного волокна осевой цилиндр (отросток нейрона) погружается в тяж из леммоцитов, цитолеммы которых прогибаются и плотно охватывают осевой цилиндр в виде муфты, края которой смыкаются над ним, образуя дупликатуру клеточной мембраны — мезаксон. Соседние леммоциты входящие в состав сплошного глиального тяжа своими цитолеммами образуют простые контакты. Безмиелиновые нервные волокна имеют слабую изоляцию, допускающую переход нервного импульса с одного волокна на другое, как в области мезаксона, так и в области межлеммоцитарных контактов.
Миелиновые нервные волокна значительно толще безмиелиновых. Принцип образования их оболочек такой же, как и безмиелиновых, то есть осевые цилиндры также прогибают цитолемму глиоцитов, образуя линейный мезаксон. Однако, быстрый рост нейронов соматического отдела нервной системы, связанный с формированием и ростом всего организма, приводит к вытягиванию мезаксонов, многократному обращению леммоцитов вокруг осевых цилиндров. В результате образуются концентрические наслоения. При этом цитоплазма с ядром леммоцитов оттесняется в область последнего витка, образующего наружный слой оболочек волокна, называемой шванновской оболочкой или неврилеммой. Внутренний слой, состоящий из витков мезаксона, называется миелиновым или миелиновой оболочкой. Следствием того, что миелинизация происходит в процессе роста как отростков нейронов, так и самих леммоцитов, является постепенное увеличение количества витков и размеров мезаксона, то есть каждый последующий виток шире предыдущего. Следовательно, последний виток, содержащий цитоплазму с ядром леммоцита является самым широким. Толщина миелина по длине волокна неоднородна, а в местах контактов соседних леммоцитов слоистая структура исчезает и контактируют лишь наружные слои, содержащие цитоплазму и ядро. Места их контактов называются узловыми перехватами (перехватами Ранвье), возникающими вследствие отсутствия здесь миелина и истончения волокна.
В ЦНС миелинизация нервного волокна происходит за счет обхвата осевых цилиндров отростками олигодендроцитов.
Как мембранная структура миелин имеет липидную основу и при обработке окисями тяжёлых металлов окрашивается в тёмный цвет. Другие компоненты мембраны и промежутки не окрашиваются, поэтому периодически встречаются светлые полоски − насечки миелина (насечки Шмидта-Лантермана), которые соответствуют небольшим прослойкам цитоплазмы леммоцита.
В цитоплазме осевого цилиндра располагаются продольно ориентированные нейрофибриллы и митохондрии, которых больше в непосредственной близости к перехватам и в концевых аппаратах волокна. Цитолемма осевого цилиндра (аксона) называется аксолеммой. Она обеспечивает проведение нервного импульса, который представляет собой волну деполяризации аксолеммы. Если осевой цилиндр представлен нейритом, то в нём отсутствуют гранулы базофильного вещества.
См. также
Сома · Аксон (Аксонный холмик, Терминаль аксона, Аксоплазма, Аксолемма, Нейрофиламенты)
Нервная ткань
Нейрон
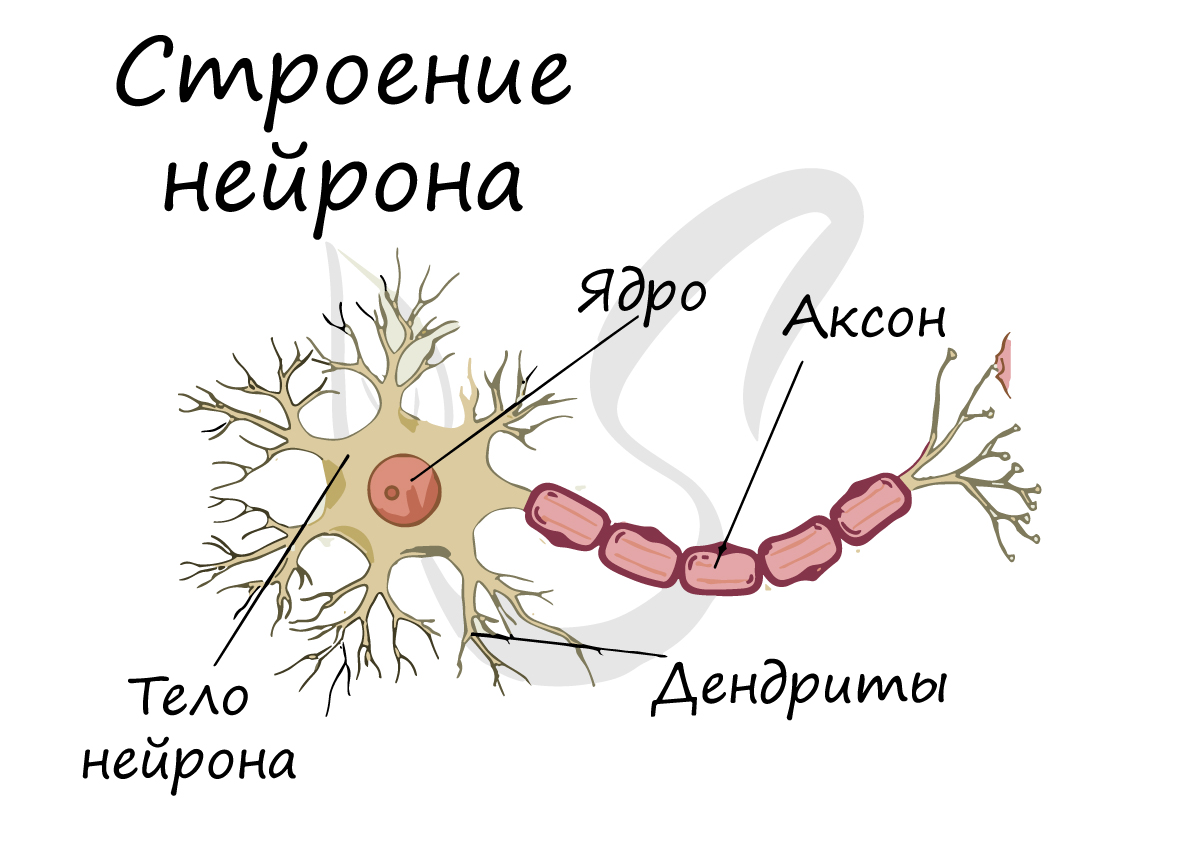
Нейроны обладают 4 свойствами:
Отростки нейронов проводят нервные импульсы и передают их другим нейронам, эффекторам, благодаря чему мышцы сокращаются или расслабляются, а секреция желез усиливается или уменьшается.
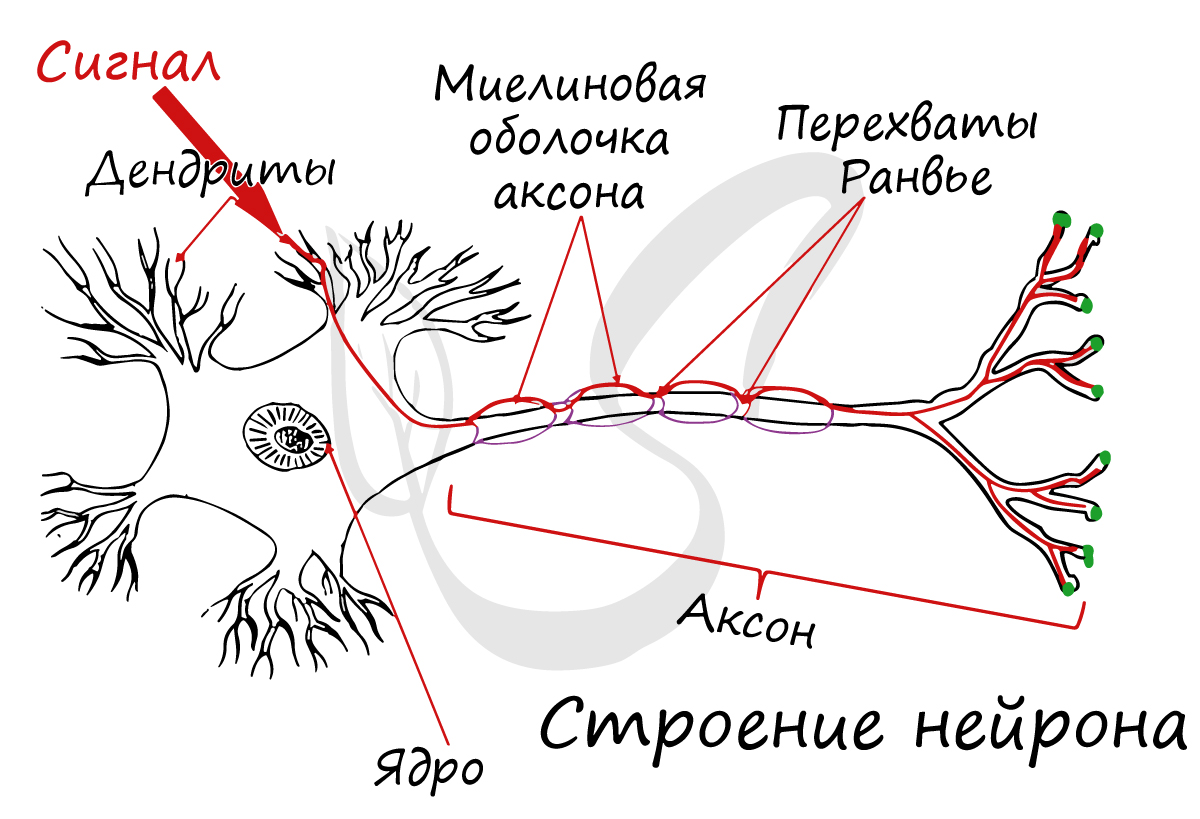
Миелиновая оболочка
В миелиновых нервных волокнах отростки нейронов покрыты миелиновой оболочкой (на 70-75% состоит из липидов (жиров)), которая обеспечивает изолированное проведение нервного импульса по нерву. Если бы не было миелиновой оболочки (вообразите!) нервные импульсы распространялись бы хаотично, и, когда мы хотели сделать движение рукой, то вместе с рукой двигалась бы нога.

Миелиновый слой представлен несколькими слоями мембраны глиальной клетки (леммоцит, шванновская клетка), которые закручиваются вокруг осевого цилиндра (отростка нейрона). Это закручивание хорошо видно на картинке, где изображен здоровый нерв, чуть выше 😉

Нейроглия (греч. νεῦρον — волокно, нерв + γλία — клей)
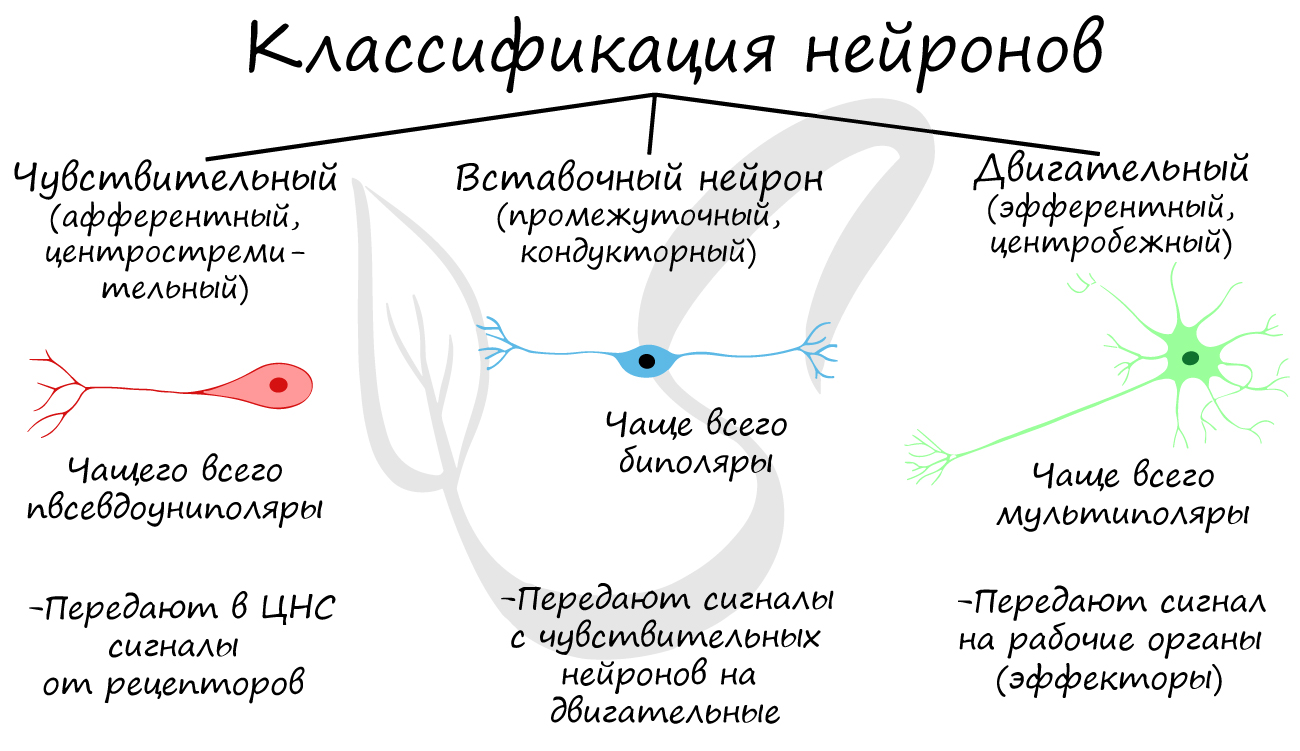
Классификация нейронов
Нейроны функционально подразделяются на чувствительные, двигательные и вставочные.
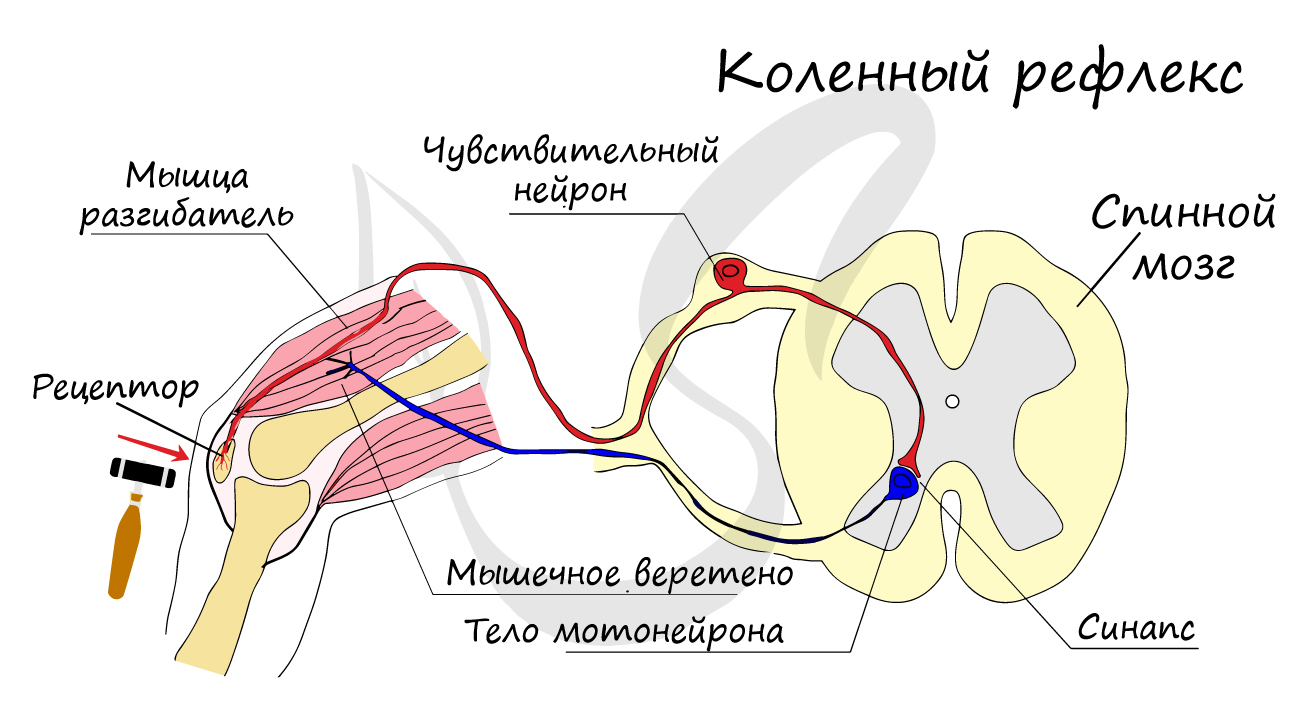
Синапс
Разберем строение синапса на схеме. Его составляют пресинаптическая мембрана аксона, рядом с которой расположены везикулы (лат. vesicula — пузырек) с нейромедиатором внутри (ацетилхолином). Если нервный импульс достигает терминали (окончания) аксона, то везикулы начинают сливаться с пресинаптической мембраной: ацетилхолин поступает наружу, в синаптическую щель.
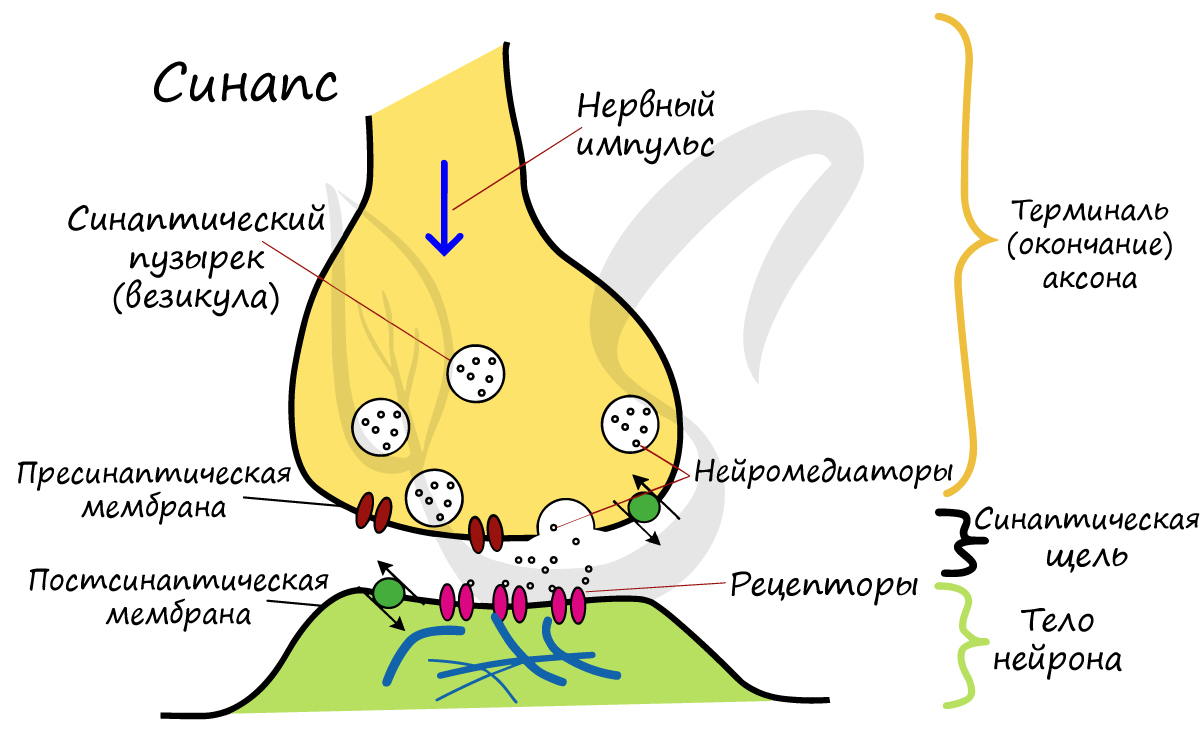
Попав в синаптическую щель, ацетилхолин связывается с рецепторами на постсинаптической мембране, таким образом, возбуждение (нервный импульс) передается другому нейрону. Так устроена нервная система: электрический путь передачи сменяется химическим (в синапсе).
Яд кураре
Гораздо интереснее изучать любой предмет на примерах, поэтому я постараюсь как можно чаще радовать вас ими 😉 Не могу утаить историю о яде кураре, который используют индейцы для охоты с древних времен.

Нервы и нервные узлы
Болезни нервной системы
Неврологические болезни могут развиваться в любой точке нервной системы: от этого будет зависеть клиническая картина. В случае повреждения чувствительного пути пациент перестает чувствовать боль, холод, тепло и другие раздражители в зоне иннервации пораженного нерва, при этом движения сохранены в полном объеме.
Если повреждено двигательное звено, движение в пораженной конечности будет невозможно: возникает паралич, но чувствительность может сохраняться.

© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Что такое нервные волокна биология
В теле человека бессчетное количество клеток, каждая из которых имеет собственную функцию. Среди них самые загадочные – нейроны, отвечающие за любое совершаемое нами действие. Попробуем разобраться как работают нейроны и в чем их предназначение.
Что такое нейрон (нейронные связи)
Нейроны работают при помощи электрических сигналов и способствуют обработке мозгом поступающей информации для дальнейшей координации производимых телом действий.
Эти клетки являются составляющей частью нервной системы человека, предназначение которой состоит в том, чтобы собрать все сигналы, поступающие из вне или от собственного организма и принять решение о необходимости того или иного действия. Именно нейроны помогают справиться с такой задачей.
Каждый из нейронов имеет связь с огромным количеством таких же клеток, создаётся своеобразная «паутина», которая называется нейронной сетью. Посредством данной связи в организме передаются электрические и химические импульсы, приводящие всю нервную систему в состояние покоя либо, наоборот, возбуждения.
К примеру, человек столкнулся с неким значимым событием. Возникает электрохимический толчок (импульс) нейронов, приводящий к возбуждению неровной системы. У человека начинает чаще биться сердце, потеют руки или возникают другие физиологические реакции.
Мы рождаемся с заданным количеством нейронов, но связи между ними еще не сформированы. Нейронная сеть строится постепенно в результате поступающих из вне импульсов. Новые толчки формируют новые нейронные пути, именно по ним в течение жизни побежит аналогичная информация. Мозг воспринимает индивидуальный опыт каждого человека и реагирует на него. К примеру, ребенок, схватился за горячий утюг и отдернул руку. Так у него появилась новая нейронная связь.
Стабильная нейронная сеть выстраивается у ребенка уже к двум годам. Удивительно, но уже с этого возраста те клетки, которые не используются, начинают ослабевать. Но это никак не мешает развитию интеллекта. Наоборот, ребенок познает мир через уже устоявшиеся нейронные связи, а не анализирует бесцельно все вокруг.
Познание нового опыта на протяжении всей жизни приводит к отмиранию ненужных нейронных связей и формированию новых и полезных. Этот процесс оптимизирует головной мозг наиболее эффективным для нас образом. Например, люди, проживающие в жарких странах, учатся жить в определенном климате, а северянам нужен совсем другой опыт для выживания.
Сколько нейронов в мозге
Нервные клетки в составе головного мозга занимают порядка 10 процентов, остальные 90 процентов это астроциты и глиальные клетки, но их задача заключается лишь в обслуживании нейронов.
Подсчитать «вручную» численность клеток в головном мозге также сложно, как узнать количество звезд на небе.
Тем не менее ученые придумали сразу несколько способов для определения количества нейронов у человека:
Строение нейрона
На рисунке приведено строение нейрона. Он состоит из основного тела и ядра. От клеточного тела идет ответвление многочисленных волокон, которые именуются дендритами.

Мощные и длинные дендриты называются аксонами, которые в действительности намного длиннее, чем на картинке. Их протяженность варьируется от нескольких миллиметров до более метра.
Аксоны играют ведущую роль в передаче информации между нейронами и обеспечивают работу всей нервной системы.
Место соединения дендрита (аксона) с другим нейроном называется синапсом. Дендриты при наличии раздражителей могут разрастись настолько сильно, что станут улавливать импульсы от других клеток, что приводит к образованию новых синаптических связей.
Синаптические связи играют существенную роль в формировании личности человека. Так, личность с устоявшимся позитивным опытом будет смотреть на жизнь с любовью и надеждой, человек, у которого нейронные связи с негативным зарядом, станет со временем пессимистом.
Виды нейронов и нейронных связей
Нейроны можно обнаружить в различных органах человека, а не исключительно в головном мозге. Большое их количество расположено в рецепторах (глаза, уши, язык, пальцы рук – органы чувств). Совокупность нервных клеток, которые пронизывают наш организм составляет основу периферической нервной системы. Выделим основные виды нейронов.
Слаженная работа нейронов трех типов выглядит так: человек «слышит» запах шашлыка, нейрон передает информацию в соответствующий раздел мозга, мозг передает сигнал желудку, который выделяет желудочный сок, человек принимает решение «хочу есть» и бежит покупать шашлык. Упрощенно так это действует.
Самыми загадочными являются промежуточные нейроны. С одной стороны, их работа обуславливает наличие рефлекса: дотронулся до электричества – отдернул руку, полетела пыль –зажмурился. Однако, пока не объяснимо как обмен между волокнами рождает идеи, образы, мысли?
Единственное, что установили ученые, это тот факт, что любой вид мыслительной деятельности (чтение книг, рисование, решение математических задач) сопровождается особой активностью (вспышкой) нервных клеток определенного участка головного мозга.
Есть особая разновидность нейронов, которые именуются зеркальными. Их особенность заключается в том, что они не только приходят в возбуждение от внешних сигналов, но и начинают «шевелиться», наблюдая за действиями своих собратьев – других нейронов.
Функции нейронов
Без нейронов невозможна работа организма человека. Мы увидели, что эти наноклетки отвечают буквально за каждое наше движение, любой поступок. Выполняемые ими функции до настоящего времени в полной мере не изучены и не определены.
Существует несколько классификаций функций нейронов. Мы остановимся на общепринятой в научном мире.
Функция распространения информации
Данная функция:
Суть ее в том, что нейронами обрабатываются и переносятся в головной мозг все импульсы, которые поступают из окружающего мира или собственного тела. Далее происходит их обработка, подобно тому, как работает поисковик в браузере.
По результатам сканирования сведений из вне, головной мозг в форме обратной связи передает обработанную информацию к органам чувств или мышцам.
Мы не подозреваем, что в нашем теле происходит ежесекундная доставка и переработка информации, не только в голове и на уровне периферической нервной системы.
До настоящего времени создать искусственный интеллект, который бы приблизился к работе нейронных сетей человека, не удалось. У каждого из 85 миллиардов нейронов имеется, как минимум, 10 тысяч обусловленных опытом связей, и все они работают на передачу и обработку информации.
Функция аккумуляции знаний (сохранения опыта)
Человек обладает памятью, возможностью понимать суть вещей, явлений и действий, которые он единожды или многократно повторял. За формирование памяти отвечают именно нейронные клетки, точнее нейротрансмиттеры, связующие звенья между соседними нейронами.
Таким образом, за память отвечает не какая-то отдельная часть мозга, а маленькие белковые мостики между клетками. Человек может потерять память, когда произошло крушение этих нервных связей.
Функция интеграции
Данная функция позволяет взаимодействовать между собой отдельным долям головного мозга. Как мы уже сказали, сигналы от разных органов чувств поступают в разные отделы мозга.
Нейроны посредством «вспышек» активности передают и принимают импульсы в разных частях мозга. Так происходит процесс появления мыслей, эмоций и чувств. Чем больше таких разноплановых связей, тем эффективнее человек мыслит. Если человек способен к размышлениям и аналитике в определенном направлении, то он будет хорошо соображать и в другом вопросе.
Функция производства белков
Нейроны – настолько полезные клетки, что не ограничиваются только передаточными функциями. Нервные клетки вырабатывают необходимые для жизни человека белки. Опять же ключевую роль в производстве белков имеют нейротрансмиттеры, которые отвечают за память.
Всего в невронах индуцируется порядка 80 белков, вот основные из них, влияющие на самочувствие человека:
Прекращение выработки белков или их выпуск в недостаточном количестве способны привести к тяжелым заболеваниям.
Восстанавливаются ли нервные клетки
При нормальном состоянии организма нейроны могут жить и функционировать очень долго. К сожалению, случается так, что они начинают массово погибать. Причин разрушения нервных волокон может быть много, но до конца механизм их деструкции не изучен.
Установлено, что нервные клетки погибают из-за гипоксии (кислородное голодание). Нейронные сети рушатся при отдельных травмах головного мозга, человек теряет память или утрачивает способность к хранению информации. В этом случае сами нейроны сохранены, но теряется их передаточная функция.
Отсутствие допамина ведет к развитию болезни Паркинсона, а его переизбыток является причиной шизофрении. Почему прекращается выработка белка не известно, спусковой механизм не выявлен.
Гибель нервных клеток происходит при алкоголизации личности. Алкоголик со временем может совершенно деградировать и утратить вкус к жизни.
Формирование нервных клеток происходит при рождении. Долгое время ученые полагали, что со временем нейроны отмирают. Поэтому с возрастом человек утрачивает способность накапливать информацию, хуже соображает. Нарушение функции по выработке допамина и серотонина связывается с наличием практически у всех пожилых людей депрессивных состояний.
Гибель нейронов, действительно неизбежна, в год исчезает примерно 1 процент от их количества. Но есть и хорошие новости. Последние исследования показали, что в коре головного мозга есть особенный участок, именуемый гипокаммом. Именно в нем генерируются новые чистые нейроны. Подсчитано примерное количество генерируемых ежедневно нервных клеток – 1400.
В науке обозначилось новое понятие «нейропластичность», обозначающее возможность мозга регенерироваться и перестраиваться. Но есть одна тонкость: новые нейроны еще не имеют никакого опыта и наработанных связей. Поэтому с возрастом или после заболевания мозг нужно тренировать, как и все иные мышцы тела: получать новые знания, анализировать происходящие события и явления.
Подобно тому, как мы усиливаем бицепс при помощи гантели, активизировать процесс включения новых нервных клеток можно следующими способами:
Механизм возрождения прост. У нас имеются совершенно не задействованные новые клетки, которые нужно заставить работать, а сделать это можно лишь путем постановки новых задач и изучения неизвестных предметных сфер.
НЕРВНЫЕ ВОЛОКНА
Нервные волокна [neurofibrae (LNH)] — отростки нервных клеток (нейроцитов), окруженные глиальной оболочкой, обеспечивающие проведение возбуждения между нервными клетками и структурными компонентами других тканей. Отростки нейроцита в составе Нервного волокна называются осевыми цилиндрами, а глиальные клетки, формирующие оболочку волокна, или глиальную оболочку,— нейролеммоцитами (шванновскими клетками). Различают два вида Нервных волокон: безмякотные, или безмиелиновые (neurofibrae nonmyelinatae, LNH), и мякотные, или миелиновые (neurofibrae myelinatae, LNH). Критерием подобного разделения Нервных волокон явилась открытая Шультце (М. J. S. Schultze, 1871) способность оболочки некоторых волокон окрашиваться в черный цвет в результате импрегнации солями осмия. Волокна с четко выраженной оболочкой, импрегнирующейся солями осмия, были названы миелиновыми: волокна, не имеющие такой оболочки,— безмиелиновыми. В миелиновых волокнах нейролеммоциты формируют вокруг осевого цилиндра пространственно сложную мембранную структуру, в безмиелиновых — относительно простую (рис. 1).

Безмиелиновые нервные волокна были впервые описаны Р. Ремаком в 1838 г. Диаметр этих Н. в. 1—4 мкм. Осевые цилиндры безмиелиновых волокон окружены глиальной оболочкой, сформированной цепью нейролеммоцитов. В световом микроскопе нейролеммоциты безмиелиновых волокон видны в виде тонких удлиненных ядер. Электронно-микроскопически (исследования Г. Гассера и др.) показано, что каждый осевой цилиндр вдавливает поверхность нейролеммоцита и располагается в отдельном углублении. Между плазматической мембраной нейролеммоцитов и цитолеммой осевого цилиндра находится промежуток в 12 — 15 нм. По данным Диксона (A. Dixon, 1963), один нейролеммоцит может служить оболочкой для 5—21 осевых цилиндров. По данным Г. Гассера (1958), каждый осевой цилиндр имеет глиальную оболочку разной протяженности. В некоторых случаях осевой цилиндр окутывается нейролеммоцитом частично, складки его остаются несомкнутыми. Если складки нейролеммоцитов смыкаются, их цитоплазматические мембраны располагаются параллельно друг другу. Пара примыкающих друг к другу мембран ней-ролеммоцитов называется мезаксоном. Длина и форма мезаксона различна: он может быть коротким и прямым, удлиненным и несколько изогнутым вокруг осевого цилиндра либо окружать его неполной спиралью, между витками к-рой располагается цитоплазма нейролеммоцита.
Между осевыми цилиндрами и нейролеммоцитами возможны и иные взаимоотношения: в дупликатуре цитолеммы нейролеммоцитов иногда заключено несколько осевых цилиндров. Глиальная оболочка формируется цепочкой клеток, располагающихся вдоль осевых цилиндров. Там, где нейролеммоциты контактируют между собой, цитоплазматические выросты их переплетаются и могут перекрывать друг друга. Ядра нейролеммоцитов обычно располагаются в средней части клеток; здесь же имеется скопление цитоплазмы, где находятся органеллы — гранулярный Эндоплазматический ретикулум, пластинчатый комплекс (комплекс Гольджи), митохондрии, нейрофибриллы (микротрубочки, микрофиламенты), лизосомы. Органеллы, помимо перинуклеарной области, обнаруживаются по всей цитоплазме, но, как правило, большая часть их локализуется у цитолеммы. Осевые цилиндры безмиелиновых Н. в. имеют строение, типичное для соответствующих отростков нервной клетки (см.) — дендритов и аксонов. В ряде случаев, напр, в обонятельном нерве, тонкие осевые цилиндры могут быть собраны в отдельные пучки, каждый из к-рых имеет собственный мезаксон, причем мезаксоны соседних пучков могут тесно контактировать друг с другом.


По периферии глиальной оболочки располагается тонкая базальная мембрана (см.) толщиной 5 нм; она имеется только на наружной поверхности цитолеммы нейролеммоцитов и не проникает в щели между смежными клетками. Базальная мембрана может находиться в непосредственном контакте с осевыми цилиндрами; в этом случае осевые цилиндры не целиком погружены в нейролеммоцит, и мезаксон отсутствует. За базальной мембраной следует соединительная ткань эндоневрии (см. Нервы).
Миелиновые нервные волокна построены как и безмиелиновых: в их состав входят осевой цилиндр и глиальная оболочка. При изучении миелинового Н. в. с помощью светового микроскопа после обработки осмиевой к-той вокруг осевого цилиндра обнаруживается интенсивно окрашенная оболочка — мякотная, или миелиновая (рис. 2). Толщина ее достигает 8— 20 мкм. При обработке волокна веществом, растворяющим липиды, а затем осмиевой к-той, миелиновая оболочка не выявляется, что указывает на преобладание в ее составе липидов (22% сухого веса миелина составляет белок, 78% — липиды, из к-рых 42% фосфолипидов, 28% цереброзидов, 25% холестерина, остальное сульфатиды). Светооптически показано, что по всей длине миелиновых Н. в. на равном расстоянии друг от друга встречаются участки, лишенные миелина. Эти участки названы перехватами узла — перехваты Ранвье (рис. 2, 3). Участки миелина, находящиеся между соседними перехватами, получили название межузловых сегментов. Количество межузловых сегментов, возникающих в процессе развития, во время миелогенеза, остается постоянным на протяжении всей жизни. После того как миелинизации заканчивается, Н. в. продолжает расти в длину, а количество межузловых сегментов остается постоянным. Поэтому, чем длиннее становится Н. в. в процессе роста, тем длиннее и межузловые сегменты. Первыми миелинизируются Н. в., к-рые у взрослых особей имеют больший диаметр. У этих Н. в. межузловые сегменты длиннее, чем у тонких волокон, глиальные оболочки к-рых возникают на более поздних этапах развитии.
Первые электронно-микроскопические описании мякотных Нервных волокон сделаны в 1950 г. Шёстрандом (F. S. Sjdstrancl) и Фернандес-Мораном (H. Fernandez-Moran). Показано, что миелиновая оболочка представляет собой систему концентрических слоев. Каждый слой состоит из главных оптически плотных линий толщиной 2,5—3 нм, отделиющихси друг от друга светлыми промежутками шириной 9 нм (в совокупности это составляет так наз. период с величиной 12 нм). В каждом светлом промежутке существует еще одна плотная линия — промежуточная, к-рая тоньше главной. В процессе эмбрионального развитии при образовании миелинового Н. в. отросток нейроцита изолируется нейролеммоцитами и располагается в бороздке, проходящей вдоль поверхности удлиненного нейролеммоцита. В последующем бороздка углубляется, ее края сближаются и формируется мезаксон. В процессе миелогенеза мезаксон значительно удлиняется и спирально закручивается вокруг осевого цилиндра (рис. 3,6). В результате этого, а также по мере роста Н. в. цитоплазма вместе с мембранами нейролеммоцитов спиралеобразно окружает осевой цилиндр. Вначале спиралеобразные пластины возникающей оболочки отделены друг от друга цитоплазмой глии. При последующем наслаивании пластин цитоплазма вытесняется, так что по окончании миелинизации она остается лишь на внутренней и наружной поверхностях миелиновой оболочки (так наз. периферическая цитоплазма). В зрелых Н. в. миелиновая оболочка остается связанной как с поверхностью нейролеммоцита, так и с плазмалеммой осевого цилиндра — наружный и внутренний мезаксон.
Миелиновые Н. в. периферической и центральной нервной системы имеют нек-рые различия. В Н. в. периферической нервной системы слой периферической цитоплазмы сплошной и широкий, в нем располагается ядро нейролеммоцита, локализующееся посередине межузлового сегмента. В волокнах ц. н. с. (рис. 3) периферическая цитоплазма не образует сплошного слоя, а формирует узкий гребешок, тянущийся вдоль всего межузлового сегмента. В связи с этими особенностями слоя периферической цитоплазмы в нервных волокнах ц. н. с. степень выраженности наружного мезаксона незначительна. На внутренней поверхности миелиновой оболочки, располагающейся в этих Н. в. у осевого цилиндра, цитоплазмы также мало. Часто она, не образуя сплошного слоя, лежит на ограниченных участках по обе стороны мезаксона. Особенностью миелиновых Н. в. центральной нервной системы является отсутствие ядер нейролеммоцитов в наружном слое цитоплазмы сформированной миелиновой оболочки.
В цитоплазме нейролеммоцитов миелиновых Н. в. как в ц. н. с., так и в периферической нервной системе обнаруживается полный набор органелл, свойственный нейролеммоцитам безмиелиновых Н. в., и, кроме того, содержатся различные включения: пи-гранулы Рейха, тельца глиоцита (эльцгольцевские тельца), липидные гранулы.
Ультраструктура перехватов узла в миелиновых Н. в. как центральной, так и периферической нервной системы также одинакова. Перехват узла состоит из собственно узловой области (обнаженная часть осевого цилиндра) и двух околоузловых участков, образуемых булавовидно утолщенными отростками нейролеммоцитов, содержащими скопления цитоплазмы. Узловые области имеют протяженность 0,25—1,0 мкм и расположены друг от друга на расстоянии 0,5—2,0 мкм. Пластины, составляющие миелиновую оболочку в околоузловой области, оканчиваются в определенной последовательности: первыми оканчиваются пластины, лежащие ближе всего к осевому цилиндру, затем лежащие снаружи от Н. в. В месте окончания пластин главная оптически плотная линия каждого концентрического слоя расщепляется, и пластины миелина, направляясь к осевому цилиндру, отслаиваются под углом 45—135°. Между отщепляющимися и изменившими направление пластинами образуются свободные пространства, заполненные цитоплазмой. Т. о., в околоузловой области осевой цилиндр со всех сторон окружен цитоплазматическим тяжем, являющимся элементом спирально закрученного мезаксона. Мембраны, окружающие смежные пластины околоузлового цитоплазматического тяжа, могут образовывать между собой контакты по типу десмосом (см.). В околоузловой области плазмолемма осевого цилиндра отделена от мембраны нейролеммоцита щелью толщиной в 2,5—3,0 нм. Поскольку тяж цитоплазмы нейролеммоцита формирует ряд спиральных пластин, очертания осевого цилиндра имеют зубчатый контур, а его цитолемма образует утолщения (до 15 нм), располагающиеся на равном расстоянии друг от друга. На продольных срезах эти утолщения имеют вид плотных колец.
Ультраструктура межузловых сегментов Н. в. в периферической и центральной нервной системе имеет нек-рые различия. Так, в Н. в. периферической нервной системы осевой цилиндр покрыт переплетенными пальцевидными отростками нейролеммоцитов диам. 50 нм, к-рые представляют собой продолжение сегментов миелиновой оболочки, расположенной по обе стороны от перехвата узла. В нервных волокнах ц. н. с. аналогичные структуры отсутствуют. Места окончания пластин миелиновой оболочки при светооптическом изучении Н. в. периферической и центральной нервной системы на препаратах, импрегнированных солями серебра, выявляются как ряд темных структур, получивших название «двойные шиповидные браслеты Нажотта». На уровне перехватов узла осевой цилиндр несколько утолщается, в нем увеличивается число микротрубочек, митохондрий, везикул, сходных с синаптическими пузырьками, а также мультивезикулярных телец; под плазмолеммой располагается слой плотного зернистого вещества. Плотный слой отделен от цитолеммы промежутком в 10 нм, толщина плотного слоя 20 нм, структура его напоминает структуру плотного слоя начального сегмента аксона (см. Нервная клетка). На уровне перехвата узла может происходить разветвление миелинового нервного волокна. В ц. н. с. перехваты узла могут участвовать в образовании синапсов. В этих случаях аксон содержит образования пре- или постсинаитического комплекса (см. Синапс).
В Н. в. периферической нервной системы миелиновые оболочки межузловых сегментов прерываются рядом косых воронкообразных щелей, получивших название «насечки миелина» (насечки Шмидта—Лантерманна). Насечки миелина обнаруживаются в Н. в., фиксированных осмиевой к-той, и в нефиксированных Н. в.— при изучении их в поляризованном свете. Электронно-микроскопически показано, что в области насечек миелина происходит расщепление пластин миелиновой оболочки по ходу главной оптически плотной линии, а щель заполнена спиралевидно расположенной цитоплазмой нейролеммоцита. Размер щели 5— 19 нм, при прохождении ее через всю толщу миелиновой оболочки образуется канал, посредством к-рого цитоплазма, расположенная внутри миелиновой оболочки, сообщается с цитоплазмой, лежащей снаружи. Вид насечек миелина изменчив; возможно, это динамичные структуры, присутствие или отсутствие к-рых связано с функц, состоянием Н. в. Между глиальной оболочкой миелиновых Н. в. и соединительнотканными элементами эндоневрия имеется базальная мембрана, к-рая хорошо выражена в миелиновых Н. в. периферической нервной системы. В Н. в. центральной нервной системы базальная мембрана и эндоневрий отсутствуют, а межклеточная среда отделена от Н. в. отростками астроцитов (см. Нейроглия).
Физиология
В зависимости от направления проведения импульса различают центростремительные (афферентные, или чувствительные) Н. в., проводящие нервный импульс от рецепторов в ц. н. с., и центробежные (эфферентные, или двигательные) Н. в., передающие нервный импульс из ц. н. с. к исполнительным органам.
Важную роль в процессах проведения возбуждения по Нервному волокну играет цитолемма осевого цилиндра. Роль миелиновой оболочки в функциях Н. в. еще до конца не выяснена. Полагают, что она имеет важное значение в процессах, определяющих скорость проведения возбуждении по Н. в. Высказано также предположение, что нейролеммоциты, окружающие осевой цилиндр, снабжают его продуктами метаболизма, компенсируй тем самым удаленность участков осевого цилиндра от его собственного ядра.
Систематическое изучение физиологии Н. в. началось с середины 19 в. В изучение этой проблемы внесли существенный вклад Г. Гассер, Г. Гельмгольц, H. Е. Введенский, Б. Ф. Вериго, Б. Катц, Лоренте де Но (R. Lorente de No), А. А. Ухтомский, Э. Хаксли, А. Ходжкин, Э. Эдриан, Дж. Эрлангер и др. Благодари исследованиим этих ученых были открыты и сформулированы основные законы и особенности функционировании Н. в.: закон физиологической непрерывности, к-рый гласит, что возбуждение может распространиться по Н. в. только при сохранении его морфол, и физиол, целостности; закон изолированного проведении возбуждении, по к-рому распространившееся по Н. в. возбуждение не передается на другие волокна, находящиеся в том же нервном стволе; закон двустороннего проведении, по к-рому волна возбуждении, возникнув в какой-либо области Н. в., распространяется в обе стороны от очага возникновении.
Важными свойствами Н. в. являются возбудимость (см.) и проводимость. В естественных условиях Н. в. возбуждается в результате непосредственного действии механических, химических, температурных и других раздражителей на нервное окончание или тело нервной клетки. В эксперименте используют раздражение электрическим током. Так, ток напряжением 20—40 мв и силой 6‘10_10а уже способен вызвать в Н. в. одиночный цикл возбуждении. По Н. в. возбуждение (см.) распространяется в виде особого электрического сигнала — потенциала действия, или нервного импульса, возникающего менее чем через 0,5 мсек после действия раздражителя (см. Нервный импульс). Н. в. способно провести следующий импульс после предшествующего лишь через несколько миллисекунд. Это так наз. состояние рефрактерности, в к-ром можно выделить ряд фаз. Во время первой (фаза абсолютной рефрактерности) даже самый мощный стимул не может вызвать возбуждении Н. в. Во время второй (фаза относительной рефрактерности) Н. в. возбуждается только стимулом, превышающим порог раздражении. Затем наблюдается фаза повышенной возбудимости, к-рую H. Е. Введенский назвал экзальтационной. Весь цикл изменения возбудимости в Н. в. при прохождении одного импульса составляет 18—30 мсек.
В момент прохождения импульса по Н. в. активный участок волокна, на протяжения к-рого наблюдается отклонение потенциала от состояния покоя, носит название длины волны. Энергии, требуемой для передачи возбуждении, возникает в волокне и не зависит от силы стимула. Процесс возбуждении сопровождается выделением небольшого количества тепла (один импульс повышает температуру Н. в. на четыре миллионных градуса). Поддержание потенциала покои и восстановление возбудимости Н. в. после прохождения импульса осуществляется благодаря окислительным процессам, связанным с превращением глюкозы при гликолизе в цикле Кребса (см. Трикарбоновых кислот цикл). Минимальная сила тока, способная вызвать процесс возбуждении при неограниченно долгом действии постоянного тока на нервную ткань, называется реобазой. Минимальное время, в течение к-рого ток при силе, равной реобазе, должен действовать на ткань, чтобы вызвать распространяющийся ответ, называется полезным временем реобазного тока, или полезным временем. Графическая зависимость между длительностью стимула и пороговой интенсивностью носит название кривой сила — длительность. Эта зависимость связана со многими факторами, и в первую очередь со скоростью проведении возбуждении. Чем она больше, тем выше возбудимость Н. в. и слабее необходимый дли возбуждении ток. Производной от кривой сила — длительность является хронаксия (см. Хронаксиметрия).
Каждое Нервное волокно способно передавать возбуждение в обоих направлениях (закон двустороннего проведении): центростремительно, т. е. к нервным центрам, и центробежно — от центров. Простейшим доказательством двусторонней проводимости может служить раздражение средней части нерва, к-рое сопровождается возникновением потенциала действии, распространяющегося по обе стороны от места раздражении. В ц. н. с. передача возбуждении от одной нервной клетки к другой осуществляется через синапсы. Этот путь носит название ортодромного. Раздражение Н. в., примениемое в эксперименте дли изучении механизмов нервной деятельности, распространяется не только ортодромно, но и в обратном направлении — к телу клетки. Такой путь распространения возбуждении носит название антидромного.
Электрическая возбудимость Н. в. связана с наличием в цитолемме осевого цилиндра специализированных ионных каналов (натриевых, калиевых, кальциевых), способных «открываться» или «закрываться» в ответ на сдвиги мембранного потенциала. Ионные токи, текущие через эти каналы, обеспечивают генерацию как потенциалов покои, так и потенциалов действия.
С ионными токами связаны и другие явления, наблюдаемые в Н. в. Так, показано, что пороговая сила тока, способная вызвать потенциал действии в Н. в., зависит не только от длительности, но и от крутизны нарастании потенциала; при этом уменьшение крутизны ведет к повышению порога. В основе этого явления, названного Нернстом (W. Nernst, 1908) аккомодацией, лежит инактивация натриевой и активации калиевой проницаемости ионных каналов. С активацией или инактивацией цитолеммы осевого цилиндра к ионам калия связаны и изменении ее возбудимости: повышение возбудимости под катодом (катэлектротон) и снижение ее под анодом (анэлектротон), описанные впервые Э. Пфлюгером. Однако если мембранный потенциал полностью обеспечивается ионными токами, то в генезе распространяющегося потенциала действии важную роль играют так наз. локальные токи, возникающие между возбужденными и соседними невозбужденными участками Н. в. Это положение нашло отражение в предложенной в 1905 г. Германном (L. Hermann) так наз. кабельной теории, объясняющей механизм передачи возбуждении по Н. в. Теории получила экспериментальное обоснование в работах А. Ходжкина и Раштона (W. A. H. Rushton, 1946). Кабельная теория объясняет непрерывное проведение возбуждении по безмиелиновым и прерывистое его распространение по миелиновым Н. в. При непрерывном проведении возбуждении в безмиелиновых Н. в., обладающих однородным строением цитолеммы осевого цилиндра глиальных оболочек, в возбужденном ее участке повышается проницаемость дли ионов натрии, к-рые проникают внутрь Н. в. При этом происходит увеличение положительного заряда внутри Н. в., вызывающее перераспределение ионов в окружающей среде, вследствие чего возникают локальные токи. Эти токи создаются потоком любых ионов, значение имеет лишь знак заряда иона, в соответствии с к-рым ион движется. Локальные токи, распространяясь по обе стороны от возбужденного участка, вызывают деполяризацию соседних невозбужденных участков мембраны Н. в. и их возбуждение. От того, на какое расстояние распространяются локальные токи, способные возбудить волокно, зависит и скорость проведения возбуждения по Н. в. Исходя из кабельной теории, согласно к-рой увеличение сопротивления цитолеммы осевого цилиндра должно положительно сказываться на передаче возбуждения, становится понятным функц, значение миелинизации волокна. При увеличении числа изолирующих слоев миелина Н. в. сохраняет способность к возбуждению лишь на участках перехватов узла, в к-рых цитолемма осевого цилиндра Н. в. остается неизолированной. Такое прерывистое возбуждение, ограниченное областью межузлового сегмента, называют скачкообразным, или сальтаторным. В безмиелиновых Н. в. скорость распространения потенциала действия пропорциональна величине, равной корню квадратному из их диаметра. В миелиновых Н. в. при сальтаторном проведении импульса скорость его проведения становится пропорциональной не величине, равной корню квадратному диаметра Н. в., а диаметру Н. в. Такая прямая зависимость объясняется тем,что при определенном диаметре Н. в. скорость распространения потенциала действия зависит от длины межузловых сегментов.
В обычных условиях по Н. в. передаются не одиночные импульсы, а их серии. При этом в тех случаях, когда импульсы следуют друг за другом с большой частотой, т. е. когда частота стимуляции превышает величину рефрактерного периода, Н. в. полностью воспроизводит ритм стимуляции и проведение каждого импульса подчиняется законам, изложенным выше. В тех же случаях, когда период стимуляции меньше, чем длительность рефрактерного периода, проведение импульсов имеет нек-рые особенности, в частности наблюдается трансформация в Н. в. ритма стимуляции, к-рая выражается в неполном воспроизведении частот стимуляции.
Основываясь на этом, H. Е. Введенский ввел понятие «функциональная подвижность», или лабильность (см.), к-рая характеризуется максимальным числом импульсов, воспроизводимым Н. в. в 1 сек., в точном соответствии с ритмом подаваемых на него раздражений. Лабильность Н. в. можно изменить при воздействии на него наркотиками, сильным электрическим током, нагреванием, давлением и т. д. При этом в проведении возбуждения через измененный участок Н. в. обнаруживаются следующие закономерности. Сначала исчезают различия в действии слабых и сильных ритмических раздражителей — уравнительная стадия. Затем наступает парадоксальная фаза, при к-рой сильные раздражители не действуют, а слабые продолжают вызывать ответные реакции. Наконец, следует последняя фаза — тормозная, при к-рой отмечается полная непроводимость. Основываясь на этих фактах, H. Е. Введенским была сформулирована теория парабиоза (см.), согласно к-рой при длительном и непрерывном воздействии на нерв любого раздражения в Н. в. создается очаг стойкого неколеблющегося возбуждения, ограниченного местом возникновения.
С различием в диаметре Н. в. стволов периферической нервной системы связаны и различия в форме потенциала действия, а также в скорости проведения возбуждения; в ответ на раздражение нервного ствола регистрируется потенциал действия сложной формы, в к-ром выделяют три основных пика — А, В, С. Связь между этими компонентами сложного потенциала действия и разными группами волокон была выяснена Дж. Эрлангером и Г. Гассером (1937). По их данным, за возникновение пика А ответственны толстые миелиновые волокна с диам. от 3 до 22 мкм и скоростью проведения возбуждения от 12 до 120 м/сек (волокна типа А). Дальнейшее изучение волокон типа А. позволило по скорости проведения возбуждения и диаметру разбить их на подгруппы: альфа (двигательные волокна скелетных мышц, афферентные волокна от мышечных рецепторов), бета (афферентные волокна от тактильных рецепторов), гамма (афферентные волокна от тактильных рецепторов, барорецепторов, эфферентные волокна к мышечным веретенам) и дельта (афферентные волокна от нек-рых терморецепторов, механорецепторов, болевых рецепторов). К волокнам, ответственным за возникновение пика В (волокнам типа В), относят миелиновые Н. в. средней толщины (диам. 1 — 3 мкм) и скоростью проведения возбуждения в преганглионарных стволах 3—14 м/сек. К Н. в. группы С относятся безмиелиновые волокна (постганглионарные вегетативные волокна, афферентные волокна от нек-рых болевых, механо- и терморецепторов) толщиной не более 2 мкм и скоростью проведения 1—2 м/сек.
Физические и химические воздействия оказывают различное влияние на разные группы Н. в. Так, напр., толстые миелиновые Н. в. особенно чувствительны к механическим повреждениям и менее чувствительны по сравнению с тонкими волокнами того же ствола к наркотикам и анестетикам. Асфиксия нервного ствола, вызванная нарушением кровоснабжения путем наложения жгута, блокирует проведение возбуждения по толстым миелиновым волокнам раньше, чем по тонким, проводящим болевую чувствительность. Однако, несмотря на многочисленные попытки, до сих пор не удалось установить связь между характером ощущений и типом Н. в., его передающего. Так, напр., ощущение боли может передаваться как но тонким миелиновым волокнам, так и по безмиелиновьш волокнам типа С. Патоморфология

При воздействии повреждающих факторов на Н. в. могут развиваться патол, изменения во всех его компонентах: осевом цилиндре, миелиновой оболочке, нейро-леммоцитах (цветн. рис. 3—6). Полное разрушение Н. в. может возникнуть первично, при непосредственном воздействии повреждающего агента на определенный участок нервной ткани (тяжелая травма, ишемия и др.) или вторично, в случае нарушения связи волокна с телом клетки (вторичное перерождение). В ряде случаев повреждению подвергается только миелиновая оболочка при относительной сохранности осевых цилиндров (периаксональная демиелинизация).

Процесс, возникающий в проксимальном отрезке Н. в. при перерыве последнего, носит название ретроградной дегенерации. На концах прерванных аксонов появляются шаровидные и колбовидные вздутия с разрыхлением нейрофибрилл — так наз. ретракционные колбы Рамон-и-Кахаля. С помощью гистохим, методов исследования в эксперименте показано, что при перерезке Н. в. в аксоне выше места перерезки происходит массивное накопление медиатора. Отрезок Н. в., прилежащий к месту повреждения, подвергается полному распаду, а в более проксимальном его отделе (по отношению к месту повреждения) наблюдается распад миелиновой оболочки (рис. 4). Регенерация Н. в. начинается с увеличения объема нейролеммоцитов и пролиферации их в виде тяжа. Новые нейрофибриллы из утолщенной части аксона в виде небольших аксо-плазматических выростов начинают появляться спустя 7 дней после раздавливания нерва и на 14-й день после перерезки его. К 8-й нед. они приобретают вид безмиелиновых Н. в. Ход этих волокон не всегда прямолинеен, они могут располагаться веерообразно, в виде клубков или иметь обратный ход. При наличии на их пути препятствий (напр., рубца) может образоваться так наз. ампутационная неврома, нередко являющаяся причиной болевого синдрома.



Сегментарная демиелинизация в периферической нервной системе является обратимым процессом. Морфол. признаком ремиелинизации является несоответствие между тонким слоем миелиновой оболочки и большим диаметром аксона. В ц. н. с., по данным Ю. М. Жаботинского и В.П. Иоффе (1975), Райне (С. S. Raine) с соавт. (1969), Ламперта (Н. Lampert, 1965), ремиелинизация происходит не полностью и очень медленно.

При ряде прогрессирующих заболеваний нервной системы различного генеза обнаруживается патология Н. в. с преимущественным поражением определенных проводящих систем; при амиотрофическом боковом склерозе (см.) основные изменения выявляются в пирамидных путях, при семейной атаксии Фридрейха (см. Атаксия) — в задних столбах спинного мозга и снино-церебеллярных путях, при спинной сухотке (см.) — в задних столбах спинного мозга и т. д. При этом далеко не всегда изменения можно связать с поражением тела нейроцита, т. е. провести аналогию с валлеровским перерождением. Поэтому такие поражения иногда называют первичными. Предполагается, что в подобных случаях происходит не только распад миелина, но и нарушение его синтеза, обусловленное метаболическими сдвигами. В случаях с относительно быстрым течением заболевания (напр., амиотрофический боковой склероз) в участках демиелинизации выявляются продукты жирового распада; при длительном течении заболевания (болезнь Фридрейха) можно констатировать лишь отсутствие миелина и реактивный глиоз (см.). Осевые цилиндры при этом повреждаются, но в более поздние сроки и менее грубо, чем миелиновые оболочки. При нек-рых так наз. дегенеративных заболеваниях ц. н. с. (болезнь Галлервордена — Шпатца, инфантильная нейроаксональная белковая дистрофия), опухолях (напр., при плазмоцитоме) и авитаминозах возможна патология Н. в. с преимущественным поражением аксона. Последний увеличивается в объеме и подвергается прерывистому распаду с образованием сфероидов (аксональных шаров), состоящих из гомогенного вещества, иногда содержащих аргентофильные гранулы (рис. 8).
Из других видов патологии Н. в. следует отметить отложение амилоида в Н. в. при общем амилоидозе; значительное утолщение миелиновой оболочки при нек-рых пороках развития головного мозга и истончение миелиновой оболочки при атрофических процессах в старческом возрасте, хрон, энцефалитах и нек-рых других заболеваниях.
Глиальная оболочка (шванновская оболочка) периферического Н. в. может явиться источником опухолевого роста; возникающая при этом доброкачественная опухоль носит название невриномы (см.), неврилеммомы, шванномы.
При изучении Нервных волокон применяются нейрогистологические методики, позволяющие оценить состояние миелиновой оболочки и осевого цилиндра. Для оценки состояния миелиновой оболочки используют метод Вейгерта (см. Вейгерта методы окраски) и ряд его модификаций: метод Шпильмейера, метод Кульчицкого, а также комплексную окраску лаксолевым синим прочным. Эти методики применяют для определения локализации поражения Н. в., для исследования же изменения отдельных Н. в. используют импрегнацию по методу Марки, реакции с жирорастворимыми красителями (судан, шарлах, нильский голубой), что позволяет выявить нейтральные жиры. Для исследования осевых цилиндров и безмиелиновых волокон используют различные методы импрегнации серебром (см. Бильшовского-Грос-Лаврентьева метод, Серебрения методы). См. также Нервная система, Нервы.
Библиография: Бреже М. Электрическая активность нервной системы, пер. с англ., М., 1979; Дойников Б. С. Избранные труды по неыроморфологии и невропатологии, с. 114, М., 1955; Жаботинский Ю. М. Нормальная и патологическая морфология нейрона, Л., 1965, библиогр.; Жаботинский Ю. М. и Иоффе В. И. Экспериментальные аллергические Демиелинизирующие заболевания нервной системы, с. 90, 158, Л., 1975; Жаботинский Ю. М. и Косте нецкий А. С. Регенерация поврежденных периферических нервов при ишемии, в кн.: Вопр, физиол, и морфол, центр, нервн. сист., под ред. К. М. Быкова, с. 208, М., 1953; Жуков Е. К. Очерки по нервно-мышечной физиологии, Л., 1969; Катц Б. Нерв, мышца и синапс, пер. с англ., М., 1968; Куффлер С. и Николс Дж. От нейрона к мозгу, пер. с англ., М., 1979, библиогр.; Латманизова Л. В. Очерк физиологии возбуждения, М., 1972; Леонович А. Л. Прогрессирующие лейко-энцефалиты, с. 97, М., 1970; Немечек С. и др. Введение в нейробиологию, пер. с чешек., Прага, 1978; Общая физиология нервной системы, под ред. П. Г. Костюка, Л., 1979; Оке С., Основы нейрофизиологии, пер. с англ., М., 1969] Питерс А., Палей С. и Уэбстер Г. Ультраструктура нервной системы, пер. с англ., М., ‘1972, библиогр.; Сотников О. С.; Функциональная морфология живого мякотного нервного волокна, Л., 1976, библиогр.; Bunge R. F. Glial cells and central myelin sheath, Physiol. Rev.. v. 48, p. 197, 1968, bibliogr.; Lampert P.Demyelination and remyelination in experimental allergic encephalomyelitis, J. Neuropath. exp. Neurol., v. 24, p. 371, 1965; он же, Mechanism of demyelination in experimental allergic neuritis, Lab. Invest., v. 20, p. 127, 1969; Lee J. C. Electron microscopy of Wallerian d-sgeneration, J. comp. Neurol., v. 120, p. 65, 1963; Ohmi S. Electron microscopic study on Wallerian degeneration of the peripheral nerv, Z. Zellforsch., Bd 54, S. 39, 1961; Raine G. S. a. o. An ultrastruetural study of experimental demyelination and remyelination, Lab. Invest., v. 25, p. 28, 1971; Robertson J. D. The ultrastructure of adult vertebrate peripheral myelinated nerve fibers in relation to myelinogenesis, J. biophys, biochem. Cytol., v. 1, p. 271, 1955; Schlote W. Nervus opticus und experimentelles Trauma, B., 1970; Seitelberger F. Zentrale pontine Myelinolyse, Schweiz. Arch. Neurol. Psychiat., Bd 112, S. 285, 1973; The structure and function of nervous tissue, ed. by G. H. Bourne, p. 325, N. Y., 1968; Suzuki K. a. o. Ultrastruetural studies of multiple sclerosis, Lab. Invest., v. 20, p. 444, 1969.
И. А. Чернова; Т. И. Вавилова, Р. П. Чайковская (патоморфология), А. Д. Ноздрачев (физиол.).



