Нуклеопротеиды
К нуклеопротеидам относятся устойчивые комплексы нуклеиновых кислот с белками, длительное время существующие в клетке в составе органелл или структурных элементов клетки в отличие от разнообразных короткоживущих промежуточных комплексов белок — нуклеиновая кислота (комплексы нуклеиновых кислот с ферментами — синтетазами и гидролазами — при синтезе и деградации нуклеиновых кислот, комплексы нуклеиновых кислот с регуляторными белками и т. п.).
Содержание
Структура и устойчивость
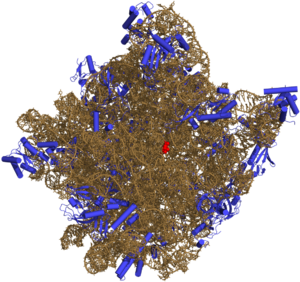

В зависимости от типа входящих в состав нуклеопротеидных комплексов нуклеиновых кислот различают рибонуклеопротеиды и дезоксирибонуклеопротеиды.
Устойчивость нуклеопротеидных комплексов обеспечивается нековалентным взаимодействием. У различных нуклеопротеидов в обеспечение стабильности комплекса вносят вклад различные типы взаимодействий, при этом нуклеиново-белковые взаимодействия могут быть специфичными и неспецифичными. В случае специфичного взаимодействия определённый участок белка связан со специфичной (комплементарной участку) нуклеотидной последовательностью, в этом случае вклад водородных связей, образующихся между нуклеотидными и аминокислотными остатками благодаря пространственному взаимному соответствию фрагментов, максимален. В случае неспецифичного взаимодействия основной вклад в стабильность комплекса вносит электростатическое взаимодействие отрицательно заряженных фосфатных групп полианиона нуклеиновой кислоты с положительно заряженными аминокислотными остатками белка.
Примером специфичного взаимодействия могут служить нуклеопротеидные комплексы рРНК — субъединицы рибосом; неспецифичное электростатическое взаимодействие характерно для хромосомных комплексов ДНК — хроматина и комплексов ДНК-протамины головок сперматозоидов некоторых животных.
Нуклеопротеиды диссоциируют на белки и нуклеиновые кислоты при воздействии агентов, разрушающих или ослабляющих нековалентные связи:
Некоторые нуклеопротеиды (рибосомные субчастицы, нуклеокапсиды вирусов) обладают способностью к самосборке, то есть к образованию, при соответствующих условиях, нуклеопротеидов in vitro без участия клеточных структур или агентов; такая самосборка возможна в случае специфичных нуклеиново-белковых взаимодействий (нуклеиново-белковым узнавании). В любом случае, при образовании нуклеопротеидов происходят существенные конформационные изменения нуклеиновых кислот и, в некоторых случаях, белков, образующих нуклеопротеидный комплекс.
Распространённость и биологическая роль
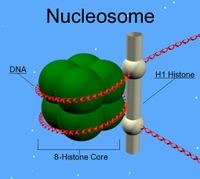
Наиболее сильные конформационные изменения при образовании нуклеопротеидов претерпевают нуклеиновые кислоты, и эти изменения наиболее существенны в случае образования дезоксирибонуклеопротеидов. В отличие от одноцепочечной РНК, способной образовывать вторичные и третичные структуры за счёт антипараллельного комплементарного спаривания смежных отрезков цепи. Двухцепочечная ДНК такой возможности не имеет и существует в растворах в виде значительно более «рыхлых», по сравнению с компактными глобулами РНК, клубков. Однако связывание ДНК с сильноосновными белками (гистонами и протаминами) за счёт электростатического взаимодействия приводит к значительно более плотно упакованным нуклеопротеидным комплексам — хроматинам, обеспечивающим компактное хранение ДНК и, соответственно, наследственной информации в составе хромосом эукариот. С другой стороны, большая конформационная подвижность РНК и её каталитические свойства приводят к большому разнообразию рибонуклеопротеидов, выполняющих различные функции.
Дезоксирибонуклеопротеиды
Рибонуклеопротеиды
В клетках в наибольших количествах содержатся два класса рибонуклеопротеидов:
Нуклеокапсиды вирусов
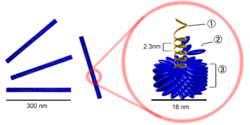
Нуклеокапсиды вирусов представляют собой достаточно плотно упакованные комплексы белков с нуклеиновой кислотой (ДНК или РНК у ретровирусов) и как функционально, так и структурно близки хроматину, представляя собой компактную форму вирусного генома.
Существует два основных типа нуклеокапсидных структур: палочковидная (нитевидная), либо сферическая («изометрическая»).
В первом случае связанные белковые субъединицы периодически располагаются вдоль нити нуклеиновой кислоты таким образом, что она сворачивается в спираль, образуя своего рода «инвертированную нуклеосому», в которой, в отличие от нуклеосом эукариот, белковая часть расположена не внутри, а снаружи структуры. Такая структура нуклеокапсидов типична для вирусов растений (в частности, вируса табачной мозаики) и миксо-, парамиксо- и рабдовирусов, нуклеокапсиды которых имеют спиральную форму.
В изометрических структурах упаковка нуклеиновой кислоты вирусного генома более сложна: белки оболочки нуклеокапсида относительно слабо связаны с нуклеиновой кислотой или нуклеопротеидами, что налагает минимум ограничений на способ упаковки нуклеиновой кислоты. Нуклеопротеиды «сердцевины» при этом могут быть весьма сложно организованы: так, в паповавирусах двухцепочечная кольцевая ДНК, связываясь с гистонами, образует структуры, очень похожие на нуклеосомы.
НУКЛЕОПРОТЕИДЫ
нуклеопротеи́ды, сложные белки, состоящие из простых белков и нуклеиновых кислот, содержащиеся в организмах. В зависимости от входящей в их состав нуклеиновой кислоты Н. подразделяют а дезоксирибонуклеопротеиды (ДНП) и рибонуклеопротеиды (РНП). Белковая часть Н. представлена главным образом гистонами и протаминами. ДНП основная составная часть ядер клеток. РНП входят главным образом в состав рибосом. Биологическая роль Н. связана с участием входящих в их состав нуклеиновых кислот в процессах биосинтеза белка, передачи наследственности и др.
Смотреть что такое «НУКЛЕОПРОТЕИДЫ» в других словарях:
Нуклеопротеиды — Нуклеопротеиды комплексы нуклеиновых кислот с белками. К нуклеопротеидам относятся устойчивые комплексы нуклеиновых кислот с белками, длительное время существующие в клетке в составе органелл или структурных элементов клетки в отличие от… … Википедия
НУКЛЕОПРОТЕИДЫ — комплексы белков с нуклеиновыми кислотами. Составляют основу заключенного в ядрах клеток наследственного вещества хроматина, образуют многие вирусы, рибосомы, информосомы … Большой Энциклопедический словарь
НУКЛЕОПРОТЕИДЫ — сложные комплексы нуклеиновых к т с белками. По характеру нуклеиновой к ты, входящей в состав Н., различают дезоксирибонуклеопротеиды (ДНП) и рибонуклеопротеиды (РНП). ДНП содержатся в ядрах всех клеток (вещество хромосом), митохондриях и… … Биологический энциклопедический словарь
НУКЛЕОПРОТЕИДЫ — (нуклеопротеины), сложные белки, в к рые в качестве небелковой части (простетической группы) входят нуклеиновые к ты, в виде которых находится весь фосфор нуклеопротеидов. Н. содержатся преимущественно в ядрах клеток, составляя главную массу их.… … Большая медицинская энциклопедия
НУКЛЕОПРОТЕИДЫ — комплексы нуклеиновых к т с белками. Содержатся в каждой клетке и выполняют важные ф ции, связанные с хранением и реализацией генетич. информации. Н. образуются с участием как ДНК (дезоксирибо нуклеопротеиды, или ДНП), так и РНК (рибонуклеопротеи … Химическая энциклопедия
нуклеопротеиды — комплексы белков с нуклеиновыми кислотами. Составляют основу заключённого в ядрах клеток наследственного вещества хроматина, образуют многие вирусы, рибосомы, информосомы. * * * НУКЛЕОПРОТЕИДЫ НУКЛЕОПРОТЕИДЫ, комплексы белков с нуклеиновыми… … Энциклопедический словарь
нуклеопротеиды — (лат. nucleus ядро + протеиды) сложные комплексы, состоящие из нуклеиновых кислот в простых белков (протеинов); входят в состав ядер в цитоплазмы всех животных и растительных клеток, микроорганизмов и вирусов. Новый словарь иностранных слов. by… … Словарь иностранных слов русского языка
Нуклеопротеиды — комплекс белков с нуклеиновыми кислотами (ДНК или РНК). Нуклеопротеиды образуют рибосомы, информосомы (внутриклеточные частицы в эука риотах) и вирусы … Начала современного естествознания
Нуклеопротеиды — широко распространённые в природе комплексы нуклеиновых кислот (См. Нуклеиновые кислоты) с белками. В зависимости от характера входящей в состав Н. нуклеиновой кислоты различают дезоксирибонуклеопротеиды (ДНП) и рибонуклеопротеиды (РНП).… … Большая советская энциклопедия
нуклеопротеиды — нуклеопроте иды, ов, ед. ч. е ид, а … Русский орфографический словарь
НУКЛЕОПРОТЕИДЫ
турируются. Почти все вращают плоскость поляризации вправо. Содержание фосфора в нуклеоп роте идах—0,5—6,1%, в нуклеинах— ДО 5% И более. В. Энгелъгардт.
NUCLEOPHAGA,мельчайшие п- топлазму амебы. Разру- шение ядра не сразу отражается на жизнедеятельности зараженной амебы. Дофлейн (Dof-lein) наблюдал амебоидное движение Amoeba vespertilio, у которой все ядро было разрушено Nucleophaga (рис. 2). Наблюдались случаи поражения кариосомы паразитом. Описано несколько форм N., но видовая самостоятельность их не ясна. Примеры: Nucleophaga amoebae, Nucleophaga in-testinalis и др. В случаях нахождения N. в паразитических амебах можно говорить об явлении сверхпаразитизма. Лит.: ЭпщтейнГ., К вопросу о параз§тах у простейших, Архив Рус. протистол. об-ва, т. I, 1922.
Полезное
Смотреть что такое «НУКЛЕОПРОТЕИДЫ» в других словарях:
Нуклеопротеиды — Нуклеопротеиды комплексы нуклеиновых кислот с белками. К нуклеопротеидам относятся устойчивые комплексы нуклеиновых кислот с белками, длительное время существующие в клетке в составе органелл или структурных элементов клетки в отличие от… … Википедия
НУКЛЕОПРОТЕИДЫ — комплексы белков с нуклеиновыми кислотами. Составляют основу заключенного в ядрах клеток наследственного вещества хроматина, образуют многие вирусы, рибосомы, информосомы … Большой Энциклопедический словарь
НУКЛЕОПРОТЕИДЫ — сложные комплексы нуклеиновых к т с белками. По характеру нуклеиновой к ты, входящей в состав Н., различают дезоксирибонуклеопротеиды (ДНП) и рибонуклеопротеиды (РНП). ДНП содержатся в ядрах всех клеток (вещество хромосом), митохондриях и… … Биологический энциклопедический словарь
НУКЛЕОПРОТЕИДЫ — комплексы нуклеиновых к т с белками. Содержатся в каждой клетке и выполняют важные ф ции, связанные с хранением и реализацией генетич. информации. Н. образуются с участием как ДНК (дезоксирибо нуклеопротеиды, или ДНП), так и РНК (рибонуклеопротеи … Химическая энциклопедия
нуклеопротеиды — комплексы белков с нуклеиновыми кислотами. Составляют основу заключённого в ядрах клеток наследственного вещества хроматина, образуют многие вирусы, рибосомы, информосомы. * * * НУКЛЕОПРОТЕИДЫ НУКЛЕОПРОТЕИДЫ, комплексы белков с нуклеиновыми… … Энциклопедический словарь
нуклеопротеиды — (лат. nucleus ядро + протеиды) сложные комплексы, состоящие из нуклеиновых кислот в простых белков (протеинов); входят в состав ядер в цитоплазмы всех животных и растительных клеток, микроорганизмов и вирусов. Новый словарь иностранных слов. by… … Словарь иностранных слов русского языка
Нуклеопротеиды — комплекс белков с нуклеиновыми кислотами (ДНК или РНК). Нуклеопротеиды образуют рибосомы, информосомы (внутриклеточные частицы в эука риотах) и вирусы … Начала современного естествознания
Нуклеопротеиды — широко распространённые в природе комплексы нуклеиновых кислот (См. Нуклеиновые кислоты) с белками. В зависимости от характера входящей в состав Н. нуклеиновой кислоты различают дезоксирибонуклеопротеиды (ДНП) и рибонуклеопротеиды (РНП).… … Большая советская энциклопедия
нуклеопротеиды — нуклеопроте иды, ов, ед. ч. е ид, а … Русский орфографический словарь
НУКЛЕОПРОТЕИДЫ — нуклеопротеиды, сложные белки, состоящие из простых белков и нуклеиновых кислот, содержащиеся в организмах. В зависимости от входящей в их состав нуклеиновой кислоты Н. подразделяют а дезоксирибонуклеопротеиды (ДНП) и рибонуклеопротеиды (РНП).… … Ветеринарный энциклопедический словарь
НУКЛЕОПРОТЕИДЫ
Нуклеопротеиды — комплексы нуклеиновых кислот с белками. Н. являются основным компонентом Многих биоструктур — хромосом (см.), рибосом (см.), вирусов (см.), информосом и др.
Образование нуклеопротеидов обеспечивает осуществление важнейших функций нуклеиновых кислот (см.) — воспроизведение, передачу и реализацию генетической информации. В зависимости от типа нуклеиновой к-ты, входящей в состав комплекса, различают дезоксирибонуклеопротеиды (ДНП) и рибонуклеопротеиды (РНП). Белки, образующие Комплексы с нуклеиновыми к-тами, могут быть разделены на две группы.
Одни связываются электростатическими связями преимущественно с двухспиральными полинуклеотидами и тем самым стабилизируют их вторичную структуру, другие, сродство к-рых к односпиральным полинуклеотидам выше, чем к двухспиральным, при связывании с двухспиральными полинуклеотидными цепями дестабилизируют их вторичную структуру. К последним относятся белки гена 5 фага fd, гена 32 фага Т4, фермент рибонуклеаза, белок SI, выделенный из рибосом и дестабилизирующий вторичную структуру РНК, и др. Нек-рые из белков, дестабилизирующих двухнитевые нуклеиновые к-ты, имеют полидоменную структуру. Каждый домен отвечает за взаимодействие с определенным лигандом. На процесс комплексиро-вания полинуклеотидной цепи с белками значительное влияние оказывает также топологическая организация молекул нуклеиновых к-т (напр., линейная, кольцевая или сверхспиральная форма ДНК). Ряд структурных, регуляторных белков, в т. ч. и ферментов, имеют разное сродство к специфическим последовательностям в молекулах ДНК, находящихся в сверхспиральной или релаксированной формах.
В ряде случаев определяющим моментом при взаимодействии нуклеиновых к-т с белками является наличие специфичных нуклеотидных последовательностей в полинуклеотидной цепи. При этом величины констант ассоциации белка со специфичными последовательностями нуклеотидов в молекуле нуклеиновой к-ты во много раз выше, чем величины констант ассоциации этого же белка с любыми другими нуклеотидными последовательностями. Это относится, напр., к нуклеопротеиду, представляющему собой участок молекулы ДНК, характеризующийся специфическими нуклеотидными последовательностями (оператор), с бел-ком-репрессором, блокирующим инициацию транскрипции соответствующих матричных РНК. Предполагают, что специфичность процесса образования этого комплекса обусловлена возникновением водородных связей между остатками определенных аминокислот в молекуле белка-репрессора и функциональными группами азотистых оснований оператора. Кроме того, комплекс стабилизируется электростатическими взаимодействиями между фосфатными группами ДНК и основными группами белка-репрессора, а в пределах последнего и белок-белковыми контактами. Взаимодействие белка-репрессора с денатурированной ДНК, а также измененным в результате мутаций оператором менее эффективно. Принципиальное значение имеет то, что в первичной структуре многих операторов обнаружены симметричные последовательности нуклеотидов. Это теоретически разрешает перестройку оператора из двойной спирали в структуру типа «шпильки», к-рая, вероятно, и узнается белком. Это предположение в некоторой степени подтверждается большими величинами констант ассоциации белка-репрессора с кольцевой ДНК, находящейся в сверхспиральной форме.
Взаимодействие между компонентами Нуклеопротеидов имеет также характерные особенности в зависимости от структурно-функционального назначения нуклеопротеидного комплекса в естественных биоструктурах. Напр., в вирусах сила и природа связи взаимодействующих групп Н. существенно зависят от размеров, хим. состава, типа симметрии белковой капсиды, структуры белковых субъединиц и вида нуклеиновой к-ты (ДНК или РНК) вируса. Так, у сферических вирусов в непосредственном контакте с молекулами белка находятся только периферические участки нуклеиновой к-ты, следствием чего является минимальный контакт между этими соединениями. В палочкообразных или нитевидных вирусах нуклеиновая к-та на всем своем протяжении погружена в белок, что приводит к более выраженному взаимодействию между ними. Взаимодействие между компонентами Н. является важным фактором, определяющим биол, свойства вирионов, их инактивацию, чувствительность к мутагенам и т. д.
Существенное значение для образования про-матричной РНК (про-мРНК) и ее транспорта от места синтеза к ядерной мембране имеют сложные полисомоподобные РНП-комплексы, образующиеся из молекулы про-мРНК и последовательно расположенных на ней белковых структур — информофер. В образовании этих комплексов заметную роль играют электростатические силы. Подобный тип взаимодействий имеет место и в информосомах — структурах, локализованных в цитозоле и представляющих собой сложные комплексы между матричной РНК и несколькими полипептидами.
Еще одним представителем Н. являются рибосомы — внутриклеточные структуры, в к-рых осуществляется синтез белков. Разные по первичной структуре и организации участки рибосомной РНК определяют расположение белковых субъединиц в субчастицах рибосом. При этом сам акт комплексирования приводит к структурной реорганизации определенных нуклеотидных последовательностей РНК. Большинство рибосомных белков имеет основный характер, что обусловлено высоким содержанием в них аргинина и лизина, что подчеркивает значительный вклад электростатических взаимодействий в образование комплекса РНК-белок. Это подтверждается диссоциацией относительно дискретных групп белков под действием высоких концентраций солей одновалентных металлов до почти полной депротеинизации рибосомных РНК. Однако этот тип взаимодействий не исчерпывает всех связей между РНК и белками в рибосомах. Так, напр., нитрирование остатков тирозина, а также модификация остатков лизина или метионина в соответствующих белках рибосом практически полностью нарушает РНК-белковые взаимодействия. Основные принципы нуклеиново-белкового «узнавания» в рибосомах, да и в других РНП-комплексах пока недостаточно выяснены.
В передаче и реализации генетической информации в клетках эукариотов основную роль играют нуклеиновые к-ты. Наиболее полно изучены комплексы ДНК с гистонами (см.) и протаминами (см.). Молекулы протаминов состоят приблизительно из 45 аминокислотных остатков, почти 3/4 из них являются остатками аргинина. Однако в подавляющем большинстве клеток эукариотов главным белковым компонентом Н. являются гистоны. Их молекулы содержат ок. 25% основных аминокислот (преимущественно аргинина и лизина). По электрофоретической подвижности гистоны делят на 5 основных фракций — Н1, Н2а, Н2в, НЗ, Н4, причем белки этих фракций отличаются друг от друга и по соотношению остатков лизина и аргинина в молекулах. В пределах одной фракции молекулы гистонов могут различаться по количеству и месту раположения модифицированных аминокислот (ацетилированных, метилированных, фосфо-рилированных, рибозилированных). Нек-рые фракции гистонов имеют подфракции. В клетках с репрессированным геномом фракция Н1 иногда замещается на гистон Н5, в состав нуклеогистона входит белок А24, представляющий собой гистон Н2а, ковалентно связанный с полипептидом убихитином. Поскольку в митозе белок А24 отсутствует, выдвигается предположение, что белок А24 является одним из факторов, препятствующих митотической конденсации ДНП в интерфазе. Молекулы гистонов и (или) их комплексы друг с другом соединяются с ДНК преимущественно за счет электростатических связей между отрицательно заряженными фосфатами ДИК и положительно заряженными группами остатков основных аминокислот гистонов. Возникающая при этом структура комплекса периодична, повторяющийся элемент — нуклеосома содержит в зависимости от источника выделения Н. ДНК длиной от 160 до 240 нуклеотидных пар и по две молекулы каждого из гистонов Н2а, Н2в, НЗ, Н4. В нуклеосоме выделяют основную часть («сердцевину»), содержащую вне зависимости от источника выделения Н. постоянное количество ДНК (140—145 пар нуклеотидов) и 8 молекул гистонов (так наз. октамер — по две молекулы каждого гистона: Н2а, Н2в, НЗ, Н4), ассоциированных друг с другом при помощи гидрофобных взаимодействий своих неполярных блоков. Моделью сердцевины нуклеосомы может служить диск диаметром 11 нм и толщиной 5,7 нм, причем ДНК в B-форме в виде спирали диаметром 9 нм и шагом 2,8 нм навивается на белковое ядро. Основные части нук-леосом соединены между собой сегментами ДНК, не входящими в основные частицы. Длина этого соединительного участка ДНК — «линкера» равна разности между длинами ДНК всей нуклеосомы и ее основной части. Т. о. вариация длин линкеров обусловливает гетерогенность нуклеосом по количеству содержащейся в них ДНК. Возможно, длина линкера определяется природой связанных с ним гистона Н1 (одна молекула на одну нуклеосому) и негистоновых белков.
Негистоновые белки хроматина (см.) весьма гетерогенны по своему составу; с помощью электрофореза в зависимости от типа клеток идентифицируется от нескольких десятков до сотен отдельных фракций таких белков. Их мол. веса (массы) составляют от 5000 до 200 000. Изоэлект-рические точки находятся в диапазоне от 3 до 10. Негистоновые белки Н. содержат большое количество аспарагиновой и глутаминовой к-т. Они обмениваются значительно быстрее, чем гистоны, и в отличие от них синтезируются в течение всего митотического цикла. Связанные с нуклеосомами негистоновые бел ки представляют собой относительно низкомолекулярные белки, содержащие большое количество диамино-и дикарбоновых аминокислот, отличающиеся высокой электрофоретической подвижностью, поэтому в литературе они обозначаются как НМ G-белки (англ. high mobility group). У нек-рых белков этой группы (НМ G 1 и НМ G 2) отмечено различие в сродстве к сверхсииральной и релаксированной формам ДНК. При обработке ДНП непосредственно в ядрах клеток ферментами, гидролизующими преимущественно те участки ДНК, к-рые представляют собой активные в данной клетке гены, из ДНП освобождаются нек-рые негистоновые белки. Это позволяет считать, что негистоновые белки (преимущественно НМ G 14 и НМ G 17) связаны с нуклеосомами функционально активного хроматина.
Нуклеосомы представляют собой динамические образования: при соответствующих параметрах среды, когда электростатическое отталкивание превышает силы гидрофобного взаимодействия, нуклеосомы разворачиваются. При механическом растяжении длина фибриллы ДНП увеличивается, и характерная для нуклеосом картина при рентгеноструктурном анализе исчезает. Введение в среду агентов, являющихся донорами межмолекулярных связей (напр., поликатионов), приводит к фиксации структуры открытых нуклеосом. Появление в среде соединений, конкурирующих за межмолекулярную связь (преимущественно, полианионов), вновь вызывает конденсацию ДНП. Изменения структуры нуклеосом указывают на существование полиморфизма (топомор-физма) ДНП уже на уровне упаковки этих образований. Возможность выбора из ряда альтернативных структур ДНП является, по-видимому, одной из основ эпигенеза, ибо фенотип можно рассматривать как возможность выбора из нескольких путей реализации информации, передаваемой через ДНК хромосом. Принципиальным является то, что образование нуклеосом сопровождается сверхспирализацией замкнутой ДНК. Следует учесть, что сверхспираль всегда отрицательна, т. е. противоположна знаку двойной спирали. С увеличением плотности витков сверхспирали начинают раскрываться достаточно протяженные области ДНК, имеющие, по-видимому, функциональное значение, о чем свидетельствует тот факт, что частота инициации транскрипции с увеличением плотности сверхспирали возрастает. Существенно, что сверхспи-рализация стимулирует и рекомбинацию. Предполагают, что в самом механизме генетических рекомбинаций заложена возможность их регуляции через изменение структурной организации ДНП. Перевод ДНК из сверхспиральной формы, индуцированной образованием нуклеосом, в релаксированную может осуществляться одним из ферментов группы топоизомераз, активность к-рого регулируется путем его взаимодействия с гистоном Н1.
Данные об организации нуклеосом получены на препаратах ДНП, выделенных из естественных биоструктур, однако полагают, что и в клетках ДНП организованы сходным образом. При наличии в составе ДНП гистона Н1 нуклеосомы плотно прилегают друг к другу, образуя фибриллу диаметром ок. 10 нм. Тот факт, что коэффициент линейной упаковки ДНК в нуклеосоме равен примерно 7, а коэффициент линейной упаковки ДНК метафазной хромосомы составляет величину порядка 104, указывает на наличие уровней организации ДНП более высоких, чем уровень организации нуклеосом. Встречающиеся в интерфазных ядрах клеток фибриллы ДНП диаметром 20—30 нм могут быть получены путем суперспиральной упаковки фибриллы диаметром 10 нм в структуру типа соленоида или упаковки нуклеосом в частицы, каждая из к-рых состоит из 6—10 нуклеосом, получившие название нуклеомеры, или супербиды. При этом коэффициент линейной упаковки ДНК равен примерно 40. Механизмы организации ДНП в структуры более высокого порядка недостаточно ясны. Эта организация специфична для каждого типа клеток, различающихся по характеру синтеза РНК. Происходящие на этом уровне процессы конденсации или деконденсации ДНП имеют существенное значение для функционирования генома, «включая» или «выключая» большие блоки генов, напр, при дифференцировке клеток, инактивации конденсированных участков хроматина в интерфазных ядрах, инактивации сегментов политенных хромосом и т. д.
Т. о., структурная динамика нуклеосом, определяющая возможность их полиморфизма, является, по-видимому, одним из механизмов регуляции функциональной активности генома и его изменений через рекомбинации. Изменения активности генома происходят также при нарушении в Н. соотношения гистон /ДНК. Напр., снижение количества или скорости синтеза гистонов, вызванное нехваткой генов, контролирующих синтез гистонов у дрозофилы, уменьшает величину соотношения гистон/ДНК в Н., повышает матричную активность Х-хромосомы и поддерживает активность гена, перенесенного в гетерохроматин. Возрастание величины отношения гистон/ДНК приводит к репрессии определенных генов. Изменения экспрессии (выражения) генов происходят и при модификации гистонов, осуществляющейся в процессе клеточного цикла. Так, ацетилирование гистонов коррелирует с активацией транскрипции генами, а их фосфорилирование — с репликацией и образованием митотических хромосом. В основном все типы модификации гистонов реализуются в ДНП через изменение их электростатических взаимодействий с ДНК.
Изменения взаимоотношения ДНК и белков в Н. имеют место при малигнизации, нек-рых хромосомных болезнях и др. Напр., при болезни Дауна в ряде участков ядер клеток ДНП более конденсированы, чем в норме. При этом локальные участки ДНК Н. попадают в иное микроокружение, что приводит к изменению констант их связывания с соответствующим субстратом, т. е. изменению экспрессии ряда генов, если последние вовлечены в эти участки.
В результате действия мутагенов физ. и хим. природы в ДНП возникают многочисленные повреждения: частичная диссоциация Н., образование «сшивок» между ДНК и нек-рыми белками Н., распад какого-то количества Н. на нуклеосомы, нарушение структурообразования нуклеопротеида. Вместе с тем белки, перехватывая свободные радикалы, частично защищают ДНК в Н., уменьшая деструкцию азотистых оснований и число одиночных и парных разрывов. Нек-рые из перечисленных повреждений имеют генетические, а часть — функциональные последствия.
Библиография: Георгиев Г. П. и Бакаев В. В. Три уровня структурной организации хромосом эукариот, Молек. биол., т. 12, в. 6, с. 1205, 1978, библиогр.; Нейфах А. А. и Тимофеева М. Я. Проблемы регуляции в молекулярной биологии развития, М., 1978, библиогр.; Спитковский Д. М. Полиморфизм надмолекулярной организации хроматина в клетках человека и его роль в наследственной патологии, в кн.: Теорет. пробл. мед. генетики, под ред. А. Ф. Захарова, с. 52, М., 1979; Хесин Р. Б. Непостоянство генома, Молек. биол., т. 14, в. 6, с. 1205, 1980; Mс Ghee J. D. a. Felsenf eld G. Nucleosome structure, Ann. Rev. Biochem., v. 49, p. 1115, 1980; Tike honenkoT. I. Structure of viral acids in situ, в кн.: Comprehensive virology, ed. by H. Fraenkel-Conrat a. R. R. Wagner, v. 5, p. 2, N. Y.—L., 1975.



