Доказательства эволюции
Пути эволюции
В своих работах советский ученый Северцов А.Н. выделил понятия биологического прогресса и регресса.
Ароморфоз представляет собой прогрессивное эволюционное преобразование, повышающее уровень организации организмов. В результате ароморфоза становится возможным освоение новых, ранее недоступных для жизни, территорий. К примеру, теплокровность птиц позволила им заселить места с холодным климатом.

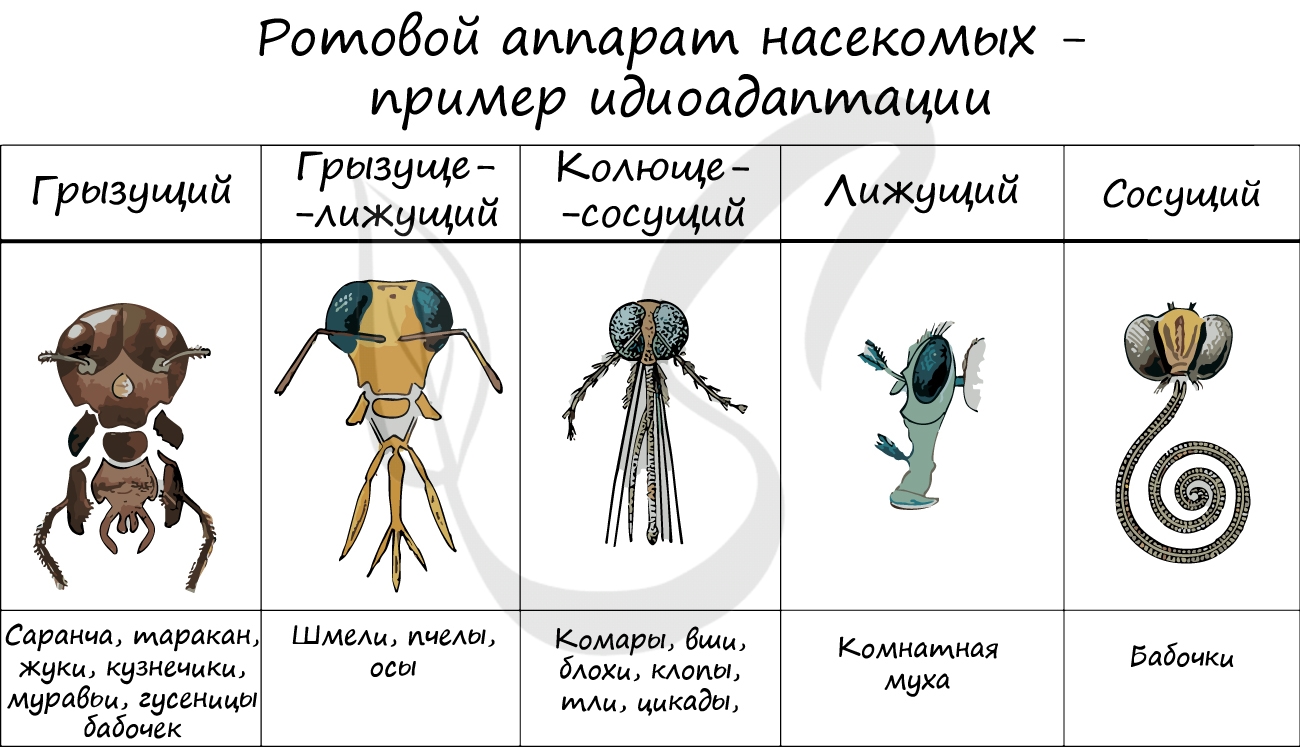
Идиоадаптация подразумевает незначительные, частные изменения в строении и функциях организма, которые помогают приспособиться к условиям среды обитания. Идиоадаптации существенно не повышают уровень организации.
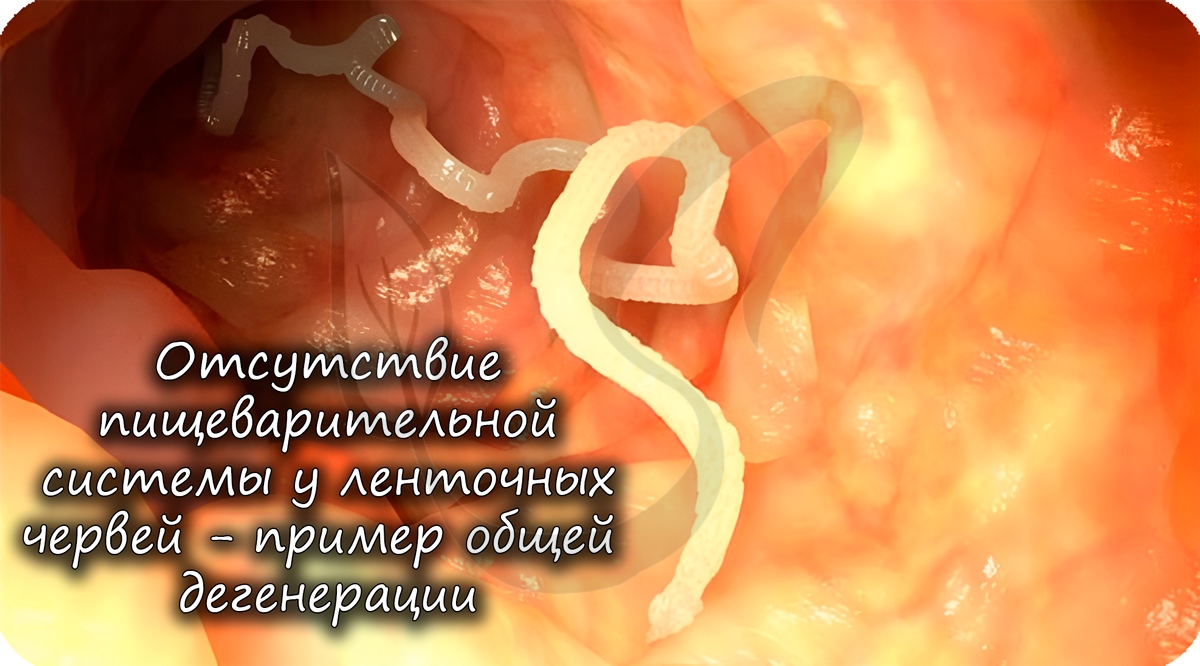
Общей дегенерацией называют упрощение организации, которое заключается в утрате отдельных органов и систем органов. У многих этот пункт вызывает внутреннее противоречие: как общая дегенерация может относиться к биологическому прогрессу?
У многих паразитов отсутствуют различные органы, к примеру, у ленточных червей нет пищеварительной системы. А зачем она им, когда пища в кишке, где они обитают, уже переварена и расщеплена организмом хозяина?
Биологический регресс характеризуется признаками, противоположными биологическому прогрессу:
Главная причина биологического регресса в том, что скорость эволюции вида отстает от скорости изменения внешней среды, эволюции других видов: это несоответствие снижает приспособленность организмов. Часто деятельность человека молниеносно меняет окружающую среду: далеко не все виды могут приспособиться к этому, происходит вымирание.

Сравнительно-анатомические доказательства эволюции
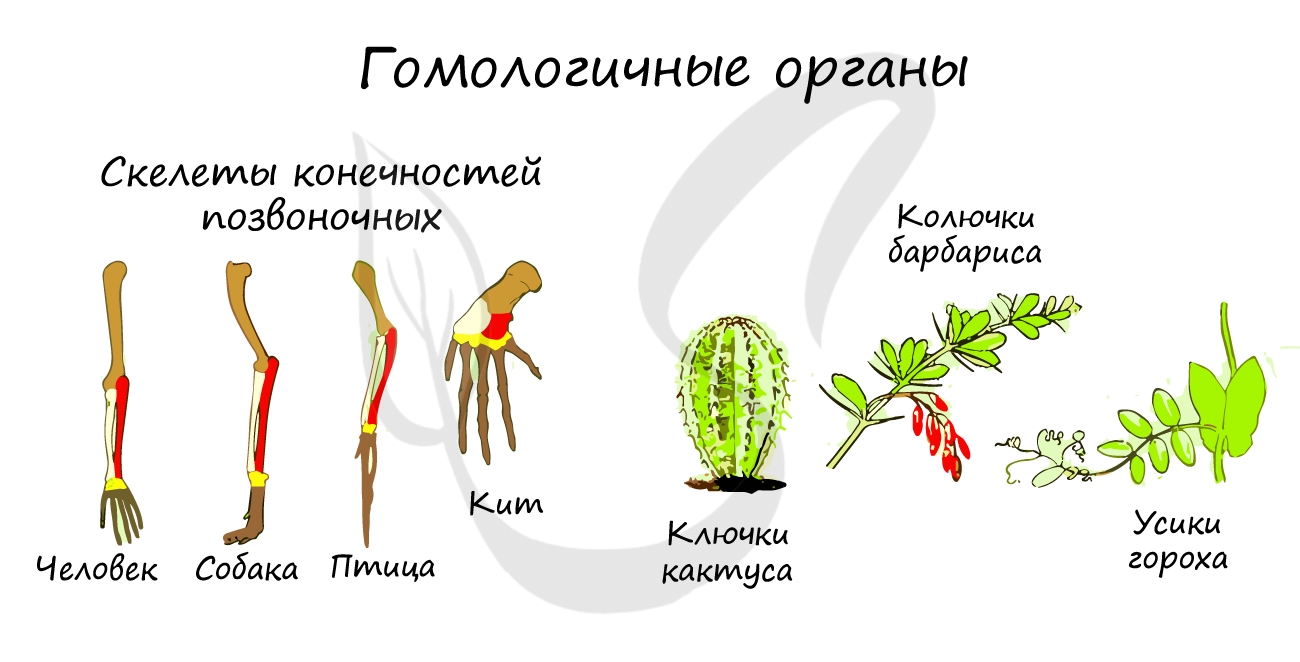
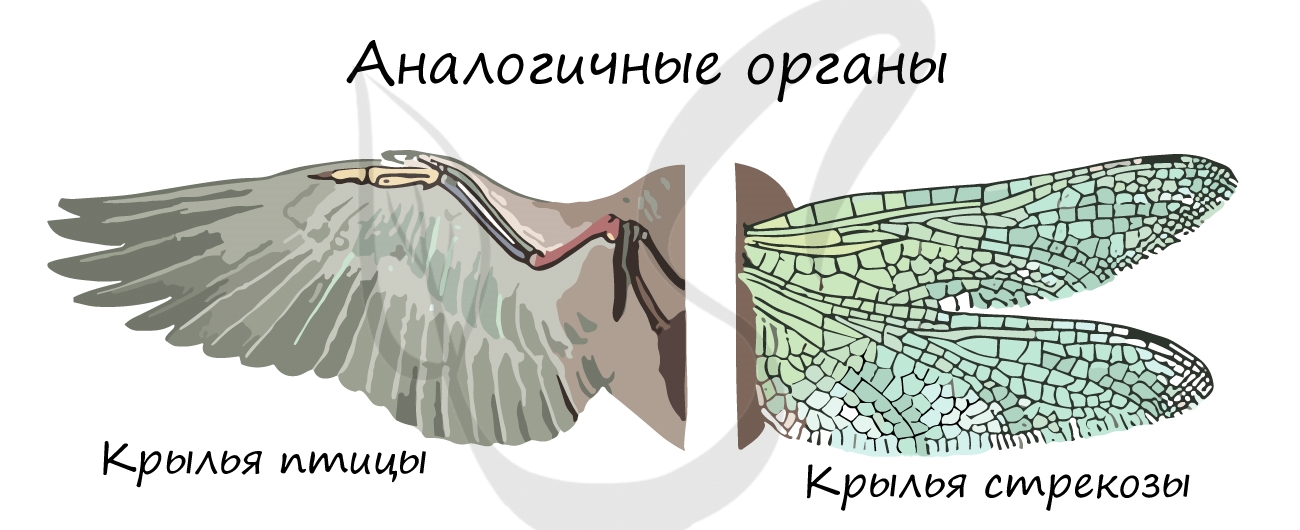
В строении нынешних животных можно найти признаки древних предковых форм, которые также свидетельствуют об эволюции. Сейчас мы обсудим рудименты и атавизмы.
У человека к рудиментарным органам относятся: зубы мудрости, копчик, ушные мышцы, аппендикс (червеобразный отросток), третье веко (эпикантус).
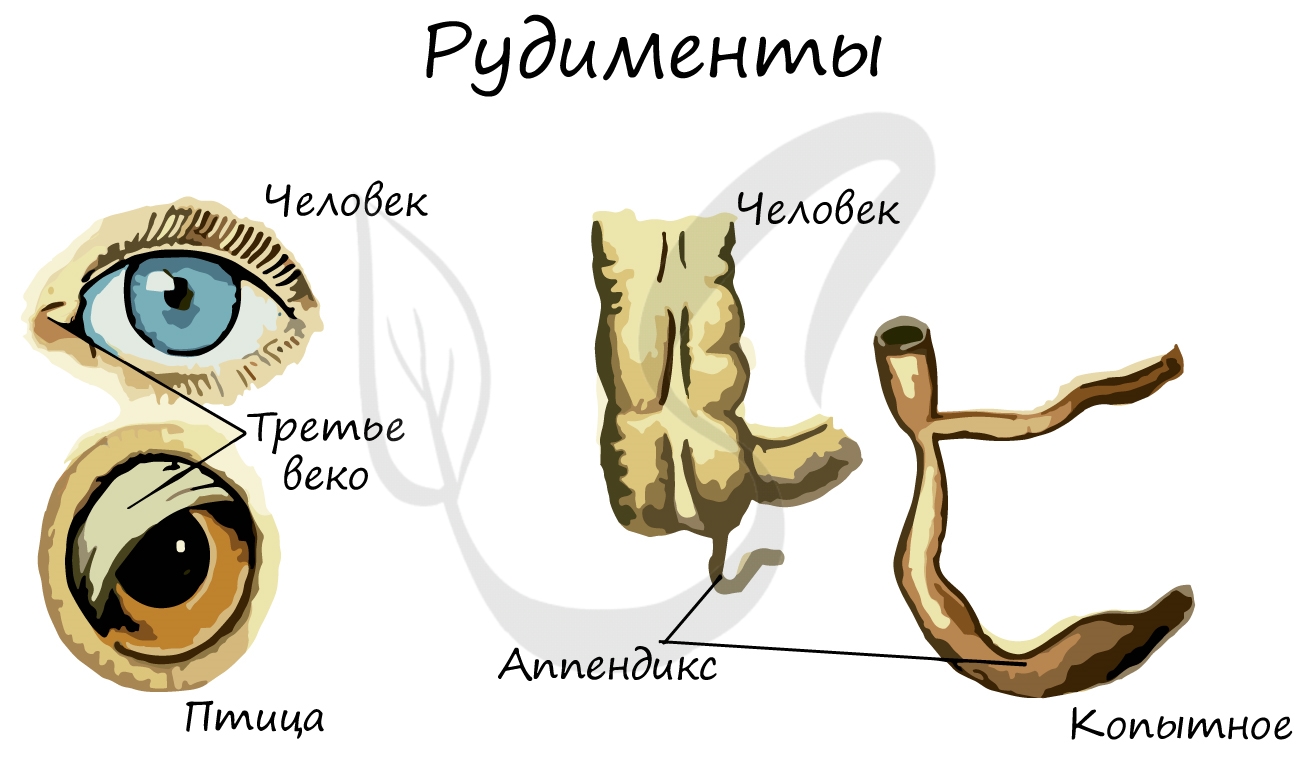
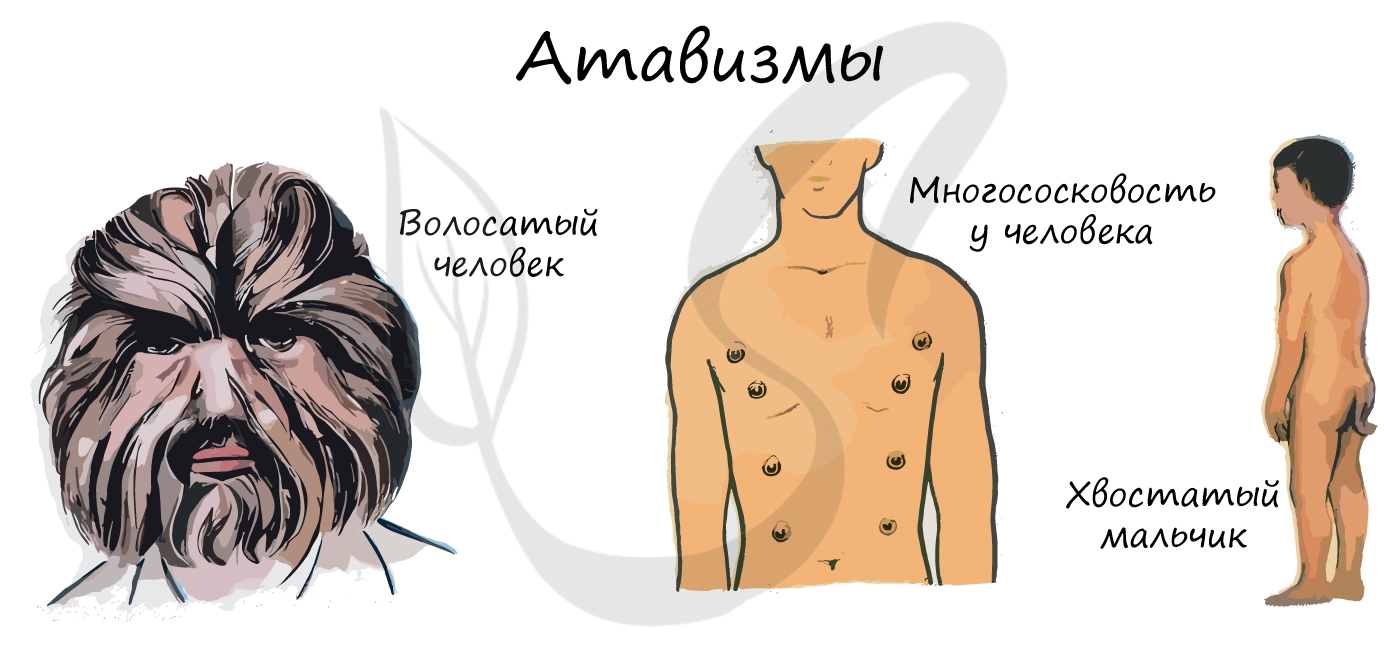
У человека атавизмами могут являться хвост, волосатое тело, добавочные молочные железы, незаращение межпредсердной перегородки.
Переходные формы
Такими формами являются, к примеру, утконос и ехидна из класса млекопитающих. При многих признаках млекопитающих, они откладывают яйца, тем самым подтверждают родство млекопитающих с пресмыкающимися.

Эмбриологические доказательства
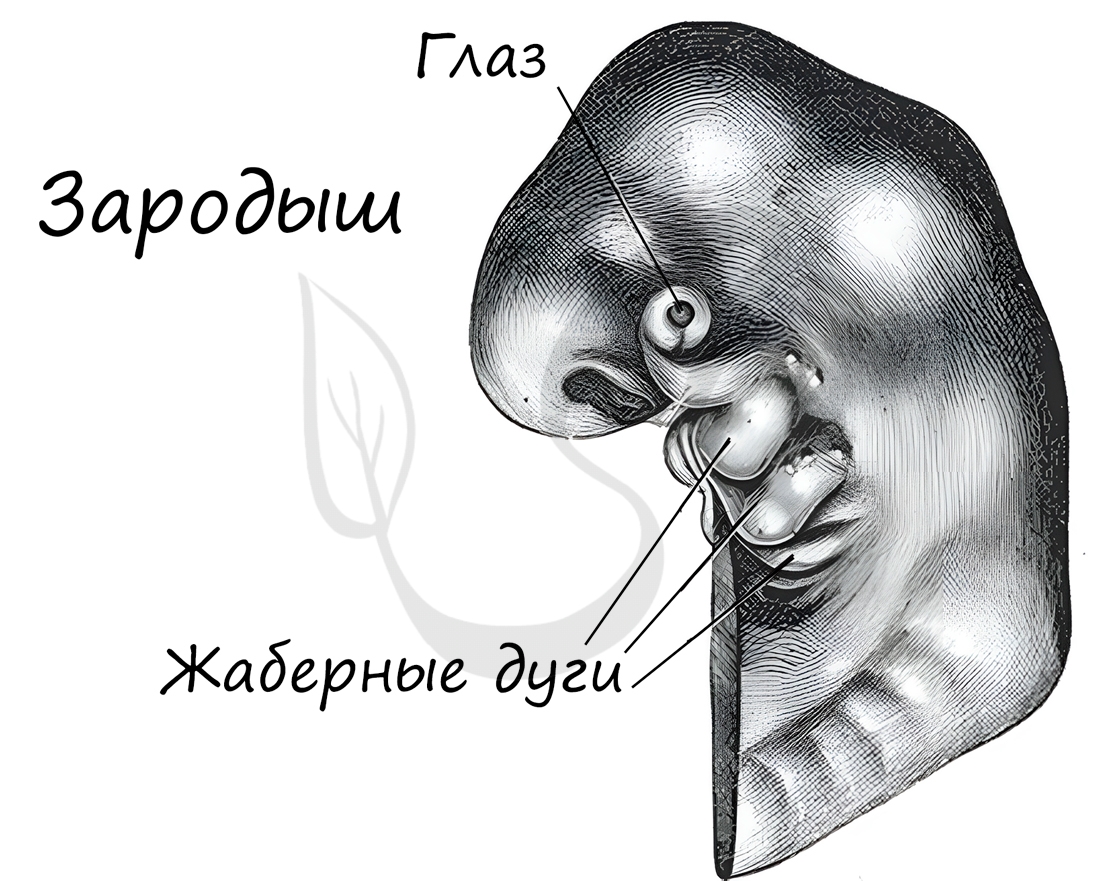
Немецкие ученые Ф. Мюллер и Э. Геккель во второй половине XIX века сформулировали биогенетический закон, гласящий, что онтогенез (индивидуальное развитие) каждой особи есть краткое и быстрое повторение филогенеза (исторического развития вида).
Биогенетический закон Мюллера-Геккеля объясняет повторение этапов (на стадии зародыша), которые были свойственны нашим далеким предкам. Таким образом, мы проходим их этапы, но, не останавливаясь на них, двигаемся дальше к более совершенным этапам.
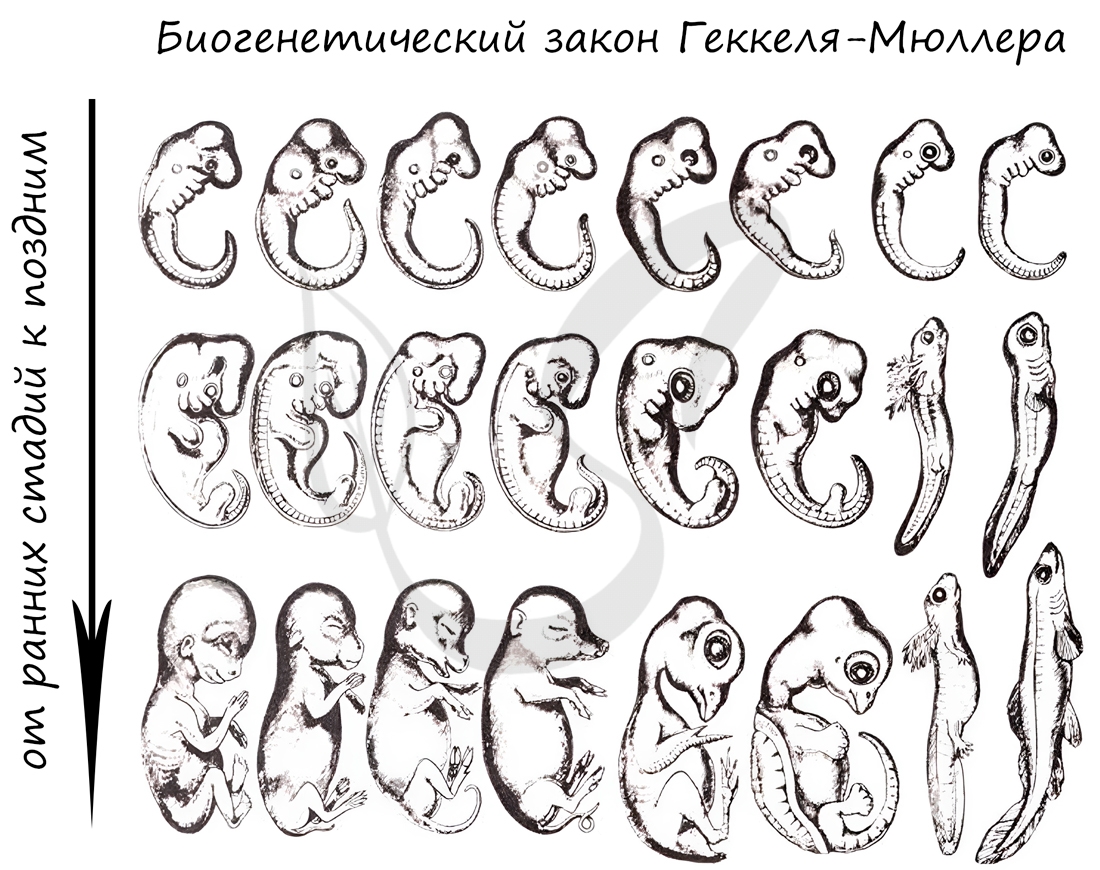
Карл Бэр сформулировал закон зародышевого сходства, который гласит, что на ранних стадиях развития зародыши позвоночных животных настолько похожи друг на друга, что практически неразличимы между собой. Это также указывает и подтверждает единство происхождения животного мира.
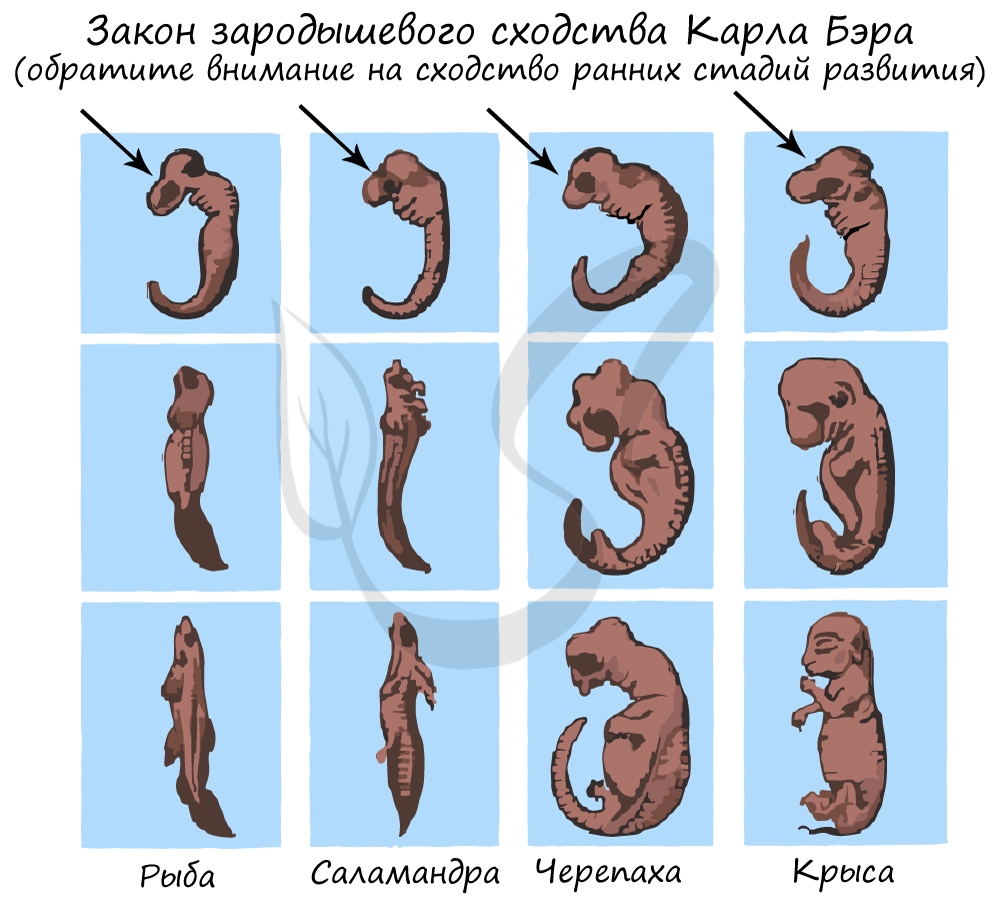
Палеонтологические доказательства эволюции
Палеонтология (греч. palaios – древний) изучает ископаемые останки вымерших животных, их сходства и различия с ныне живущими видами. Сопоставляя друг с другом ископаемые останки разных геологических эпох, можно увидеть как происходила эволюция различных видов животных и растений.
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Переходная форма


Переходная форма — организм с промежуточным состоянием, обязательно существующим при состоявшемся постепенном переходе от одного биологического типа строения к другому. Переходные формы характеризуются наличием более древних и примитивных (в значении первичных) черт, чем их более поздние родственники, но, в то же время, наличием более прогрессивных (в значении более поздних) черт, чем их предки. Как правило, говоря о промежуточных формах, имеют в виду ископаемые виды, хотя промежуточные виды вовсе не должны непременно вымирать. Известно множество переходных форм, иллюстрирующих происхождение четвероногих от рыб, рептилий от земноводных, птиц от динозавров, млекопитающих от териодонтов, китообразных от наземных млекопитающих, лошадей от пятипалого предка и человека от древних гоминид.
Содержание
Теория эволюции
Ко времени написания Чарльзом Дарвином «Происхождения видов» науке не были известны переходные формы жизни. Этот факт Дарвин считал одним из самых слабых мест своей теории. Однако уже через два года был обнаружен археоптерикс — переходное звено между рептилиями и птицами.
Переходные формы между двумя группами организмов не обязательно являются потомками одной группы и предками другой. По ископаемым останкам, как правило, невозможно достоверно установить, является ли один организм прямым предком другого. Кроме того, вероятность найти в палеонтологической летописи чьего-нибудь прямого предка очень мала, и по умолчанию среди биологов принято такую возможность вообще не рассматривать. Гораздо выше вероятность обнаружить относительно близких родственников этого предка, схожих с ним по строению. Поэтому любая переходная форма автоматически интерпретируется как боковая ветвь эволюции, а не «участок ствола». [1]
Примеры

В качестве примеров переходных форм можно привести Амбулоцетуса — «ходячего кита» (переходная форма между китообразными и наземными млекопитающими), Тиктаалика и Ихтиостегу (переходная форма от рыб к земноводным), Мезогиппуса (между ранними лошадиными и современной лошадью).
Переходные формы в эволюции Человека Разумного

Спор о постепенности эволюционных преобразований
Переходные формы являются объектом исследований в споре между градуалистами и пунктуалистами. Градуалисты считают, что эволюция является постепенным, непрерывным процессом. Пунктуалисты, или сторонники теории прерывистого равновесия считают, что виды в течение долгого времени остаются неизменными, а изменения происходят в очень короткий промежуток времени. Переходные формы являются аргументом обеих сторон. Пунктуалисты утверждают, что большинство видов появляются внезапно и на протяжении всей своей истории меняются слабо. Градуалисты отмечают, что среди позвоночных нет ни одного бесспорного примера очень быстрого перехода от одного вида к другому, большинство переходов вообще плохо задокументированы ископаемыми. Также много хорошо описанных внутривидовых изменений. Например, описаны изменения среднекайнозойских кроликов, миоценовых грызунов, гиппарионов, эоценовых парнокопытных, переход между родами эоценовых приматов. [2]
Недостающие звенья
| 1850 |  |
| 1900 |  |
| 1950 | 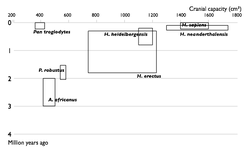 |
| 2002 | 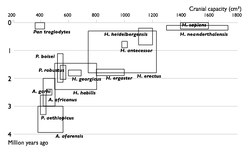 |
| Прогресс в изучении эволюции человека. Увеличение числа известных науке видов гоминин по времени. Каждый вид изображен в виде прямоугольника, показывающего границы, в которых варьировал объём черепа, и место вида в палеонтологической летописи. Видно постепенное заполнение промежутков между видами. | |
Многие переходные формы между живыми организмами пока не найдены. Обычно их называют недостающими звеньями. Редкость переходных форм объясняется неполнотой палеонтологической летописи. Сторонники теории прерывистого равновесия также указывают на быстроту эволюционных преобразований. Однако палеонтологи постоянно находят новые ископаемые, заполняя пробелы (например, в 2004 году был найден Тиктаалик — ещё одно переходное звено между рыбами и земноводными)
Неполнота палеонтологической летописи
Далеко не каждая переходная форма существует в виде окаменелости из-за принципиальной неполноты палеонтологической летописи. Неполнота вызвана особенностями процесса фоссилизации, то есть перехода в окаменелое состояние. Для образования окаменелости необходимо, чтобы погибший организм был погребён под большим слоем осадочных пород. Из-за крайне медленной скорости осадконакопления на суше, чисто сухопутные виды редко переходят в окаменелое состояние и сохраняются. Кроме того, редко удаётся обнаружить виды, живущие в глубинах океана, из-за редкости поднятия на поверхность больших массивов дна. Таким образом, большинство известных ископаемых (а, соответственно, и переходных форм) — это либо виды, обитающие на мелководье, в морях и реках, либо наземные виды, ведущие полуводный образ жизни, либо обитающие недалеко от береговой линии. Закономерности процессов захоронения останков живых существ изучает особая отрасль палеонтологии — тафономия.
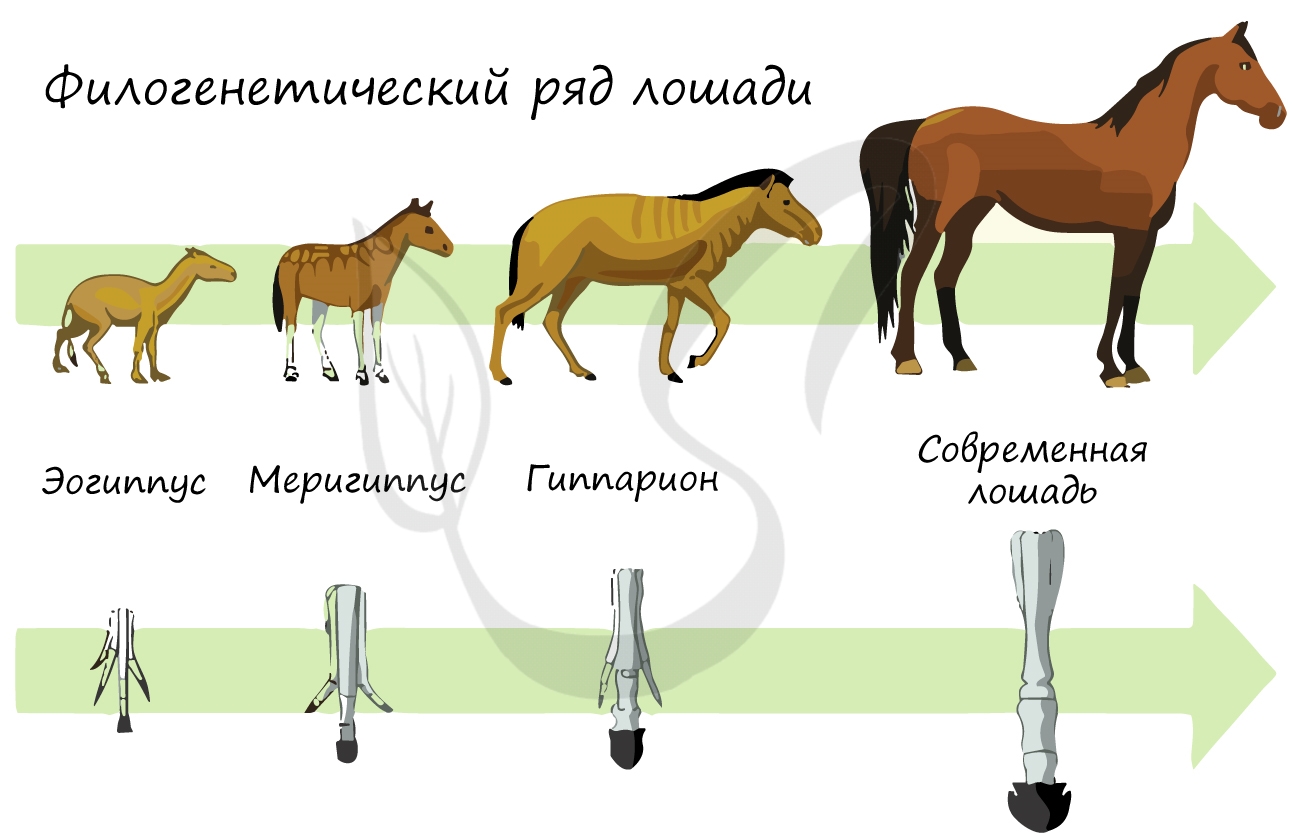
Филогенетические ряды

Филогенетические ряды − ряды видов, последовательно сменявших друг друга в процессе эволюции различных групп животных и растений.
Впервые были открыты В. О. Ковалевским, который показал, что современные однопалые копытные происходят от древних пятипалых мелких всеядных животных.
Креационизм
Сторонники креационизма заявляют, что переходных форм не обнаружено. Научное сообщество считает подобные заявления ложью и намеренным введением в заблуждение.
См. также
Примечания
Литература
Ссылки

Полезное
Смотреть что такое «Переходная форма» в других словарях:
Форма государственного устройства — способ территориальной организации государства или государств, образующих союз. Определяет внутреннее строение государства, деление его на составные части (территории) и принципы их взаимоотношения между собой. Для характеристики территориальной… … Википедия
переходная антиква — Переходная антиква, Антиква переходного стиля (Transitional Serif) Наз. также антиква барокко. Шрифты начала и середины XVIII в, в которых сочетаются черты антиквы старого стиля [один из видов шрифтов с засечками], обусловленные процессом… … Шрифтовая терминология
ФОРМА ПЕРЕХОДНАЯ — звено филогенетического ряда, имеющее самостоятельное таксономическое значение, связующее во времени предшествующие (предковые) и последующие формы. Геологический словарь: в 2 х томах. М.: Недра. Под редакцией К. Н. Паффенгольца и др.. 1978 … Геологическая энциклопедия
Сонатная форма — При возникновении С. формы, до XVII ст., каждая пьеса, предназначавшаяся для исполнения на инструменте, называлась сонатой (от sonare или suonare звучать), а для пения кантатой (cantare петь). Вокальные пьесы, переложенные на орган или клавикорды … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
прецистная форма — (пре + циста) переходная стадия развития простейших от трофозоита к цисте, лишенная оболочки … Большой медицинский словарь
Прибиржевой рынок ценных бумаг — – переходная форма рынка ценных бумаг, рынки, создаваемые фондовыми биржами и находящиеся под их регулирующим воздействием … Рынок ценных бумаг. Словарь основных терминов и понятий
Evolution: The Game of Intelligent Life — Разработчик Crossover Technologies, Discovery Channel Multimedia Издатель СА Interplay Entertainment … Википедия
Государство — (Country) Государство это особая организация общества, обеспечивающая единство и целостность, гарантирующая права и свободы граждан Происхождение государства, признаки государства, форма государственного правления, форма государственного… … Энциклопедия инвестора
Шлем — «Святой Георгий». Гравюра Л. Кранаха. 1506 г. На гравюре представлен полный рыцарский доспех начала XVI в. с различными съемными усилениями. Доспех максимилиановского типа без гофрировки. Любопытно, что на нем отсутствует ожерелье, функция… … Энциклопедия средневекового оружия
Поземельная община — Содержание: I. П. община в Западной. Европе. II. П. община в Византии. III. П. община во внеевропейских странах. IV. П. община в Древней Руси и в Великороссии. V. П. община в Малороссии и в Литве. VI. П. община (современное положение; вопрос о П … Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона
Переходная форма
Переходные (промежуточные) формы — организмы, которые сочетают в своем строении признаки двух больших систематических групп.
Переходные формы характеризуются наличием более древних и примитивных (в смысле первичных) черт, чем более поздние формы, но, в то же время, наличием более прогрессивных (в смысле более поздних) черт, чем их предки. Как правило, термин «переходные формы» употребляют по отношению к ископаемых форм, хотя промежуточные виды совсем не обязательно должны умирать.
Переходные формы используют как одно из доказательств существования биологической эволюции.
История понятия
В 1859 г. Когда была издана работа Ч. Дарвина «Происхождение видов», количество ископаемых остатков была крайне малой, науке не были известны переходные формы. Дарвин описал отсутствие промежуточных форм «как наиболее очевидное и тяжелое возражение, которое может быть против теории», но объяснил это крайней неполнотой геологической летописи. Он отмечал ограниченное количество доступных коллекций в то время, в то же время описал имеющуюся информацию об имеющихся ископаемые образцы с точки зрения эволюции и действия естественного отбора. Только два года спустя, в 1961 году. Был найден археоптерикс, который представлял классическую переходную форму между пресмыкающимися и птицами. Его находках, стала не только подтверждением теории Дарвина, а также знаковым фактом, подтверждающий реальность существования биологической эволюции. С тех пор было найдено большое количество ископаемых форм, которые показывают, что все классы позвоночных животных являются родственными между собой, причем большинство из них — через переходные формы.
С увеличением сведений о таксономическое разнообразие сосудистых растений в начале ХХ в., Начались исследования по поиску их возможного предка. В 1917 г.. Роберт Кидстон и Уильям Генри Ленд обнаружили остатки очень примитивной растения возле поселка Rhynia в Шотландии. Это растение было названо Rhynia. Она сочетает в себе признаки зеленых водорослей и сосудистых растений.
Трактовка понятия
Переходные формы между двумя группами организмов не обязательно потомками одной группы и предком другой. По ископаемыми, как правило, невозможно точно установить является ли определенный организм предком другого. Кроме того, вероятность найти в палеонтологической летописи прямого предка определенной формы чрезвычайно мала. Гораздо больше вероятность обнаружить относительно близких родственников этого предка, которые сходны с ним по строению. Поэтому любая переходная форма автоматически интерпретируется как боковая ветвь эволюции, а не «участок филогенетического ствола».
Переходные формы и таксономия
Эволюционная таксономия оставалась доминирующей формой таксономии течение ХХ в. Выделение таксонов базируется на различных признаках, вследствие чего таксоны изображают в виде ветвей разветвленного эволюционного дерева. Переходные формы рассматриваются как «падающие» между различными группами в плане анатомии, они смесь характеристик от внутренней и внешней клади, что недавно разделилась.
С развитием кладистики в 1990-х гг. Взаимосвязи обычно изображают в виде кладограмы, иллюстрирующую дихотомическое ветвление эволюционных линий. Поэтому в кладистици переходные формы рассматриваются как более ранние ветви дерева, где еще не развились не все черты, характерные для ранее известных потомков на этой ветке. Такие ранние представители группы обычно называют основным таксоном (англ. Basal taxa) или сестринским таксоном (англ. Sister taxa), в зависимости от того, принадлежит ли ископаемый организм к данной клади или нет.
Проблемы выявления и интерпретации
Отсутствие переходных форм между многими группами организмов является предметом критики со стороны креационистов. Однако далеко не каждая переходная форма существует в виде окаменелостей из-за принципиальной неполноты палеонтологической летописи. Неполнота вызвана особенностями процесса фосилизации, то есть перехода в окаменевший состояние. Для образования окаменелости необходимо, чтобы организм, который погиб, был погребен под большим слоем осадочных пород. Из-за очень медленную скорость осадконакопления на суше, сухопутные виды редко переходят в окаменевший состояние и сохраняются. Кроме того, редко удается выявить виды, которые живут в глубинах океана через редкие случаи поднятия на поверхность больших массивов дна. Таким образом, большинство известных ископаемых (а, соответственно, и переходных форм) — это либо виды, обитающие на мелководье, в морях и реках, или наземные виды, которые ведут полуводный образ жизни, или живут у береговой линии. К упомянутым выше проблемам следует добавить чрезвычайно малую (в масштабах планеты) количество палеонтологов, которые осуществляют раскопки.
Переходные формы, как правило, не живут на больших территориях и не существуют в течение большого времени, иначе они были бы персистентный. Этот факт также снижает вероятность фосилизации и последующего обнаружения переходных форм.
Поэтому вероятность обнаружения промежуточных форм чрезвычайно мала.
Примеры среди животных
Древнейшими представителями земноводных считают ихтиостеги. Их считают переходным звеном между кистеперых рыбами и земноводными. Несмотря на то, что в ихтиостеги была пятипалая концовка, адаптированная к жизни на суше, значительную часть жизни они проводили как рыбы, имели хвостовой плавник, боковую линию и некоторые другие признаки рыб.
Батрахозавры, существовавшие в каменноугольный и пермский периоды, рассматривают как переходную форму между земноводными и пресмыкающимися. Батрахозавры, хоть и проводили жизнь во взрослой стадии на суше (подобно пресмыкающихся), были тесно связаны с водоемами и сохранили ряд признаков, присущих земноводным, в частности, откладывания икры и развитие личинок в воде, наличие жабр и тому подобное.
Обнаружено большое количество пресмыкающихся, которые выработали способность летать, часть из них имела перья, поэтому их рассматривают как переходные формы между пресмыкающимися и птицами. Наиболее известен археоптерикс. Он был размером примерно с современную ворону. Формой тела, строением конечностей и наличием оперения подобный современных птиц, возможно, летал. Общим с пресмыкающимися была особое строение таза и ребер, наличие клюва с коническими зубами, по три свободные пальцы на крыльях, подвийноувигнути позвонки, длинный хвост с 20-21 позвонка, кости могло не пневматизовани, грудная кость без киля. Другие известные переходные формы между пресмыкающимися и птицами — протоавис, конфуциусорниса.
Одной из форм, сохранилась в ископаемом состоянии является амбулоцетус Ambulocetus natans («ходячий кит») — переходная форма между наземными млекопитающими и китообразными, которые являются вторинноводнимы формами. Внешне животное напоминало нечто среднее между крокодилом и дельфином. Кожа должна частично редуцированную шерсть. Животное имело лапы с перепонками; хвост и конечности приспособлены как вспомогательные органы передвижения в воде.
Примеры среди растений
Первые наземные растения из класса риниопсид, семей риниевих и псилофитовых, живших в силуре — девоне, сочетали признаки зеленых водорослей и примитивных форм высших растений. Их тело было безлистным, цилиндрический осевой орган — телом в верхней части дихотомически разветвленным на верхушках с спорангиями. Функцию минерального питания риниопсид выполняли ризоиды.
Ископаемые формы семенных папоротников, которые процветали в конце девона, сочетают в себе признаки папоротников и голосеменных. Они образовывали не только споры (как папоротники), но и семена (как семенах растения). Проводящая ткань их стеблей по строению напоминает древесину голосеменных (саговников).
Другой предшественник семенных растений был идентифицирован из отложений среднего девона. Рункария (Runcaria heinzelinii) существовала около 20 млн лет назад. Это была небольшая растение с радиальной симметрией; имела спорангий, окруженный интегументом и плюской. Рункария демонстрирует путь эволюции растений от споровых к семенных.
Переходные формы в эволюции человека
В наше время найдено большое количество ископаемых останков, которые раскрывают эволюционный путь человека разумного от ее человекообразных предков. К формам, которые в большей или меньшей степени можно отнести к переходным, относятся: сахелантропа, ардипитека, австралопитеки (африканский, афарский и другие), человек умелый, человек работающий, человек прямоходящий, человек-предшественник, гейдельбергский человек и кроманьонцы.
Среди упомянутых форм значительное внимание заслуживают австралопитеки. Австралопитек афарский с точки зрения эволюции находится между современными двуногими людьми и их четвероногими древними предками. Большое количество рис скелета этого австралопитека четко отражают двуногость, причем до такой степени, что некоторые исследователи считают, что это свойство возникла задолго до появления австралопитека афарского. Среди общих черт анатомии, его таз гораздо больше похож на этих костей у человека, чем у обезьян. Края подвздошных костей короче и шире, крестцовая кость широкая и расположена непосредственно позади тазобедренного сустава. Существует явное свидетельство о существовании мест крепления для мышц-разгибателей колена, предусматривает вертикальное положение этого организма. В то время, как таз австралопитека не совсем как у человека (заметно шире, с ориентацией края подвздошных костей наружу), эти особенности указывают на принципиальную перестройку, связанную с хождением на двух ногах. Бедренная кость образует угол в направлении колена. Эта черта позволяет ноге размещаться ближе к средней линии тела и является явным свидетельством привычный характер передвижения на двух ногах. В наше время человек разумный, орангутаны и коаты имеют такие же черты. Ноги австралопитека имели большие пальцы, что делает практически невозможным захват стопой ветвей деревьев. Кроме особенностей локомоции, в австралопитека был также значительно больше мозг, чем у современных шимпанзе и зубы были значительно больше подобными зубов современного человека, чем к обезьянам.
Филогенетические ряды
Филогенетические ряды — ряды ископаемых форм, связанные между собой в процессе эволюции и отражают постепенные изменения их исторического развития.
Были исследованы русским ученым А. Ковалевским и английским Дж. Симпсоном. Они показали, что современные однопалого копытные происходят от древних мелких всеядных животных. Анализ ископаемых лошадей помог установить постепенность процесса эволюции в пределах этой группы животных, в частности, как изменяясь во времени, ископаемые формы приобретали все большего сходства с современными лошадьми. Сравнивая эоценового еогипуса с современным конем, трудно доказать их филогенетическую родство. Однако наличие ряда переходных форм, которые последовательно сменяли друг друга на больших пространствах Евразии и Северной Америки, позволила восстановить филогенетический ряд лошадей и установить направление их эволюционных изменений. Он состоит из ряда следующих форм (в упрощенном виде): Phenacodus — Eohippus — Miohippus — Parahippus — Pliohippus — Equus.
Гильгендорф (1866) описал палеонтологический ряд брюхоногих моллюсков из миоценовых отложений, накопившихся в течение двух миллионов лет в озерных отложениях Штейнгеймського бассейна (Вюртемберг, Германия). Было обнаружено в последовательных слоях 29 различных форм, принадлежащих к ряду планорбис (Planorbis). Древние моллюски имели раковину в виде спираи, а более поздние — в виде турбоспирали. Ряд имел два ответвление. Предполагается, что изменение формы черепашки была вызвана повышением температуры и увеличением содержания карбоната кальция в результате горячих вулканических источников.
Таким образом, филогенетические ряды представляют собой историческую последовательность переходных форм.
В настоящее время известны филогенетические ряды для аммонитов (Вааген, 1869), брюхоногих моллюсков рода живородок (Viviparus) (Неймайром, 1875), носорогов, слонов, верблюдов, парнокопытных и других животных.



