Глава 9. Экосистемы
9.4. Динамика экосистем
9.4.2. Сукцессии и дигрессии
Поступательные изменения в сообществе приводят в конечном счете к смене этого сообщества другим, с иным набором господствующих видов. Причиной подобных смен могут быть внешние по отношению к ценозу факторы, длительное время действующие в одном направлении, например возрастающее в результате мелиорации иссушение болотных почв, увеличивающееся загрязнение водоемов, усиленный выпас скота, вытаптывание лесопарков населением городов и т. п. Возникающие при этом смены одного биоценоза другим называют экзогенетическими. Если при этом усиливающееся влияние фактора приводит к постепенному упрощению структуры сообществ, обеднению их состава, снижению продуктивности, то подобные смены называют дигрессионными или дигрессиями.
Эндогенетические смены возникают в результате процессов, происходящих внутри самого сообщества. Закономерный направленный процесс изменения сообществ в результате взаимодействия живых организмов между собой и окружающей их абиотической средой называют сукцессией.
Причины возникновения сукцессии. Сукцессия (от лат. successio– преемственность, наследование) – это процесс саморазвития сообществ. В основе сукцессии лежит неполнота биологического круговорота в данном ценозе. Каждый живой организм в результате жизнедеятельности меняет вокруг себя среду, изымая из нее часть веществ и насыщая ее продуктами метаболизма. При более или менее длительном существовании популяций они меняют свое окружение в неблагоприятную сторону и в результате оказываются вытесненными популяциями других видов, для которых вызванные преобразования среды оказываются экологически выгодными. Таким образом, в сообществе происходит смена господствующих видов. Длительное существование биоценоза возможно лишь в том случае, если изменения среды, вызванные деятельностью одних организмов, точно компенсируются деятельностью других, с противоположными экологическими требованиями.
В ходе сукцессии на основе конкурентных взаимодействий видов происходит постепенное формирование более устойчивых комбинаций, соответствующих конкретным абиотическим условиям среды. Примерами сукцессий могут быть смены видов при зарастании стоячих водоемов (рис. 153) или барханных песков в пустыне.
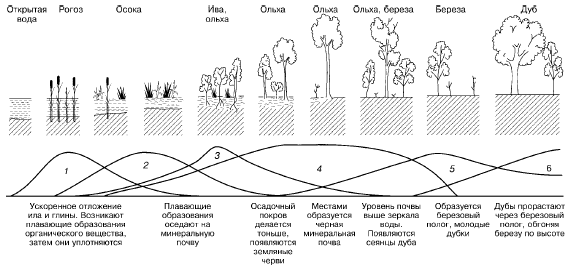
Рис. 153. Изменение экосистемы в ходе сукцессии при зарастании озера (по Д. Казенс, 1982): 1–6– смена доминирующих видов растений
Сыпучие барханные пески Каракумов и других районов Средней Азии сначала полностью лишены растительности и постоянного населения животных. Первым на них поселяется многолетний злак аристида, хорошо приспособленный к жизни в условиях постоянного переноса песка ветром. Корни у этого растения шнуровидные, и каждый заключен в чехол из сцементированных песчинок, что защищает корни от высыхания и механического повреждения, если они окажутся на поверхности. Побеги растут вертикально вверх и образуют дополнительные зоны кущения выше материнского, если песок засыпает растение. За счет аристиды уже могут существовать некоторые насекомые, и поэтому на барханы начинают забегать в поисках пищи ящерицы рода Eremias. На слегка скрепленных корнями аристиды песках получает возможность поселиться длиннокорневищная песчаная осока. Она успешно борется с песком, быстро прорастая сквозь его наносы и пронизывая песок ветвящимися корневищами на глубине 1–5 см. Покров разрежен, так как для обеспечения влагой одного растения нужна значительная площадь. На скрепленных злаками и осокой песках поселяются затем кустарники джузгун и белый саксаул, а также другие травянистые растения, в основном эфемеры: злаки, крестоцветные, мотыльковые, бурачниковые и т. п. Вслед за растительностью появляются растительноядные млекопитающие: тонкопалый суслик, мохноногий тушканчик, полуденная песчанка. Увеличивается видовое разнообразие насекомых – кормовой базы ящериц: ушастой и песчаной круглоголовок, сетчатой ящурки, гекконов. Появляются птицы – саксаульная сойка, дрофа-красотка, затем змея и хищные млекопитающие. Закрепленные пески Средней Азии отличаются большим видовым богатством и разнообразием жизненных форм растений и животных, так как водный режим их достаточно благоприятен: пески обладают способностью сгущать водяные пары в почве во влагу.
Последовательный ряд постепенно и закономерно сменяющих друг друга в сукцессии сообществ называется сукцессионной серией.
Сукцессии в природе чрезвычайно разномасштабны. Их можно наблюдать в пробирках с сенным настоем, где сменяют друг друга разные виды микроорганизмов и простейших (рис. 154), в лужах и прудах, в одной и той же порции листового опада в лесу, где происходят закономерные смены сапрофитов и сапрофагов, на стволах отмирающих деревьев, в пнях, на кротовинах, выбросах из нор сурков, зарастающих отмелях, выветривающихся скалах, на заброшенных пашнях, насыпях, на болотах, лугах, в лесах и т. д. Иерархичность в организации сообществ проявляется и в иерархичности сукцессионных процессов: более крупные преобразования биоценозов складываются из более мелких. Даже в стабильных экосистемах с хорошо отрегулированным круговоротом веществ постоянно осуществляется множество локальных сукцессионных смен, поддерживающих сложную внутреннюю структуру сообществ.
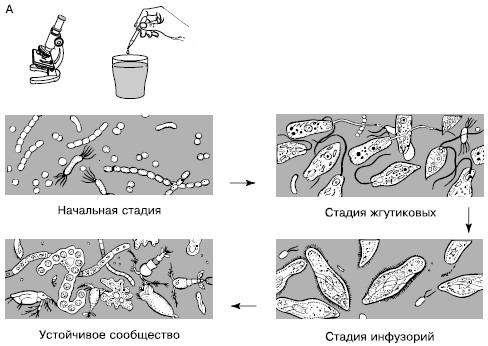
Рис. 154. Смена простейших в сенном настое (по Р. Дажо, 1975):
Б – конкретная серия:
1 – жгутиковые; 2 – инфузории кольноды; 3 – нарамеции; 4 – брюхо-ресничные инфузории; 5 – амебы; 6 – сувойки
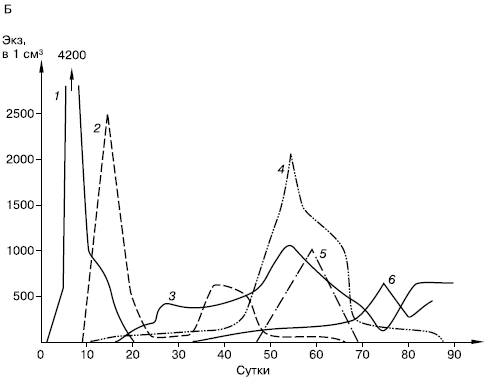
Типы сукцессионных смен. Выделяют два основных типа сукцессионных смен: 1) с участием как автотрофного, так и гетеротрофного населения и 2) с участием лишь гетеротрофов. Сукцессии второго типа совершаются только в таких условиях, где создается предварительный запас или постоянное поступление органических соединений, за счет которых существует сообщество: в сильно загрязненных органическими веществами водоемах, в скоплениях разлагающейся растительной массы, в кучках или буртах навоза, компостах, в пещерах с гуано летучих мышей и т. п. (рис. 155).
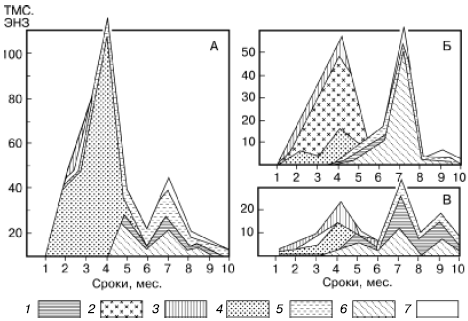
Рис. 155. Изменение численности разных групп микроартропод в ходе сукцессии при разложении растительных остатков клевера (А), картофельной ботвы (Б), ячменя (В) в пересчете на 100 г сухого органического вещества (по Н. М. Черновой, 1977): 1 – панцирные клещи; 2 – клещи-аноетиды; 3 – гипопусы аноетид; 4 – тироглифоидные клещи; 5 – их гипопусы; 6 – коллемболы; 7 – прочие группы
Сукцессии со сменой растительности могут быть первичными и вторичными.
Первичные сукцессии начинаются на лишенных жизни местах – на скалах, обрывах, наносах рек, сыпучих песках и т. п. При заселении таких участков живые организмы необратимо меняют местообитание и сменяют друг друга. Основная роль принадлежит накоплению отмерших растительных остатков или продуктов разложения, что зависит как от характера растительности, так и от комплекса разрушителей мертвой растительной массы – животных, грибов и микроорганизмов. Постепенно формируется почвенный профиль, изменяется гидрологический режим участка, его микроклимат. Такие сукцессии в геоботанике называют экогенетическими, так как они ведут к преобразованию самого местообитания.
В первичных сукцессиях, протекающих на скалах в лесах Урала, различают четыре этапа.
1. Поселение эндолитических и накипных лишайников, которые сплошь покрывают каменистую поверхность. Накипные лишайники несут своеобразную микрофлору и содержат богатую фауну простейших, коловраток и нематод. Мелкие клещи-сапрофаги и первичнобескрылые насекомые обнаруживаются сначала только в трещинах. Активность всего населения прерывиста, в основном после смачивания скал дождем или туманом.
2. Преобладание листоватых лишайников, постепенно образующих сплошной ковер. Под круговинками лишайников Parmelia в результате действия выделяемых ими кислот и механического сокращения слоевищ при высыхании образуются выщербленности, идет отмирание слоевищ и накопление детрита. Под лишайниками в большом количестве обитают мелкие членистоногие: коллемболы, панцирные клещи, сеноеды, личинки комаров-толкунчиков и другие, образуется микрогоризонт, состоящий из их экскрементов.
3. Поселения литофильных мхов Hedwigia ciliata и Pleurozium schreberi. Они погребают лишайники и подлишайниковые пленочные почвы. Ризоиды мхов прикрепляются не к камню, а к мелкозему, мощность которого достигает уже 3 см. Под мхами колебания температуры и влажности уже в несколько раз меньше, чем под лишайниками. Усиливается деятельность микроорганизмов, и увеличивается разнообразие групп животных.
4. Появление гипновых мхов и сосудистых растений. В разложении растительных остатков и формировании почвенного профиля постепенно уменьшается роль мелких членистоногих и растет участие более крупных беспозвоночных-сапрофагов: энхитреид, дождевых червей, личинок насекомых.
Вторичные сукцессии представляют собой восстановительные смены. Они начинаются в том случае, если в уже сложившихся сообществах частично нарушены установившиеся взаимосвязи организмов, например удалена растительность одного или нескольких ярусов (в результате вырубки, пожара, вспашки и т. д.). Смены, ведущие к восстановлению прежнего состава ценоза, получили в геоботанике название демутационных. Примером может служить демутация залежей в Абаканских степях, происходящая в четыре основных этапа: 1) преобладание однолетних сорняков – на 1-2-й год после вспашки; 2) господство «крупного бурьяна», преимущественно разных полыней – на 3-4-й год; 3) вытеснение их корневищными злаками (пыреем и др.) и появление большого количества бобовых – начиная с 5-го года; 4) установление господства степных дерновинных злаков с появлением ковыля – на 11-12-й год после вспашки.
Восстановительные смены совершаются быстрее и легче, чем экогенетические, так как в нарушенном местообитании сохраняются почвенный профиль, семена, зачатки и часть прежнего населения и прежних связей. Демутации не являются повторением какого-либо этапа первичных сукцессии.
Частными вариантами сукцессии являются смены, которые по продолжительности точно совпадают с каким-либо сезоном, а на следующий год начинаются сначала. Эти «сезонные» сукцессии выглядят как обычная фенологическая смена форм, но природа их иная. Например, в планктоне северных морей весной массовое размножение водорослей сменяется волной размножения рачков-калянусов, затем в большом количестве появляются гребневики, а после них – некоторые виды рыб. Эти смены происходят как постепенное развертывание в течение короткого северного лета единой гигантской трофической цепи организмов, так как каждая последующая группа питается за счет предыдущей. Если же в силу неблагоприятной ситуации не возникает, например, весной массовое размножение фитопланктона, то и все последующие группы будут угнетены, несмотря на благоприятные сочетания любых прочих факторов в период обычного появления группы.
Своеобразный вариант смен представляют поточные, или конвейерные, сукцессии. Они развиваются в подвижной среде: реках, потоках, круговоротах водных масс в океанах. Планктонные сообщества, увлекаемые водой, на разных стадиях развития оказываются в разных географических точках, а в одном и том же регионе постоянны по составу. Такой же конвейерный тип преобразований наблюдается, например, в лесной подстилке (рис. 156). В разных ее подгоризонтах: верхнем рыхлом слое опада, среднем спрессованном «ферментативном» подгоризонте и нижнем, полностью переработанном муллевом слое – существует постоянная значительная разница в многочисленном микронаселении. Но в каждой конкретной порции опада, поступающего сверху, происходит сукцессионная смена микроорганизмов, простейших, нематод и мелких членистоногих, обеспечивающая все более глубокое разложение растительных остатков. Опад вместе со всем населением постепенно занимает все более глубокое положение, захораниваясь под новыми поступлениями, а общая стратификация подстилки выглядит неизменной.
Рис. 156. Аналогии в сменах жизненных форм мелких членистоногих в изолированном опаде (вверху) и верхних слоях почвенного профиля (внизу) (по Н. М. Черновой, 1977):
1– поверхностные формы микроартропод;
2– полупочвенные микроартроподы;
Процесс сукцессии. Процесс сукцессии, по Ф. Клементсу, состоит из нескольких этапов: 1) возникновения незанятого жизнью участка; 2) миграции на него различных организмов или их зачатков; 3) приживания их на данном участке; 4) конкуренции их между собой и вытеснения отдельных видов; 5) преобразования живыми организмами местообитания, постепенной стабилизации условий и отношений.
В настоящее время практически вся доступная жизни поверхность суши занята различными сообществами, и поэтому возникновение свободных от живых существ участков имеет локальный характер. Это или места, освободившиеся в результате отодвигания ледников, отступания уреза воды в водоемах, обвалов, эрозии и т. п., или возникшие в результате деятельности человека, например выноса наверх больших масс глубинных пород при разработке полезных ископаемых.
Занос спор, семян растений, проникновение животных на освободившийся участок имеют большей частью случайный характер и зависят от того, какие виды есть в окружающих биотопах. Из числа видов, попавших в новое местообитание, приживаются лишь те, экологическая валентность которых соответствует данному комплексу абиотических условий. Прижившиеся виды постепенно занимают весь новый биотоп, вступая в конкуренцию друг с другом. В результате происходит перестройка видового состава и количественных соотношений разных форм. Параллельно идет процесс преобразования самого местообитания под влиянием развивающегося сообщества. Процесс завершается формированием более или менее стабильной системы с уравновешенным типом биологического круговорота.
Сукцессии любого масштаба и ранга характеризуются целым рядом общих закономерностей, многие из которых чрезвычайно важны для практической деятельности человека.
В любой сукцессионной серии темпы происходящих изменений постепенно замедляются. Конечным итогом является формирование относительно устойчивой стадии – климаксового сообщества, или климакса. Начальные, пионерные группировки видов отличаются наибольшей динамичностью и неустойчивостью. Климаксовые же экосистемы способны к длительному самоподдерживанию в соответствующем диапазоне условий, так как приобретают такие черты организации биоценозов, которые позволяют поддерживать сбалансированный круговорот веществ.
В ходе сукцессии постепенно нарастает видовое многообразие. Это ведет к усложнению связей внутри ценоза, разветвлению цепей питания и усложнению трофической сети, умножению симбиотических отношений, усилению регуляторных возможностей внутри системы. Тем самым уменьшается вероятность слишком сильного размножения отдельных видов и снижается степень доминирования наиболее массовых форм.
Чрезвычайное увеличение численности отдельных видов возможно чаще всего на начальных этапах развития сукцессионных серий, когда еще в недостаточной мере сложилась система взаимного регулирования. В незрелых сообществах, т. е. находящихся в начале сукцессионных рядов, преобладают мелкоразмерные виды с короткими жизненными циклами и высоким потенциалом размножения, специализированные на быстром захвате освободившегося пространства. Они обычно обладают широкими расселительными возможностями, позволяющими им первыми проникать на незанятые участки, но малоспособны к конкуренции и длительному удержанию за собой пространства.
Постепенно в развивающихся сообществах появляются и закрепляются более крупные формы с длительными и сложными циклами развития. Нарастание экологического разнообразия ведет к более четкому распределению групп организмов по экологическим нишам. В растительном покрове становится сильнее выражена ярусность и мозаичность, создающие основу пространственной структуры наземных экосистем. Усиливается зависимость успешного существования одних видов от биохимических выделений, роста или поведения других: преобладание конкурентных взаимоотношений сменяется преобладанием мутуалистических и трофических зависимостей.
В результате сообщества приобретают известную степень автономности и независимости от окружающих условий, не подчиняя свою жизнь флюктуациям внешней среды, а вырабатывая собственные эндогенные ритмы.
Не меньшие преобразования происходят и в энергетическом балансе системы (рис. 157). С энергетических позиций сукцессия – такое неустойчивое состояние сообщества, которое характеризуется неравенством двух показателей: общей продуктивности и энергетических трат всей системы на поддержание обмена веществ.
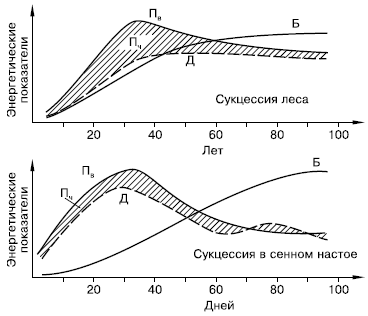
Рис. 157. Изменение энергетических показателей сообществ при развитии соснового леса (вверху) и лабораторной культуры простейших в сенном настое (внизу) (по Ю. Одуму, 1975):
Пв – продукция валовая; Д – траты на дыхание; Пч – продукция чистая (заштриховано); Б – биомасса
В ходе сукцессии общая биомасса сообщества сначала возрастает, но затем темпы этого прироста снижаются, и на стадии климакса биомасса системы стабилизируется. Это происходит потому, что на первых этапах сукцессии, когда видовой состав сообществ еще беден и пищевые цепи коротки, не вся часть прироста растительной массы потребляется гетеротрофами. Таким образом, относительно высока чистая продукция сообщества, которая идет на увеличение его биомассы. Накапливаются как общая масса живых организмов, так и запасы мертвого, неразложившегося органического вещества. В зрелых, устойчивых экосистемах практически весь годовой прирост растительности поступает и расходуется в цепях питания гетеротрофами, поэтому чистая продукция биоценоза, его «урожай», приближается к нулю.
Биогенные элементы растения сначала черпают из запасов почвы, но постепенно, по мере истощения этих запасов и накопления в системе мертвого органического вещества, разложение его становится основным источником минерального питания растений, и круговороты биогенных веществ из открытых превращаются в закрытые.
Неравенство трат энергии на образование первичной продукции и суммарный метаболизм (дыхание) сообщества проявляется в тех сукцессиях, в которых участвуют только гетеротрофные организмы. В этом частном случае первичная продукция равна нулю, и если приток мертвого органического вещества, за счет которого существует сообщество, не восполняет запасы в системе, то сообщество вскоре истощает свою энергетическую базу.
С удлинением цепей питания увеличивается эффективность использования поступающей энергии, иначе – КПД всей системы, так как одна и та же порция энергии идет на поддержание большого количества биомассы. Знание этих закономерностей имеет большое значение в практической деятельности человека.
Изымая избыток чистой продукции из биоценозов, находящихся в начале сукцессионных рядов, мы задерживаем сукцессию, но не подрываем основу существования сообщества. Вмешательство же в стабильные, климаксовые системы, с большой полнотой расходующие энергию на «свои» нужды, неминуемо вызывает нарушения сложившегося равновесия. Пока нарушения не превышают самовосстановительной способности ценоза, возникающие демутационные смены могут вернуть его к исходному состоянию. Этим пользуются, например, при рациональном планировании рубок леса. Но если сила и частота воздействия выходят за рамки этих возможностей, то первоначально устойчивое, богатое видами сообщество постепенно деградирует, сменяясь производными с малой способностью к самовозобновлению.
Вырубка леса на локальных участках с оставлением части территории под коренными типами лесной растительности вызывает ускоренные сукцессии, исходные фитоценозы восстанавливаются за относительно короткий срок – немногие десятилетия. Сплошные же рубки на больших площадях, особенно если используется мощная корчевальная техника, полностью разрушают не только лесное сообщество, но и весь почвенный покров, эволюция которого шла тысячелетия. В этом случае сукцессии приводят к иным, упрощенным типам сообществ, и на месте лесов возникают пустоши, болота или другие малопродуктивные экосистемы.
Таким образом, сообщество не может одновременно сочетать два противоположных свойства: быть высокостабильным и давать большой запас чистой продукции, который можно было бы изымать без вреда для самого ценоза.
Сукцессии, протекающие при разложении органического вещества в почве, лежат в основе биологического круговорота. Это естественные регуляторные процессы, восстанавливающие нормальное состояние нарушенных почв. Такие проблемы века, как подрыв естественного плодородия в результате нарушения процессов образования гумуса, загрязнение среды ядохимикатами и органическими отходами, эрозия, «утомление почв» и другие отрицательные явления, возникли вследствие ослабления регуляторных возможностей почв.
Стабильность экосистем, надежность протекания биологического круговорота веществ основаны на видовом разнообразии и полноте сукцессий.
Лекция 11. ДИНАМИКА БИОГЕОЦЕНОЗОВ: ФЛУКТУАЦИИ И СУКЦЕССИИ
Воронов А.Г. Геоботаника. Учеб. Пособие для ун-тов и пед. ин-тов. Изд. 2-е. М.: Высш. шк., 1973. 384 с.
Коробкин В.И., Передельский Л.В. Экология. Ростов на Дону: феникс, 2005. 576 с. (Высшее образование)
Основы лесной биогеоценологии / под ред. Сукачева В.Н. и Дылиса Н.В.. М.: Наука, 1964. 574 с.
Цветков В.Ф. Лесной биогеоценоз. Архангельск, 2003. 2-е изд. 267 с
Вопросы
1. О динамике фитоценозов
2. Флюктуации: определение понятия, типы флюктуаций
3. Сукцессии: определение понятия; серийные и коренные (климаксовые) сообщества, динамическое равновесие.
а) факторы, определяющие развитие растительности в первой фазе сукцессии – на оголенных территориях;
б) эцезис и его особенности;
в) стадии развития фитоценоза первичной сукцессии (по А.Г. Воронову);
г) стадии в развитии растительного покрова (по В.Н. Сукачеву);
д) суть понятий сингенез, эндоэкогенез и гологенез;
4.2. Вторичные сукцессии
5. Понятие климакса в биогеоценологии.
6. Теория подвижного равновесия.
7. Методология изучения вековых смен.
8. Создание рудеральных местообитаний и отвалов
ДОПОЛНИТЕЛЬНЫЙ МАТЕРИАЛ
б) классификация типов сукцессий по П.Д. Ярошенко;
в) кратковременные и вековые смены фитоценозов;
г) эволюция фитоценозов.
1. О динамике фитоценозов
Как растительный покров в целом, так и слагающие его фитоценозы никогда не остаются одинаковыми. Меняются и условия существования растительности, вызывая изменения растительного покрова, и сам растительный покров независимо от изменений условий существования (онтогенетические изменения, эволюция видов, образующих растительные сообщества, и т. д.).
В растительном сообществе можно выделить следующие типы изменений (Александрова, 1964): 1) суточные, 2) сезонные, 3) погодичные или разногодичные (флюктуационные), 4) возрастные, 5) изменения, определяемые процессами возобновления, 6) изменения, обусловленные микроэволюцией видов растений.
Рассмотрим эти разные проявления динамики фитоценоза.
а) суточные изменения – связаны с суточной ритмикой жизнедеятельности растений, образующих фитоценоз;
Определяются особенностями фитоклимата, создаваемого растительным сообществом. Выражаются в изменениях интенсивности транспирации, дыхания, фотосинтеза, в суточных движениях цветков и листьев, в ритме открывания и закрывания цветков.
б) сезонные различия определяются особенностями ритма развития видов, образующих фитоценоз. Постоянно наблюдаются в фитоценозах, ослабляясь или прекращаясь в неблагоприятные периоды (зима, сухой период) и усиливаясь в остальную часть года. Эти изменения позволяют существовать совместно большему числу видов растений, чем в том случае, если бы они развивались одновременно.
Весной – ранние эфемероиды, летом – позднелетние травы, кустарники, деревья.
в) погодичные или разногодичные изменения связаны с различиями в условиях среды обитания фитоценоза в разные годы, т.е. с изменением метеорологической обстановки по годам. От нее зависит интенсивность и особенности деятельности животных, обитающих в фитоценозе, а также урожайность семян, развития вегетативной массы растений и других особенностей биотопа и биоценоза, меняющихся по годам.
г) возрастные изменения – вызываются изменением возрастной структуры популяций растений, образующих фитоценоз, в частности возрастными изменениями (онтогенезом) эдификаторов.
Возрастные смены могут быть очень длительными, от нескольких лет до нескольких десятков лет, например, развитие лесного сообщества от стадии молодняка до приспевающего и спелого леса. Оно сопровождающееся не только изменением возраста дерева-эдификатора, но и соответствующими изменениями фитосреды и соотношений компонентов растительного покрова. Однако изменения фитосреды, т.е. режимов средообразующих факторов, не выходят за пределы определенной нормы, соответствующей растительным условиям, подходящим для данного типа сообщества.
Длительность прохождения фитоценозом возрастных стадий определяется условиями существования вида в данном фитоценозе и урожайностью семян.
ПРИМЕР. Б. П. Колесников (1968) указывает, что возрастных смен лесных фитоценозов сопровождаются существенными изменениями структуры и состава сопутствующих пород древостоя и нижних ярусов при сохранении лесообразующего значения господствующей (главной) породы. Эти смены протекают «внутри» лесной ассоциации (типа леса) и охватывают промежуток времени, как минимум, равный продолжительности жизни (периоду онтогенеза) одного поколения лесообразующей породы (40-300 лет в подавляющем большинстве типов леса умеренного пояса).
Циклы возрастных смен лесообразующей породы, повторяясь из поколения в поколение, постепенно накапливают изменения в режиме лесорастительных условий, что может привести к необратимым сменам типов леса.
ПРИМЕР. Некоторые парцеллы формируются после вывала старых деревьев. По мере смыкания крон, разрастания подлеска, они вновь сменяются прежними. Бывшими на данном месте. Такова маакиево-диморфантовая волжанковая парцелла в липово-широколиственном лесу с лиановой растительностью разнотравном. Она то и дело возникает в разных местах фитоценоза на ЭП «Горнотаежный». В лесу всегда отмирает дерево какого-либо вида, и на его месте вырастает другое, часто другого вида. Если осмотреть участок леса через 20-25 лет, расположение деревьев будет в значительной степени иным, чем ранее.
Периодически наблюдается массовое отмирание взрослых особей, связанное с метеорологическими аномалиями, массовыми размножениями вредителей, и т. п. Оно приводит к массовому развитию новых особей как того же самого, так и других видов. Обильное семенное возобновление обусловливает появление большого количества проростков, что меняет условия существования растений травяного яруса и мохового покрова и, следовательно, вызывает изменения в парцеллах, входящих в состав леса.
Такие смены, связанные с возобновлением эдификаторов, наблюдаются не только в лесных сообществах, но и в луговых. На лугах периоды, когда отмирание взрослых особей не выражено или выражено слабо, чередуются с периодами массового их отмирания.
Эти изменения протекают очень медленно и при обычном изучении биогеоценозов их выявить очень сложно.
2. Флюктуации: определение понятия, типы флюктуаций
Все вышеперечисленные изменения растительности происходят в пределах одного фитоценоза. При определенных условиях они могут выйти за его рамки:
— разногодичные изменения при увеличении амплитуды колебаний условий среды могут перейти в смены одного сообщества другим;
— массовое возобновление того или иного вида при гибели большей части его материнских особей способны привести к смене фитоценозов;
— процессы микрозволюции лежат в основе макроэволюции; вызывая изменения видов, накапливающиеся в пределах одного фитоценоза, приводят к смене одного фитоценоза другим.
1) изменения в пределах одного фитоценоза, не приводящие к последовательной смене эдификаторов сообщества, к резким изменениям его видового состава, структуры и других особенностей – флюктуации;
2) изменения, выражающиеся в смене одних сообществ другими – сукцессии.
Флюктуации – это обратимые изменения в фитоценозе, при которых основные компоненты сохраняются, хотя бы в латентном состоянии. По Т.А. Работнову, они могут вызываться погодичными изменениями метеорологических и гидрологических условий и антропогенным воздействием.
По степени выраженности различают три типа флюктуаций:
ПРИМЕРЫ. Вышеприведенный пример с формированием маакиево-диморфантовой волжанковой парцеллы в липово-широколиственном лесу после вывала старых деревьев является флюктуацией. По мере смыкания крон, разрастания подлеска, «волжанковая» парцелла вновь сменяется «разнотравной редкопокровной».
Изменения растительности, связанные с циклом активности солнечных пятен, при которых более влаголюбивые растения и растения более ксерофильные, «по очереди» находятся то в покоящемся состоянии, то в активном, несомненно, тоже представляют собой флюктуации.
Различают ненаправленные и направленные смены растительного покрова.
Ненаправленные представляют собой дальнейшее углубление процесса погодичных (разногодичных) изменений в пределах фитоценоза и это еще флюктуации.
Направленные смены представляют собой необратимые изменения растительного покрова, при которых одни сообщества сменяются другими. И это уже сукцессия. Необратимость процесса – обязательное свойство любой сукцессии, так же как обратимость – обязательное свойство флюктации.
3. Сукцессии: определение понятия; серийные и коренные (климаксовые) сообщества, динамическое равновесие
Термин сукцессия введен в 1901 г. американским ботаником Г. Каульсом (1869-1939). Развитие учения о сукцессии связано с именем его соотечественников, тоже ботаников Ф. Клементса и Д. Уивера. Детально изучали сукцессии и английские ботаники во главе с А. Тенсли, ученые франко-швейцарской школы, руководимые Ж. Браун-Бланке. В России изучение сукцессий лесов было начато Г.Ф. Морозовым и продолжено В.Н. Сукачевым, Е.М. Лавренко, П.Д. Ярошенко и другими лесоводами, а исследование смен растительности на степных залежах и выгонах – И.К. Пачоским, Г.Н. Высоцким, Л.Г. Раменским и др.

ПРИМЕР. Простым примером сукцессии служит зарастание озер заводей (рис. 1). Наземная расти-тельность, развивающаяся на месте сухого водоема, не имеет ничего общего с той водной растительностью, которая была характерна для него в годы наполнения его водой. Диаспоры (семена, части побегов, проростки) наземных растений попадают на высохшее днище или берег после ухода воды. И их приживание обеспечивает смену одного ценоза другим, завершаясь становлением сообществ кустарников и деревьев.
Сукцессия растительности – это последовательный ряд смены серийных (временно существующих) растительных сообществ на конкретном местообитании после выведения конкретной экосистемы из состояния динамического равновесия.
Смены растительности на вырубках или на залежах, приводящие к восстановлению коренной растительности, также считают сукцессиями.
Всю сукцессию в целом называют серией (американские авторы). Каждое сообщество, которое в процессе сукцессии сменяет другое, называют стадией этой сукцессии, или серийным сообществом (рис. 2).
Завершающая, относительно устойчивая стадия развития любого ценоза назы-вается климаксом (греч. klimax — лестница). На этой стадии сообщество приходит в равновесие со своей средой обитания.
Такое сообщество с присущим ему фитоклиматом состоит из «подогнанных» к совместному обитанию видов и в наибольшей степени соответствует условиям среды, как климатическим, так и эдафическим, и его геоботаники называют климаксовым (американские авторы), или узловым сообществом (П.Д. Ярошенко).
ПРИМЕРОМ экологического климакса может служить еловый лес Подмосковья, темнохвойная тайга в горах Восточной Сибири, ковыльные степи и влажные тропические леса.
Климаксовое сообщество принято также называть коренным. Оно считается устойчивым в данных климатических условиях, и все компонентами биогеоценоза находятся в «равновесии». Но об этом «равновесии» говорят, как о динамическом, подвижном.

Динамическое равновесие подразумевает непрерывную «настройку» и «перенастройку» биогеоценоза, так как никогда не прекращаются изменения внешних условий, а вместе с ними и популяций. В течение тысячелетий условия среды колеблются около некоторого среднего значения, и климакс изменяется значительно медленнее, чем остальные стадии сукцессии. Тем не менее, со временем происходят глубокие изменения в количественном и качественном составе климаксовой экосистемы.
НАПРИМЕР, около 10 000 лет назад в эпоху последнего оледенения на месте нынешних листопадных лесов востока Северной Америки находилась тундра.
Вполне понятно, что в силу своей «пригнанности» к условиям обитания климаксовая стадия гораздо более устойчива, чем начальная и переходные (серийные) стадии сукцессионного ряда. И всегда в ходе сукцессии увеличивается внутреннее разнообразие экосистемы, она становится более стабильной.
В равновесно-устойчивом, стабильном состоянии климаксовое, или коренное, сообщество пребывает до тех пор, пока не изменятся климат, рельеф, гидрологический режим, пока вновь не пройдет пожар, или не случится какая-то другая катастрофа. И вновь начнется новая сукцессия, которая либо приведет к восстановлению исходного сообщества, либо нет.
На одном и том же месте однотипная серия может повторяться неоднократно, если повторяются нарушения растительного покрова или его уничтожение, или резко меняются условия существования.
ПРИМЕР. Повсеместно растущие дубняки из дуба монгольского, сменившие в результате рубок и пожаров коренные кедрово-шроколиственные леса на южных склонах гор в Приморье.
4. Типы сукцессий
По типу развития растительного покрова сукцессии бывают первичными и вторичными.
4.1. Первичные сукцессии
Первичная сукцессия — это процесс развития и смены экосистем на участках, полностью свободных от растительного покрова и не сохранивших семенных зачатков растений.
Образование таких участков вызывается действием ветра (ветровая эрозия или ветровая аккумуляция), отступлением берега водоема и заливанием участка водой, выгоранием растительного покрова, выгрызанием растений животными, вырубкой леса, распашкой земель, т. е. климатическими, эдафическими, биотическими и антропогенными факторами.

Пример антропогенной сукцессии – формирование облепиховых зарослей на южном склоне на восточной окраине г. Уссурийск в долине р. Раковка и на одном из южных склонов гор Пржевальского (рис. 3).
История расселения облепихи в пригородной зоне г. Уссурийска связано со строительством в 1992-1993 гг. Уссурийской ТЭЦ и Уссурийского картонного комбината, под застройку которых были отведены дачные угодья на низком равнинном участке в долине р. Раковка. Грунт для строительства добывался на близлежащем склоне южной экспозиции, ранее занятом вторичным дубовым редколесьем с подлеском из лещины и разнотравно-осоковым ярусом. Во время разработки карьера на склоне был полностью уничтожены растительный покров и почвы.

Формирование облепиховых зарослей обусловлено одновременным сочетанием трех условий: 1 – отсутствием конкурентов по эконише – почвы и растительность при проведении строительных работ были полностью уничтожены на большей части склона, 2 – наличием на бывших дачных участках источников семян – плодоносящих куртин облепихи, 3 – обитанием в данном районе птиц, питающихся ягодами облепихи.
А.Г. Воронов (1940, 1973) выделяет в первичной сукцессии растительности на обнаженных почвах или грунтах две фазы:
— заселение оголенной территории и формирование фитоценоза из поселяющихся на голой площади растений (рис. 4).
— смена одного сформировавшегося фитоценоза другим (рис. 5).
а) факторы, определяющие развитие растительности в первой фазе сукцессии – на оголенных территориях
Растения проникают на освободившуюся территорию путем переноса диаспор (семян, спор, кусочков растения) при помощи ветра, воды, животных или человека, или путем постепенного вегетативного разрастания растений, находящихся близ границ оголенной территории. В составе нового фитоценоза часто преобладают растения с диаспорами, легко разносимыми ветром, а около воды – с диаспорами, хорошо держащимися на воде. Часто то действие, которое вызвало оголение почвы (отложение наноса водой, перевевание песка ветром) способствует и появлению на этой территории диаспор, т.е. деятельность человека. Поэтому так быстро на этих участках вырастают сорные и рудеральные растения.
Случаи внедрения растения на новую территорию с краев только путем образования вегетативных подземных или надземных побегов без образования генеративных органов наблюдаются во много раз реже, чем внедрение путем заноса семян.
Заселение новой территории зависит от ряда факторов, случайных по отношению к особенностям самой территории:
— от того, какие растения и как далеко они растут поблизости от нарушенного участка,
— от направления господствующего ветра,
— от высоты и силы паводка,
— от качества субстрата участков,
— от характера увлажнения, и т. д.
Следует отметить, что легкость семян, способствующая их переносу при помощи ветра, достигается за счет уменьшения запасов питательных веществ, а это отрицательно сказывается на развитии проростков, снижая шансы на их сохранение.
б) эцезис и его особенности
После того как растение проникло на оголенную территорию, оно начинает приспосабливаться к новым условиям. Процесс приспособления особей растений к новым для них условиям называют эцезисом. Он заканчивается тогда, когда растение дало плоды и семена.
Не все диаспоры, попавшие на оголенную территорию, прорастают сразу. Семена большинства видов долго сохраняют всхожесть, нередко десятки или даже сотни лет. При этом они прорастают не в один год, а при благоприятном стечении обстоятельств. Это обеспечивает условия лучшего сохранения проростков
ПРИМЕР. Леспедеца в дубняках вокруг пос. Горнотаежеое (Уссурийский р-н) возобновилась в первый же год после пожара, образовав сплошной покров. Пожара до этого не было более 20 лет. Лишь немногие виды (конский каштан, чозения, ивы, и др.) имеют семена, теряющие всхожесть в течение нескольких дней или немногих недель.
Семена, образующие почвенный запас, нередко принадлежат растениям разных жизненных форм и потому обеспечивают развитие растений при разнообразных условиях среды (семена одних видов прорастают при более высоких температурах, других – при более низких, одни при большей влажности грунта, другие – при меньшей и т. д).
Внедрившиеся на оголенную территорию растения начинают плодоносить и сами становятся источником диаспор. Теперь диаспоры попадают на заселяющуюся площадь не только со стороны, но и от тех растений, которые уже растут и плодоносят здесь же.
На стадии проростков многие растения, развивающиеся на оголенных территориях, гибнут под действием неблагоприятных условий, в результате уничтожения животными или паразитическими растениями и вследствие конкуренции с другими проростками.
В зависимости от условий существования оголенную территорию населяют один или несколько видов. Чем суровее условия, тем меньшее количество видов растений может начать здесь свое развитие. Наиболее бедный состав проростков типичен для сильно засоленных грунтов, обнажений скал и пр.
Когда растение из стадии проростка переходит в более поздние стадии развития, его потребности в воде и пище возрастают, а запасы питательных веществ в семени или в плоде к этому времени исчерпываются, и растение целиком зависит от пищевых ресурсов внешней среды. Поэтому с ростом растений усиливается и конкуренция. Чем суровее условия среды, тем большую роль для растений, внедряющихся на данную территорию, играет прямое влияние внешних условий и тем меньшее значение имеет конкуренция. Чем менее суровы условия среды, тем меньшую роль играют внешние условия и тем больше значение конкуренции.
в) стадии развития фитоценоза первичной сукцессии (по А.Г. Воронову)
Пионерная группировка – случайное сочетание растений. Фитоценозы, формирующиеся на оголенных площадях, на первой стадии развития характеризуются:
— случайным составом растений,
— отсутствием сомкнутого растительного ковра,
— слабым воздействием на среду и
— почти полным отсутствием взаимовлияния между особями.

Пионерная группировка может быть чистой (одновидовой, рис. 6), как в нижней части склона с облепиховыми зарослями, так и смешанной (многовидовой) – на том же склоне, на других участках. Если условия среды быстро меняются в направлении возрастания суровости (например, грунт высыхает, засоляется и пр.), то число видов, поселившихся на техногенном участке, уменьшается и смешанная пионерная группировка обедняется и, в конце концов, может перейти в чистую пионерную группировку.
Простая группировка – следующая стадия развития фитоценоза после пионерной группировки. В такой группировке растительный покров:
— в надземной части не сомкнут, однако растения располагаются значительно теснее, чем в пионерной группировке.
— хорошо заметно взаимовлияние растений.
— обычен групповой характер распределения растений: вокруг особи, давшей семена, развивается ее потомство.
Простые группировки, как и пионерные, могут быть чистыми (одновидовыми) и смешанными (многовидовыми), образованными несколькими видами, и растения в них, в отличие от смешанных пионерных группировок, всегда относятся к одной жизненной форме. Простые группировки обычно образованы некоторыми из видов, входивших в состав пионерных группировок.
Простые смешанные группировки, существующие весьма длительное время – сообщества однотипных (например, накипных) лишайников на камнях. Простыми группировками обычно представлена бурьянистая стадия залежи.
Сложная группировка – следующая за простой группировкой стадия развития фитоценоза. Она характеризуется следующими признаками:
ПРИМЕР. Зарастающая долина в районе строительства Уссурийской ТЭЦ. Сложные группировки здесь образуют рогоз (в мочажинах), донник, осоки разных размеров, мелкотравье. Растительный покров разрежен, но уже наметились ярусы: – рогоз – до 1 м, донник, лебеда, полынь, ослинник – 0,7-0,8 м, осоки – 0,4-0,5 м, мелкие злаки и травы высотой не более 10 см.
Стадия замкнутого фитоценоза – следующая стадия развития фитоценоза характеризуется:
Представлена двумя формами сочетаний растений – фитоценозами зарослей и 2 и более ярусных фитоценозов.
Заросли развиваются в условиях, в которых не может существовать сообщество из большого числа видов: сильная засоленность, сильная сухость, переувлажнение, высокая конкуренция, и т. п. Одноярусны. Ярус образован либо одним видом (чистые заросли), либо несколькими видами (смешанные заросли).
ПРИМЕР. Травяные заросли (чистые): сообщество тростника (Phragтites coтmunis) без участия более низкорослых растении, сообщество пырея ползучего (Agropyron repens), и др. Пример травяных смешанных зарослей – сообщество камыша Табернемонтана (Scirpus tabernaeтontani) и рогоза узколистного (Typha angustifolia), заросли крапивы и лебеды на заброшенных площадках из-под удобрений. Кустарниковые заросли – кедрово- и ольховниково-стланиковые – на Крайнем Северо-Востоке, ивняков и облепиховые в Южном Приморье, чозениевые в среднем возрасте на Дальнем Востоке. Сфагновые болота тоже можно считать моховыми зарослями – сфагнума.
Многоярусный фитоценоз (простой из 2-х ярусов, сложный – из более, чем 2-х ярусов), развивающийся не в столь суровых условиях, как заросли. Это все типы лугов (пойменные, суходольные, залежные), все лесные сообщеста. Не стоит думать, что со стадией развития фитоценоза завершается его динамика. Он вступает во вторую фазу процесса сукцессии: смену одного сформировавшегося фитоценоза другим.
Не во всех случаях фитоценоз обязательно проходит последовательно все перечисленные стадии – пионерная группировка ® простая группировка ® сложная группировка ® заросли или сложный фитоценоз. Этот путь бывает и проще, и сложнее.
ПРИМЕР. На скалах: нередко пионерная группировка сине-зеленых водорослей ® пионерная группировка лишайников ® смешанная или простая группировка лишайников ® смешанная длительно существующя заросль лишайников ® сложный фитоценоз с участием лишайников, мхов ® сложный фитоценоз ® цветковых растений.
На освободившемся из-под воды днище пресного озера: смешанная пионерная (гигро) группировка ® чистая пионерная группировка (ксерофит) ® чистая простая группировка ® смешанная простая группировка ® сложная группировка ® сложный фитоценоз. В других случаях чистую группировку сменяет чистая заросль, которая существует на этом участке неопределенно долгое время.
Таким образом, путь развития фитоценоза разнообразен: более длинный и более короткий, включает то одни, то другие стадии. Но во всех случаях развитие его идет от раздельно-группового сложения к диффузному, от несомкнутого покрова к сомкнутому, от незамкнутого покрова к замкнутому.
г) стадии в развитии растительного покрова по В.Н. Сукачеву
В.Н. Сукачев (1938, 1964 и др.) выделял следующие стадии формирования фитоценоза:
1. Отсутствие фитоценоза (соответствует пионерной группировке в начальной стадии ее существования).
2. Открытый фитоценоз (соответствует пионерной группировке в значительной части периода ее существования и простой группировке).
3. Закрытый невыработавшийся фитоценоз (соответствует сложной группировке).
4. Выработавшийся фитоценоз.
д) суть понятий сингенез, эндоэкогенез и гологенез
На самых первых этапах развития сообщества преобладает процесс, который В.Н. Сукачев (1942) назвал сингенезом. Это процесс первоначального формирования растительного покрова, связанный с вселением растений на данную территорию, их приживанием (эцезисом), а затем и конкуренцией между ними из-за средств жизни. Затем начинается другой процесс, названный В. Н. Сукачевым эндоэкогенезом. Это процесс изменения фитоценоза под влиянием среды, изменененной им самим. Эндоэкогенез постепенно усиливается и, в конце концов, становится основным процессом, определяющим ход смен фитоценоза.
На эти два процесса налагается третий, названный В. Н. Сукачевым (1954) гологенезом. Это «процесс изменения растительного покрова под влиянием всей географической среды или отдельных ее частей: атмосферы, литосферы и т.п., т.е. изменения более крупного единства, в состав которого входит данный биогеоценоз.
Все три процесса идут одновременно, но на разных стадиях развития преобладающее значение приобретает один из них. Несомненно, сингенез господствует только на начальных стадиях развития фитоценоза, а затем главенствующая роль переходит к эндоэкогенезу. Гологенетический процесс протекает постоянно, но, очевидно, в переломные моменты геологической истории Земли роль его усиливается.
Такой ход развития фитоценоза продолжается большее или меньшее время до тех пор, пока какие-то внешние силы, случайные по отношению к ходу развития фитоценоза, резко не нарушают его. Тогда смена, вызванная внутренним развитием самого фитоценоза (эндодинамическая), прерывается, и начинается смена, вызванная внешним толчком (экзодинамическая).
Исходя из сказанного выделяются два основных типа смен фитоценозов (Сукачев, 1928):
2. экзодинамические (Сукачев, 1928; Лавренко, 1940), или стихийные (Ярошенко, 1953), или внезапные (Ярошенко, 1961), возникающие под непредвиденным воздействием внешних факторов.
Причины возникновения сукцессий (смен) растительного покрова весьма разнообразны.
ПРИМЕРЫ эндодинамических смен – процессы заболачивания лесов в результате изменений в их напочвенном покрове, процессы зарастания водоемов, развития растительного покрова на скалах и обсохших днищах водоемов и т. д. Экзодинамические смены – смены растительного покрова на вырубках, залежах, пожарищах, участках, засыпанных лавиной или залитых селевым потоком, и т.д.
4.2. Вторичные сукцессии
Вторичная сукцессия – это восстановление экосистемы, когда-то уже существовавшей на данной территории, на местах разрушенных сообществ, где почва и часть организмов сохранились или смена одного (коренного) фитоценоза другим (производным, вторичным).
Если поле на месте вырубленного когда-то леса перестать обрабатывать, обычно за несколько стадий здесь вновь сформируется лесная экосистема, типичная для данного района. Процесс восстановления леса после пожара и вырубки также относится ко вторичной сукцессии.
ПРИМЕР вторичной сукцессии – восстановление елового леса на рис. 2. После вырубки или пожара условия на месте ельника настолько изменяются, что ель не может снова заселить освободившуюся площадь. На открытых местах всходы ели повреждаются поздними весенними заморозками, страдают от перегрева и не могут конкурировать со светолюбивыми растениями. В первые два года на вырубках и гарях буйно развиваются травянистые растения: кипрей, вейник и др. Вскоре появляются многочисленные всходы березы, осины, а иногда сосны, семена которых легко разносятся ветром. Деревья вытесняют травянистую растительность и постепенно образуют мелколистный или сосновый лес.
Только тогда возникают условия, благоприятные для возобновления ели. Теневыносливые всходы ели успешно конкурируют с подростом светолюбивых лиственных пород. Когда ель достигает верхнего яруса, она полностью вытесняет лиственные деревья, затеняя их. Так же протекает сукцессия пихтово-кедровой тайги. Например, сукцессия сибирского темнохвойного леса (пихтово-кедровая тайга) после опустошительного пожара.
Типы вторичных сукцессий (по В.М. Урусову):
5. Понятие климакса в биогеоценологии
В процессе сукцессий возникают биогеоценозы, в наибольшей степени соответствующие условиям среды, как климатическим, так и эдафическим, а также состоящие из «подогнанных» к совместному обитанию видов со свойственным этому ценозу фитоклиматом и гидрологическим режимом. Среда обитания внутри такого ценоза им и была преобразована. Эта завершающая стадия сукцессии получила название климаксовой, климакса.
Вполне понятно, что в силу своей «пригнанности» к условиям обитания климаксовая стадия гораздо более устойчива, чем начальная и переходные стадии данной серии. Тем не менее, и климакс продолжает изменяться, хотя значительно более медленно, чем остальные стадии. Эти изменения объясняются тем, что никогда не прекращаются изменения внешних условий, вызывающие гологенез, и соответствующая перестройка фитоценоза.
Климакс, по мнению основателя теории климакса Клементса, соответствует формации в понимании американских авторов, т. е. зональному типу или в некоторых случаях подтипу растительности в понимании российских исследователей. Так, формацией следует считать всю зональную растительность тундровой зоны или зональную растительность подзоны хвойных лесов, или зональную растительность подзоны широколиственных лесов. Климакс постоянен до тех пор, пока не изменится климатическая обстановка. Стабильность климакса может быть нарушена только человеком или изменением климата.
Сторонники поликлимакса для всех местообитаний различают сообщества, находящиеся в условиях относительного соответствия местообитанию (климаксы), и сообщества, быстро изменяющиеся (стадии серии, или серийные сообщества). Согласно этим воззрениям, каждое местообитание имеет свой климакс. Этим термином можно и следует пользоваться, понимая под климаксовыми такие сообщества, которые находятся в относительном соответствии с условиями существования и потому изменяются весьма медленно, значительно медленнее, чем сообщества, представляющие собой стадии сукцессионных серий.
Следует отметить, что в настоящее время сторонников теории моноклимакса по существу нет.
6. Теория подвижного равновесия
Очень близка к теории климакса теория подвижного равновесия растительного покрова, которую в 20-е годы в России развивали А. А. Еленкин и Л. Г. Раменский. Сочувственно относились к ней и некоторые другие исследователи. За рубежом ее развивали Р. Cepнандep (Sernander, Швеция), А. Тенсли (Tansley, Англия), Каулс (Кawlеs, США) и др.
Согласно этой теории, все сообщества можно разделить на три категории: равновесные, равновесно-сменные и нарушенные (Алехин, 1928).
Равновесные сообщества характеризуются тем, что их строение и состав колеблются около какой-то средней точки, представляющей как бы типичное состояние растительного покрова. К их числу относится большая часть фитоценозов.
Равновесно-сменные сообщества отличаются от равновесных тем, что сама точка равновесия постепенно смещается в каком-либо направлении, и колебание особенностей сообщества совершается вокруг этой движущейся точки.
Примером равновесно-сменных сообществ могут быть фитоценозы поймы, где одни участки все время меняются в сторону высыхания, другие – в сторону заболачивания.
Нарушенными сообществами называют такие, которые из-за резкой смены условий выведены из состояния равновесия. Эти сообщества стремятся вернуться к тому состоянию равновесия, из которого они были выведены внешней по отношению к ним силой. Таковы сообщества, возникающие при распашке, порубках, пожарах и т. д.
Сторонники этой теории, как и некоторые сторонники теории климакса, все развитие сообщества объясняют воздействием внешних факторов. При этом из их поля зрения полностью выпадает самодвижение, развитие фитоценоза в результате действия внутренних причин.
Таким образом, теория подвижного равновесия весьма близка к теории климакса в ее первоначальном варианте, когда полагали, что климакс не меняется без изменения внешних условий, и отличается от нее не принципиально, а лишь постановкой вопроса в другой плоскости. Равновесные сообщества соответствуют климаксу (поликлимаксу), но сторонники теории подвижного равновесия подчеркивают колебательные изменения этих сообществ вслед за колебаниями условий среды, а сторонники климакса – неподвижность этих сообществ без изменения условий среды.
В свое время теория подвижного равновесия в применении к растительным сообществам сыграла до некоторой степени положительную роль, так как заставила внимательнее присмотреться к погодичным, флюктуационным изменениям сообществ в зависимости от условий погоды разных лет, к влиянию на фитоценозы медленных изменений среды (например, в поймах рек). Однако решала она эти вопросы неправильно, с механистических позиций, не дооценивая явления самодвижения, саморазвития фитоценозов.
Но! Смена одного сообщества другим совершается в результате изменения конкурентных отношений между видами, образующими сообщество. При сукцессиях, за исключением катастрофических смен, происходящих, например, под влиянием наводнений, пожаров, смена одного сообщества другим идет постепенно: часть видов первого сообщества исчезает быстрее, часть – медленнее, часть остается в новом сообществе, нередко более или менее резко меняя свое фитоценотическое значение. При этом растения одного фитоценоза попадают в другой, конкурируют с компонентами последнего, вытесняют их, занимая их место. При успешном развитии этого процесса количество «наступающих» особей и видов все увеличивается, меняется и структура подвергающегося «агрессии» фитоценоза под влиянием изменения видового состава и соотношения видов, а также изменений среды, и в конечном итоге первый фитоценоз вытесняется вторым. Именно изменение не только видового состава, но и структуры (и сезонного хода развития) фитоценоза позволяет утверждать, что здесь имеет место не только борьба отдельных видов растений, но и фитоценозов в целом, представляющих определенные единства. Эта борьба фитоценозов осуществляется в конкретных условиях и нельзя считать один из фитоценозов во всех случаях более сильным, а другой более слабым. Сила и слабость фитоценозов зависят от экологических условий.
В заключение: Все изменения направлены на повышение конкурентной устойчивости в сообществе: зрелые экосистемы, завершающие сукцессионные серии, характеризуются высоким видовым разнообразием, что ведет к усложнению связей внутри биоценоза, развитию детритных цепей питания, умножению симбиотических связей. В таких экосистемах продуцирование органического вещества уравновешивается его размножением. Потери вещества минимальные, круговороты химических элементов почти замкнутые.
7. Создание рудеральных местообитаний и отвалов
В результате многообразной деятельности человека возникают местообитания, не свойственные природе – так называемые рудеральные, или мусорные. Такие местообитания обычны в селениях, вдоль дорог, по межам полей. Их характеризует прежде всего повышенное содержание азота, так как они всегда богаты полуперегнившими и сгнившими растительными остатками. Поэтому в этих местообитаниях особенно часто поселяются так называемые нитрофильные растения.
Типичный представитель нитрофильных растений – крапива двудомная (Urtica dioica), которая, как указывает Г. Вальтер (Вальтер – Алехин, 1936), вообще не растет на почвах с содержанием нитратов менее 40 мг/л. Интересно, что в природных условиях это растение встречается либо в ольшаниках, где микроорганизмы, живущие на корнях ольхи, обогащают почву азотом, либо в зарослях тростника (Phragmites communis), где весьма обильны мертвые органические остатки, образующие род торфа.
Сходство рудеральных местообитаний, выражающееся в богатстве нитратами и другими соединениями азота, влечет за собой сходство их флоры, в которой заметную роль играют космополитные (встречающиеся на значительной части суши земного шара) растения. Однако отдельные рудеральные местообитания отличаются содержанием в почвах различных солей, прежде всего солей щелочных металлов – натрия и калия, степенью увлажнения почвы, климатическими особенностями. Поэтому наряду с космополитами здесь произрастает значительное число видов локальных (местных). Сообщества рудеральных растений в различных географических регионах резко отличны друг от друга.
Отвалы, образующиеся при открытых разработках полезных ископаемых, а также терриконы, представляющие собой кучи выброшенной из шахт породы, имеют огромную массу и нередко занимают значительные площади. Отрицательное воздействие открытых разработок распространяется на прилегающие территории, значительно превосходящие по площади сами выработки.
Рекультивация отвалов затрудняется тем, что выброшенные на поверхность грунты нередко оказываются токсичными.
Лесной рекультивации отвалов или их сельскохозяйственному освоению должна предшествовать подготовка поверхности отвалов. Сюда входит разравнивание и планировка, устройство дренажа, применение различных химических веществ для мелиорации токсичных горных пород, нанесение на поверхность выровненных отвалов плодородных пород и почв (в тех случаях, когда заблаговременно убранные почвы сохраняются до окончания горных работ) и пр.
Работы по рекультивации отвалов и терриконов в настоящее время стали одним из важнейших разделов восстановления нарушенных деятельностью человека ландшафтов
ДОПОЛНИТЕЛЬНЫЙ МАТЕРИАЛ
Классификация типов сукцессий по П.Д. Ярошенко
П.Д. Ярошенко (1961), разрабатывая теорию сукцессий, составил более подробную следующую классификацию типов смен:
А. Природные смены.
а) эндоэкогенетические (включают и сингенетические смены В.Н. Сукачева), б) гологенетические.
а) климатогенные, б) эдафогенные, в) биогенные.
Б. Антропогенные смены:
а) последовательные, б) внезапные.
В данной системе природные внезапные смены и антропогенные смены соответствуют экзодинамическим сменам В.Н. Сукачева, поскольку с точки зрения существования растительного сообщества безразлично, какими обстоятельствами вызвана серия смен: вырубкой леса, его уничтожением в результате пожара или в результате действия насекомого-вредителя, хотя ход процессов при этом будет неодинаков.
Любая смена, начавшаяся под влиянием внешних причин (экзодинамически), в дальнейшем постепенно переходит в эндодинамическую, так как ее ход во все большей степени определяется воздействием фитоценоза на среду. Если вновь не повторятся воздействия экзогенных факторов на фитоценоз.
Кратковременные и вековые смены фитоценозов
По длительности сукцессии подразделяются на кратковременные (частные), вековые (общие) и филоценогенетические (эволюция фитоценозов). Все перечисленные типы смен очень тесно сплетаются друг с другом и накладываются один на другой. В группу кратковременных, или частных, смен объединяют эндодинамические и экзодинамические. Им противопоставляются вековые (Лавренко), или общие (Ярошенко), смены. Последние протекают значительно медленнее и охватывают территорию, занимаемую не одним или немногими связанными друг с другом фитоценозами, а обширные территории.
Многочисленные вековые смены растительного покрова, происходившие в различных районах суши, могут быть до известной степени выяснены с помощью спорово-пыльцевого анализа, широко вошедшего в практику историко-фитоценологических исследований.
Некоторое представление о прежнем растительном покрове дают и так называемые реликтовые фитоценозы, сохранившиеся в том или ином районе от прошлых геологических периодов.
ПРИМЕРЫ. Так, степные острова в долинах рек Амги (приток Алдана), Лены, Вилюя, Яны и по их склонам или в районе так называемой Кунгурской островной лесостепи (Пермская область) свидетельствуют о том, что степь ранее (очевидно, во время так называемого ксеротермического пустынно-степного периода) была здесь широко распространена. Торфяные сфагновые болота, встречающиеся в подзоне южных степей (Кустанайская область, Казахская ССР), молоды, но они возникли в так называемом субатлантическом периоде, когда климат был значительно более влажным, чем в настоящее время. Сохранение таких реликтовых фитоценозов, чуждых общей климатической обстановке района (степи среди тайги, сфагновыe болота среди южных степей), связано с местными условиями рельефа, почв, вод, создающими возможность сохранения здесь этих фитоценозов.
Между кратковременными и вековыми сменами существуют переходы, хотя в общей форме можно сказать, что в результате кратковременных смен вырабатываются растительные ассоциации и формации в пределах географических зон, а в результате вековых смен растительность одной зоны сменяется растительностью другой.
ПРИМЕРЫ кратковременных смен: смены сосняков из сосны могильной – сухими дубняками, хвойно-широколиственных лесов – смешанными широколиственными лесами, белоберезняками, осинниками, чозенников – лиственничниками; вековых смен: изменения растительного покрова на севере европейской части СССР после отступления ледника и на юге Дальнего Востока – после отступления-наступления моря. Ксерофитизация климата?
От вековых смен следует отличать возникновение и эволюцию различных типов растительности.
Изучение динамики растительного покрова, различных смен как кратковременных, так и вековых, позволяет человеку овладеть этим процессом и направить развитие растительного покрова по тому пути, который наиболее целесообразен.
Наконец, различают филоценогенез, или эволюцию фитоценозов.
Эволюция фитоценозов и филоценогенетические сукцессий.
Эволюция фитоценозов, как указывал В. Н. Сукачев (1954), слагается из двух процессов: выработки структуры ценоза (филоценогенез) и видообразования, идущего в связи с эволюцией ценозов (флорогенез).
Эволюции растительных сообществ совершается также путем естественного отбора, который В. В. Ревердатто (1935) предложил называть естественным отбором второго порядка (в отличие от естественного отбора первого порядка – отбора особей в процессе видообразования. Благодаря естественному отбору существует определенное число фитоценозов, а не бесчисленное количество комбинаций, сочетаний растений. В процессе эволюции выработались как растительные ассоциации, так и более крупные единицы растительного покрова вплоть до типов растительности.
В качестве примеров можно привести примеры частных смен фитоценозов, происходящих в различных условиях.
Существует два способа зарастания водоемов: со дна и с поверхности. Детали этого процесса в разных районах и в водоемах разных типов неодинаковы, но общий ход его для каждого из этих способов один и тот же. О ходе сукцессий в водоемах можно судить, исследуя отложения сообществ, сменявших друг друга на данной территории, или путем изучения поясов растительности, сменяющих друг друга по мере изменения условий существования в пространстве.
Процесс зарастания водоема со дна
При зарастании пресноводных стоячих водоемов сo дна, чаще всего при пологих берегах, наблюдается несколько стадий. Связь между стадиями заключается в оседании на дно остатков организмов и повышения уровня дна. Это делает возможным продвижение пояса больших растений – макрофитов, связанных с определенными глубинами, от первоначального берега к центру водоема и приводит к смене озера болотом. На болоте также по мере накопления торфа и изменения условий существования одна растительная ассоциация сменяет другую.
ПРИМЕР № 1. При зарастании водоема со дна обычно наблюдаются следующие стадии (Лепилова, 1934):
1. Господство планктона. Глубины еще слишком значительны, чтобы зеленые растения-макрофиты могли здесь обитать.
2. Подводные луга с глубины 5, иногда 6 м, до нижнего предела существования растительности макрофитов. Здесь растут водяные мхи скорпидиум (Scorpidiuт), каллиергон (Calliergon) и др., виды харовых водорослей (Chara), иногда сюда спускается элодея канадская (Elodea canadensis).
3. Заросли широколиственных рдестов, развитые на глубинах 3-5 м (иногда до 6 м). Образованы погруженными в воду растениями, выносящими на поверхность воды или поднимающими над ее поверхностью свои цветки для опыления. Здесь наиболее часты рдесты пронзеннолистный и блестящий (Potaтogeton perfoliatus, P. lucens) и др.
4. Заросли растений с плавающими листьями, развитые на глубинах 2-3 м, иногда более (до 5 м), обычно в защищенных от ветра местах: в заливах, заводях, старицах и т. д. Для этих зарослей характерны кубышки (Nuphar), кувшинки (Nyтphaea), рдест плавающий (Potaтogeton nataпs).
5. Пояс камыша и тростника. Развит с глубины 0,5 до 2-3 м, редко глубже. Здесь растут виды, поднимающие свои стебли высоко над поверхностью воды. Это камыш озерный (Scirpus lacustris), тростник обыкновенный (Phragтites coттuпis), рогозы (Typha), манник водяной (Glyceria aquatica), тростянка (Scolochloa) и др. На этой стадии дно водоема покрыто мощным слоем ила.
6. Прибрежная растительность мелководий. Обитает от уреза воды до глубины 0,5 м, т.е. до пояса камыша и тростника. Характер дна на этой стадии очень изменчив. Оно бывает илистым, песчаным и каменистым (песчаное и каменистое дно обычны в северных, слабо зарастающих озерах). Характерны для этой стадии осоки, главным образом осока вздутая (Саrех rostrata), рдест разнолистный (Potaтogetoп heterophyllus), хвощ приречный (Equisetuт heleocharis).
7. Прибрежная растительность. Занимает участки, расположенные выше уреза воды, но по временам заливаемые водой при высоком ее уровне или при сильном прибое. Дно здесь может быть илистым, торфянистым, песчаным, каменистым, глинистым и т. д. Растения, характеризующие эту стадию, относятся к числу земноводных, иногда болотных. Здесь характерны вех (Cicuta), ситняги болотный и игольчатый (Heleocharis palustris, Н. acicularis), лютик стелющийся (Raпuпculus reptaпs), осоки (Саrех) и др.
Дальнейшее развитие сукцессий может привести к возникновению сообществ кустарников и деревьев.

ПРИМЕР № 2. Зарастание со дна заводей в Полабье (Чехословакия) (рис. 7):
1) водное сообщество Myriophylleto-Nupharetum;
2) сообщество Parvocharetum, подготовливающее заполнение заводи подстилкой, пропитанной известняком;
3) заросли тростника – Phragтitetum;
4) заросли Cladiuт-Cladietum marisci;
5) пояс дерновинных осок (сообщество из группы Magno-caricetum);
6) заросли Schoeпus-Schoenetum nigricantis; уровень грунтовых вод находится ближе к поверхности, как и в …
7) … сообществе Moliпia-Molinietum caficeto-sumpaniceae, которое вытесняется на крепкой почве стадией …
8) … верб и крушины Franguleto-Salicetum, подготовляющей почву для …
9) … ольшаника – ассоциации Alпus glutiпosa-Dryopteris thelypteris.
Процесс зарастания водоемов с поверхности

ПРИМЕР № 3. Совершенно иначе зарастают водоемы с поверхности (заболачивание путем нарастания) При этом с берега на поверхность воды наплывает зыбучий ковер из мхов и некоторых цветковых растений, с мощными корневищами (рис. 8). Такой ковер называется сплавиной, или зыбуном. Сплави-на образуется только в защищен-ных от ветра местах с относитель-но круто уходящим в глубь от берега дном.
Сплавина разрастается, от ее нижней поверхности отрыва-ются куски торфа, перегнившие и полуперегнившие остатки расте-ний и на дне образуется мощный слой полужидкого ила. Постепен-но весь водоем заполняется кашицеобразной массой, а с поверхности его покрывает сплошной слой сплавины, в котором просвечивают «окна» – отверстия или участки, где сплавина очень тонка. Попав в такое «окно», человек или животное погибает в трясине. После заполнения водоема илом на его месте развивается болото.
Процесс образования болот

В зависимости от условий местности даль-нейшее развитие зарос-ших водоемов приводит или к возникновению лесной растительности, или к образованию верхо-вого болота (рис. 9), лишенного связи с грунто-выми водами, имеющего атмосферное питание и характеризующегося гос-подством сфагнума.
Причины заболачивания суши различны.
1. Материковые луга могут заболачиваться в результате уплотнения почвы под влиянием весеннего выпаса скота.
2. Существенная причина заболачивания материковых лугов – обеднение почв. Питательные вещества выносятся животными с поедаемыми растениями или вывозятся со скошенным сеном. Более требовательные к питательным веществам ценные травы постепенно исчезают, появляются растения, мирящиеся с бедностью почвы, среди которых большую роль играют мхи.
3. После вырубки леса или после лесного пожара в районах с неглубоким уровнем грунтовых вод, также начинаются процессы заболачивания, которые связаны с подъемом грунтовых вод по почвенным капиллярам, вследствие прекращения вытягивания воды из почвы деревьями.

4. На севере лесной зоны заболачивание может вызвать естественная смена зеленых гипновых мхов кукушкиным льном и сфагнумом (рис. 10). Кукушкин лен удерживает большое количество воды, что влияет на степень увлажнения почвы. Когда влажность становится чрезмерной для кукушкина льна, он сменяется сфагновыми мхами, а те, способствуя сохранению вечной мерзлоты еще более способствуют заболачиванию. Сфагновые мхи появляются сначала небольшими пятнами, а затем постепенно завоевывают господство, могут и не предваряться зелеными мхами.
Одинаковые фитоценозы могут иметь совершенно различное происхождение, т. е. могут возникать полифилетически.
Зарастание горных территорий с каменистыми субстратами
Ход и характер сукцессий определяется составом пород, образующих скалы, крутизной и экспозицией скалистого склона.
Очень медленно происходит завоевание растительностью скал. На обнаженных скалах сначала поселяются бактерии, синезеленые и зеленые водоросли, затем лишайники. Лишь спустя очень долгое время, когда в результате выветривания и деятельности бактерий и лишайников на поверхности скалы местами образуется незначительный почвенный слой, здесь могут развиваться мхи. После поселения мхов накопление частиц почвы происходит быстрее. Спустя еще некоторое время на скалах появляются папоротникообразные и цветковые растения, сначала по трещинам и расщелинам, где скапливаются органические вещества и вода, а затем на их поверхности.

ПРИМЕР № 4. На известняковых скалах в районе Праги, на склонах южной экспо-зиции, сменяют друг друга сле-дующие сообщества (рис. 11):
1 – папоротник Aspleпiuт rutaтuraria в трещинах скал;
2 – сообщество Festuca duriuscula-Seseliesseum на сильно скелетной неглубокой почве, которую оно скрепляет, обогащает гумусом и подготовляет для следующего сообщества;
3 – сообщество Carex huтilis-Festuca sulcata, которое преобладает на более пологих ступенчатых склонах с почвой менее скелетной и более омощной;
4 – на более глубоких почвах пологих склонов развито замкнутое сообщество Festuca vallesiaca-Erysiтuт crepidifoliuт, в которое проникают …
5 – … кустарники, особенно степная вишня – Prunus fruticosa, предшествующая …
6 – … грабовому лесу с дубом краснеющим или теплолюбивой дубраве (Quercetum-Carpinetum primuletosum).
На верхушках скал стадии 4-6 мозаично сменяют друг друга, внешнеи напоминая (образуя) «лесостепь». Местные причины могут изменить ход сукцессии или привести к фрагментарному выражению ее стадий (2-6).

На склонах северных и западных экспозиций сукцессии происходят иначе, чем на южных.
На рис. 12 показана ярусность в лесной серии сукцессий. Показаны три горизонта почв (А1, A2 и В); растительность образует над поверхностью почвы по существу один ярус в пионерном сообществе Danthonia (6), но число ярусов возрастает до четырех, отчетливо различи-мых в лесу из клена: травы, кустарники, низкие деревья, высокие деревья.
Смены биогеоценозов могут быть прогрессивными и регрессивными. Под прогрессивными сменами обычно понимают такие, которые связаны с увеличением продуктивности и усложнением строения фитоценоза. Смены, связанные с уменьшением запасов растительной массы и упрощением строения фитоценоза, следует считать регрессивными. Так, заселение растительностью скал служит примером прогрессивной смены, а заболачивание лесов – пример регрессивной смены.
Человек заинтересован в том, чтобы направлять смены растительного покрова в сторону прогресса. Для управления этими процессами не обходимо знание направления и темпа сукцессий. Вопросы возобновления леса, развития растительности на пастбищах, озеленения терриконов и выемок и многие другие могут быть разрешены только после тщательного изучения сукцессий.
БЕСПРЕДЕЛЬНЫ ЛИ СУКЦЕССИИ ИЛИ ОНИ ЗАКАНЧИВАЮТСЯ КАКИМ-ЛИБО ФИТОЦЕНОЗОМ, ДАЛЕЕ НЕ МЕНЯЮЩИМСЯ? КАК ПРОХОДЯТ САМИ ИЗМЕНЕНИЯ?
Развитие растительного покрова никогда не останавливается. Он меняется в силу своего взаимодействия со средой (процессы сингенеза и эндоэкогенеза). Меняя среду, фитоценоз меняется сам.
Изменения в составе фитоценоза происходят непрерывно, быстро или постепенно. В ходе этого непрерывного развития неизбежно приходится выделять стадии, поскольку определенная группа видов, доминирующая в данный момент, уступает доминирование другой группе видов, а влияние первой группы уменьшается.
Если в силу каких-либо причин растительный покров внезапно нарушен, он восстанавливается относительно быстрыми темпами, но если таких резких нарушений нет, изменения растительного покрова протекают более медленно. Эти изменения происходят даже в тех случаях, когда климатические условия относительно постоянны. При этом фитоценоз изменяется за счет микроэволюции образующих его видов, т. е. путем все более глубоких приспособлений этих видов к условиям фитосреды и друг к другу.
Темпы сукцессий неодинаковы. На участке, лишенном растительного покрова, первые стадии сменяют друг друга через год несколько лет, а далее процесс смен замедляется и более позднее стадии восстановления или формирования растительного покрова занимают десятилетия, а затем и столетия. Сложившиеся фитоценозы сменяют друг друга уже в течение тысяч, десятков и сотен тысяч лет. Наконец, продолжительность филоценогенетических смен соразмерна с продолжительностью геологических периодов.
Изменения фитосреды, вначале благоприятные для фитоценоза, затем могут стать для него неблагоприятными, что приводит к смене одного фитоценоза другим. Процесс создания фитоценозом неблагоприятных условий для своего существования на первых стадиях развития идет быстрее, при этом обычно быстрее изменяются и условия среды. Все это влечет за собой более скорую смену одного фитоценоза другим; на более поздних стадиях развития фитоценоза эти процессы идут медленно и соответственно более медленно идут смены стадий сукцессии. Вымирания видов под влиянием прямого неблагоприятного действия фитосреды, как правило, не наблюдается; постепенно среда становится благоприятнее для видов, которые имеют в фитоценозе подчиненное значение, и менее благоприятной для видов, господствующих на данной стадии в фитоценозе. В результате первые получают преимущество в борьбе за существование со вторыми и постепенно вытесняют их.
Растения, которые при смене фитоценозов остаются в новых фитоценозах, приспосабливаются к новым условиям фитосреды и образуют новые биотипы и экотипы.



