Синапс

Основные элементы синапса
Си́напс (от греч. sýnapsis — соединение, связь, обнимать, обхватывать) — место контакта между двумя нейронами или между нейроном и получающей сигнал эффекторной клеткой. Служит для передачи нервного импульса между двумя клетками, причём в ходе синаптической передачи амплитуда и частота сигнала могут регулироваться. [1]
Термин был введён в 1897 г. английским физиологом Чарльзом Шеррингтоном.
синапс—особая структура, обеспечивающая передачу нервного импульса с нервного волокна на какую-либо другую нервную клетку или нервное волокно, также с рецепторной клетки на нервное волокно(область соприкосновения нервных клеток друг с другом и другой нервной клеткой). Для образования синапса необходимы 2 клетки.
Содержание
Структура синапса
Классификации синапсов
В зависимости от механизма передачи нервного импульса различают
так как сопротивление внеклеточной жидкости мало(в данном случае), импульсы проходят не задерживаясь через синапс. Электрические синапсы обычно бывают возбуждающими.
Наиболее распространён первый тип.
Химические синапсы можно классифицировать по их местоположению и принадлежности соответствующим структурам:
В зависимости от медиатора синапсы разделяются на
При этом в синапсе не всегда вырабатывается только один медиатор. Обычно основной медиатор выбрасывается вместе с другим, играющим роль модулятора.
Если первые способствуют возникновению возбуждения в постсинаптической клетке (в них в результате поступления импульса происходит деполяризация мембранны, которая может вызвать потенциал действия при определённых условиях.), то вторые, напротив, прекращают или предотвращают его появление, препятствуют дальнейшему распространению импульса. Обычно тормозными являются глицинергические (медиатор — глицин ) и ГАМК-ергические синапсы (медиатор — гамма-аминомасляная кислота). Таким образом, тормозные синапсы бывают двух видов: 1) синапс, в пресинаптических окончаниях которого выделяется медиатор, гиперполяризующий постсинаптическую мембрану и вызывающий возникновение тормозного постсинаптического потенциала; 2) аксо-аксональный синапс, обеспечивающий пресинаптическое торможение. Синапс холинергический (s. cholinergica) — синапс, медиатором в котором является ацетилхолин.
В некоторых синапсах присутствует постсинаптическое уплотнение — электронно-плотная зона, состоящая из белков. По её наличию или отсутствию выделяют синапсы асимметричные и симметричные. Известно, что все глутаматергические синапсы асимметричны, а ГАМКергические — симметричны.
В случаях, когда с постсинаптической мембраной контактирует несколько синаптических расширений, образуются множественные синапсы.
К специальным формам синапсов относятся шипиковые аппараты, в которых с синаптическим расширением контактируют короткие одиночные или множественные выпячивания постсинаптической мембраны дендрита. Шипиковые аппараты значительно увеличивают количество синаптических контактов на нейроне и, следовательно, количество перерабатываемой информации. «Не-шипиковые» синапсы называются «сидячими». Например, сидячими являются все ГАМК-ергические синапсы.
Механизм функционирования химического синапса
Открыты два механизма высвобождения: с полным слиянием везикулы с плазмалеммой и так называемый «поцеловал и убежал» (англ. kiss-and-run ), когда везикула соединяется с мембраной, и из неё в синаптическую щель выходят небольшие молекулы, а крупные остаются в везикуле. Второй механизм, предположительно, быстрее первого, с помощью него происходит синаптическая передача при высоком содержании ионов кальция в синаптической бляшке.
Так называемый «принцип Дейла » (один нейрон — один медиатор) признан ошибочным. Или, как иногда считают, он уточнён: из одного окончания клетки может выделяться не один, а несколько медиаторов, причём их набор постоянен для данной клетки.
Что такое синапсы кратко
Область контакта между двумя нейронами называют синапсом.
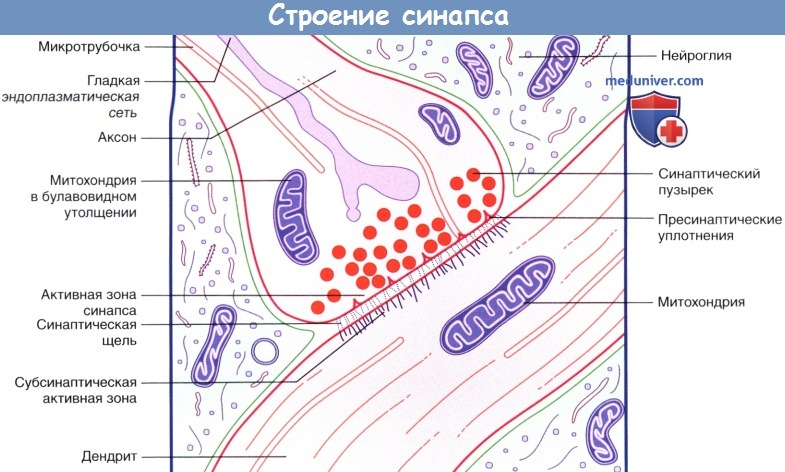 Внутреннее строение аксодендритического синапса.
Внутреннее строение аксодендритического синапса.
а) Электрические синапсы. Электрические синапсы в нервной системе млекопитающих встречаются редко. Они образованы щелевидными контактами (нексусами) между дендритами или сомами соприкасающихся нейронов, которые соединяются с помощью цитоплазматических каналов диаметром 1,5 нм. Процесс передачи сигнала происходит без синаптической задержки и без участия медиаторов.
Посредством электрических синапсов возможно распространение электротонических потенциалов от одного нейрона к другому. Вследствие тесного синаптического контакта модуляция проведения сигнала невозможна. Задача этих синапсов — осуществление одновременного возбуждения нейронов, выполняющих одинаковую функцию. Примером служат нейроны дыхательного центра продолговатого мозга, которые во время вдоха синхронно генерируют импульсы. Кроме того, примером могут служить нейронные цепи, управляющие саккадами, при которых точка фиксации взора перемещается от одного объекта внимания к другому.
б) Химические синапсы. Большинство синапсов нервной системы — химические. Функционирование таких синапсов зависит от высвобождения медиаторов. Классический химический синапс представлен пресинаптической мембраной, синаптической щелью и постсинаптической мембраной. Пресинаптическая мембрана — часть булавовидного расширения нервного окончания клетки, передающей сигнал, а постсинаптическая мембрана — часть клетки, получающей сигнал.
Медиатор высвобождается из булавовидного расширения посредством экзоцитоза, проходит через синаптическую щель и связывается с рецепторами на постсинаптической мембране. Под постсинаптической мембраной расположена субсинаптическая активная зона, в которой после активации рецепторов постсинаптической мембраны происходят разнообразные биохимические процессы.
В булавовидном расширении расположены содержащие медиаторы синаптические пузырьки, а также большое количество митохондрий и цистерны гладкой эндоплазматической сети. Применение традиционных методик фиксации при исследовании клеток позволяет различить на пресинаптической мембране пресинаптические уплотнения, ограничивающие активные зоны синапса, к которым при помощи микротрубочек направляются синаптические пузырьки.
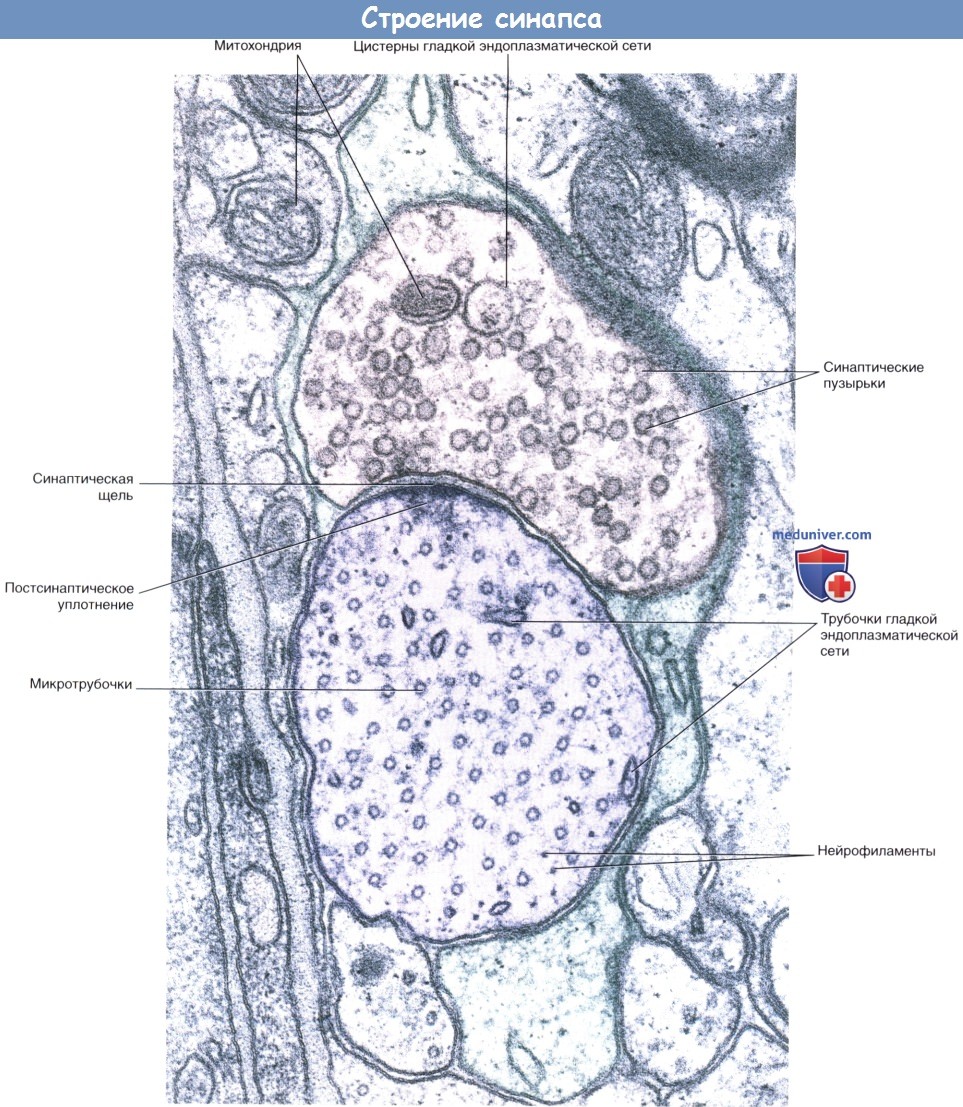 Аксодендритический синапс.
Аксодендритический синапс.
Срез препарата спинного мозга: синапс между концевым участком дендрита и, предположительно, двигательным нейроном.
Наличие округлых синаптических пузырьков и постсинаптического уплотнения характерно для возбуждающих синапсов.
Срез дендрита проведен в поперечном направлении, о чем свидетельствует наличие множества микротрубочек.
Кроме того, видны некоторые нейрофиламенты. Участок синапса окружен протоплазматическим астроцитом. 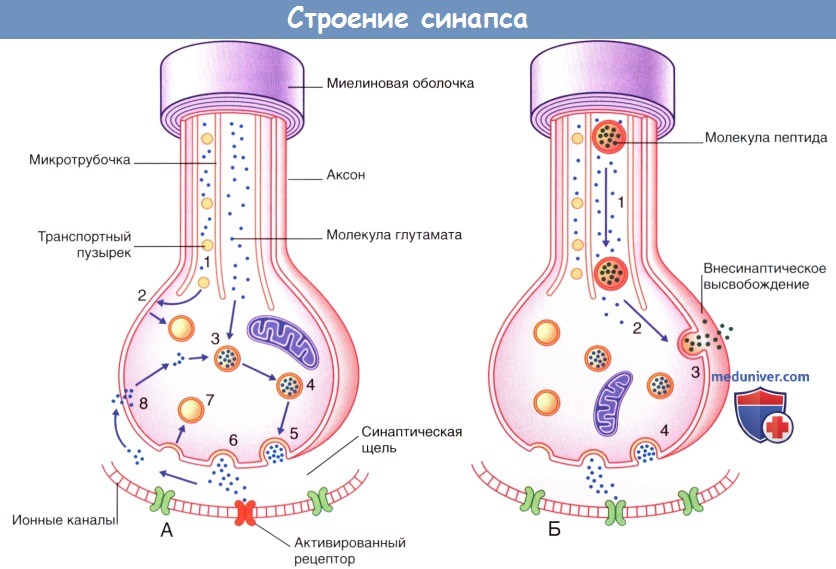 Процессы, происходящие в нервных окончаниях двух типов.
Процессы, происходящие в нервных окончаниях двух типов.
(А) Синаптическая передача небольших молекул (например, глутамата).
(1) Транспортные пузырьки, содержащие мембранные белки синаптических пузырьков, направляются вдоль микротрубочек к плазматической мембране булавовидного утолщения.
В это же время происходит перенос молекул ферментов и глутамата путем медленного транспорта.
(2) Мембранные белки пузырьков выходят из плазматической мембраны и формируют синаптические пузырьки.
(3) Глутамат погружается в синаптические пузырьки; происходит накопление медиатора.
(4) Пузырьки, содержащие глутамат, подходят к пресинаптической мембране.
(5) В результате деполяризации происходит экзоцитоз медиатора из частично разрушенных пузырьков.
(6) Высвобождающийся медиатор распространяется диффузно в области синаптической щели и активирует специфические рецепторы на постсинаптической мембране.
(7) Мембраны синаптических пузырьков транспортируются обратно в клетку путем эндоцитоза.
(8) Происходит частичный обратный захват глутамата в клетку для повторного использования.
(Б) Передача нейропептидов (например, субстанции Р), осуществляющаяся одновременно с синаптической передачей (например, глутамата).
Совместная передача этих веществ происходит в центральных нервных окончаниях униполярных нейронов, обеспечивающих болевую чувствительность.
(1) Синтезированные в комплексе Гольджи (в области перикариона) пузырьки и предшественники пептидов (пропептиды) транспортируются к булавовидному расширению путем быстрого транспорта.
(2) При их попадании в область булавовидного утолщения завершается процесс формирования молекулы пептида, и пузырьки транспортируются к плазматической мембране.
(3) Деполяризация мембраны и перенос содержимого пузырьков в межклеточное пространство путем экзоцитоза.
(4) Одновременно с этим происходит высвобождение глутамата.
1. Активация рецепторов. Молекулы медиаторов проходят через синаптическую щель и активируют рецепторные белки, расположенные парами на постсинаптической мембране. Активация рецепторов запускает ионные процессы, которые приводят к деполяризации постсинаптической мембраны (возбуждающее постсинаптическое действие) или гиперполяризации постсинаптической мембраны (тормозящее постсинаптическое действие). Изменение электротонуса передается в сому в виде затухающего по мере распространения электротонического потенциала, за счет которого происходит изменение потенциала покоя в начальном сегменте аксона.
Ионные процессы подробно описаны в отдельной статье на сайте. При преобладании возбуждающих постсинаптических потенциалов начальный сегмент аксона деполяризуется до порогового уровня и генерирует потенциал действия.
Наиболее распространенный возбуждающий медиатор ЦНС — глутамат, а тормозной — гамма-аминомасляная кислота (ГАМК). В периферической нервной системе медиатором для двигательных нейронов поперечно-полосатой мускулатуры служит ацетилхолин, а для чувствительных нейронов — глутамат.
Последовательность процессов, происходящих в глутаматергических синапсах, показана на рисунке ниже. При передаче глутамата совместно с другими пептидами высвобождение пептидов осуществляется внесинаптическим путем.
Большинство чувствительных нейронов помимо глутамата выделяет и другие пептиды (один или несколько), высвобождающиеся в различных участках нейрона; однако основная функция этих пептидов — модуляция (повышение или снижение) эффективности синаптической передачи глутамата.
Кроме того, нейротрансмиссия может происходить путем диффузной внесинаптической передачи сигнала, характерной для моноаминергических нейронов (нейронов, использующих биогенные амины для обеспечения нейротрансмиссии). Выделяют две разновидности моноаминергических нейронов. В одних нейронах осуществляется синтез катехоламинов (норадреналина или дофамина) из аминокислоты тирозина, а в других — серотонина из аминокислоты триптофана. Например, дофамин высвобождается как в синаптической области, так и из варикозных утолщений аксона, в которых также происходит синтез этого нейромедиатора.
Дофамин проникает в межклеточную жидкость ЦНС и до момента деградации способен активировать специфические рецепторы на расстоянии до 100 мкм. Моноаминергические нейроны присутствуют во многих структурах ЦНС; нарушение передачи импульса этими нейронами приводит к различным заболеваниям, среди которых выделяют болезнь Паркинсона, шизофрению и глубокую депрессию.
Оксид азота (газообразная молекула) также участвует в диффузной нейропередаче в глутаматергической системе нейронов. Избыточное влияние оксида азота оказывает цитотоксическое действие, особенно в тех участках, кровоснабжение которых нарушено за счет тромбоза артерий. Глутамат также является потенциально цитотоксическим нейромедиатором.
В отличие от диффузной нейротрансмиссии, традиционную синаптическую передачу сигнала ввиду ее относительной стабильности называют «проводниковой».
в) Резюме. Мультиполярные нейроны ЦНС состоят из сомы, дендритов и аксона; аксон образует коллатеральные и терминальные ветви. В соме расположены гладкая и шероховатая эндоплазматическая сети, комплексы Гольджи, нейрофиламенты и микротрубочки. Микротрубочки пронизывают нейрон на всем протяжении, принимают участие в процессе антероградного транспорта синаптических пузырьков, митохондрий и веществ для построения мембран, а также обеспечивают ретроградный транспорт «маркерных» молекул и разрушенных органелл.
Существует три вида химических межнейрональных взаимодействий: синаптическое (например, глутаматергическое), внесинаптическое (пептидергическое) и диффузное (например, моноаминергическое, серотонинергическое).
Химические синапсы классифицируют по анатомическому строению на аксодендритические, аксосоматические, аксоаксональные и дендро-дендритические. Синапс представлен пре- и постсинаптическими мембранами, синаптической щелью и субсинаптической активной зоной.
Электрические синапсы обеспечивают одновременную активацию целых групп нейронов, образуя между ними электрические связи за счет щелевидных контактов (нексусов).
![]() Диффузная нейротрансмиссия в головном мозге.
Диффузная нейротрансмиссия в головном мозге.
Аксоны глутаматергического (1) и дофаминергического (2) нейронов образуют плотные синаптические контакты с отростком звездчатого нейрона (3) полосатого тела.
Дофамин высвобождается не только из пресинаптической области, но и из варикозного утолщения аксона, откуда диффузно распространяется в межклеточное пространство и активирует дофаминовые рецепторы дендритного ствола и стенки перицита капилляра. 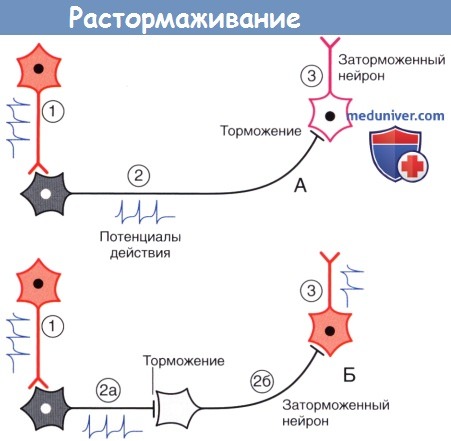 Растормаживание.
Растормаживание.
(А) Возбуждающий нейрон 1 активирует тормозной нейрон 2, который в свою очередь затормаживает нейрон 3.
(Б) Появление второго тормозного нейрона (2б) оказывает противоположное влияние на нейрон 3, поскольку происходит торможение нейрона 2б.
Спонтанно-активный нейрон 3 генерирует сигналы в условиях отсутствия тормозных влияний.
3. Торможение и растормаживание. Функционирование спонтанно-активных нейронов сдерживается под влиянием тормозных нейронов (обычно, ГАМКергических). Деятельность тормозных нейронов, в свою очередь, может быть ингибирована воздействующими на них другими тормозными нейронами, в результате чего происходит растормаживание клетки-мишени. Процесс растормаживания — важная особенность нейрональной активности в базальных ганглиях.
4. Редкие виды химических синапсов. Выделяют два типа аксоаксональных синапсов. В обоих случаях булавовидное утолщение образует тормозной нейрон. Синапсы первого типа образуются в области начального сегмента аксона и передают мощное ингибирующее влияние тормозного нейрона. Синапсы второго типа образуются между булавовидным утолщением тормозного нейрона и булавовидными утолщениями возбуждающих нейронов, что приводит к угнетению высвобождения медиаторов. Этот процесс получил название пресинаптического торможения. В этом плане традиционный синапс обеспечивает постсинаптичсекое торможение.
Дендро-дендритические (Д-Д) синапсы образуются между дендритными шипиками дендритов смежных шипиковых нейронов. Их задача — не генерирование нервного импульса, а изменение электротонуса клетки-мишени. В последовательных Д-Д-синапсах синаптические пузырьки располагаются только в одном дендритном шипике, а в реципрокном Д-Д-синапсе— в обоих. Возбуждающие Д-Д-синапсы изображены на рисунке ниже. Тормозные Д-Д-синапсы широко представлены в переключающих ядрах таламуса.
Кроме того, выделяют немногочисленные сомато-дендритические и сомато-соматические синапсы.
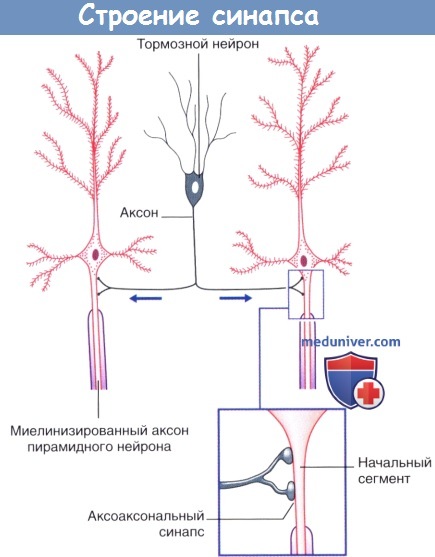 Аксоаксональные синапсы коры головного мозга.
Аксоаксональные синапсы коры головного мозга.
Стрелками указано направление проведения импульсов. 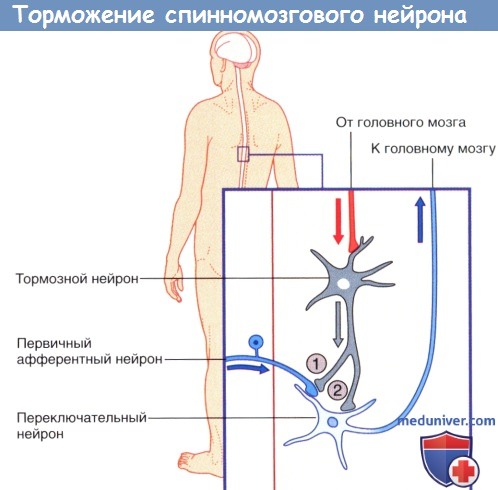 (1) Пресинаптическое и (2) постсинаптическое торможение спинномозгового нейрона, направляющегося к головному мозгу.
(1) Пресинаптическое и (2) постсинаптическое торможение спинномозгового нейрона, направляющегося к головному мозгу.
Стрелками указано направление проведения импульсов (возможно торможение переключательного нейрона под действием тормозных влияний). 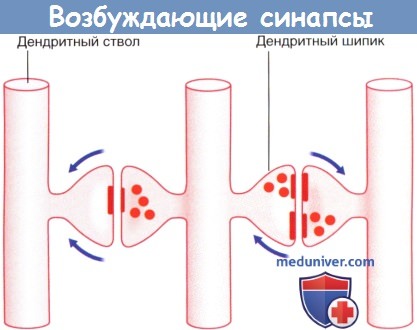 Возбуждающие дендро-дендритические синапсы. Изображены дендриты трех нейронов.
Возбуждающие дендро-дендритические синапсы. Изображены дендриты трех нейронов.
Реципрокный синапс (справа). Стрелками указано направление распространения электрото-нических волн.
Редактор: Искандер Милевски. Дата публикации: 11.11.2018
СИ́НАПС
Том 30. Москва, 2015, стр. 213-214
Скопировать библиографическую ссылку:

СИ́НАПС (от греч. σύναψις – соединение, связь), специализированные функциональные контакты между возбудимыми клетками (нервными, мышечными, секреторными), служащие для передачи и преобразования нервных импульсов. Термин «С.» ввёл Ч. Шеррингтон в 1897. Синаптич. связи (гл. механизм межнейронного взаимодействия) обеспечивают все осн. проявления деятельности нервной системы, составляют один из наиболее существенных, структурно-функциональных элементов мозга. В С. различают пресинаптич. часть (обычно окончание пресинаптич. аксона), синаптич. щель (пространство, разделяющее мембраны контактирующих клеток) и постсинаптич. часть (участок клетки, к которому подходит пресинаптич. окончание). Межнейронные С., образованные преим. окончаниями аксонов одних нейронов и телом, дендритами или аксонами других, соответственно называют аксосоматическими, аксодендритными (наиболее распространены) и аксо-аксонными; выделяют также дендродендритные, дендросоматические и соматосоматические С. Большинство нейронов образует тысячи синаптич. контактов, причём только один пресинаптич. аксон может устанавливать десятки контактов с индивидуальным постсинаптич. нейроном. По функциональному значению С. могут быть возбуждающими и тормозными в соответствии с тем, активируют они или подавляют деятельность соответствующей клетки.
Санкт-Петербургский государственный университет.Санкт-Петербург, Центр «Интеллект» Лисий нос. ЛО.
Биология в школе 2016, № 7, с. 3-12.
В статье приведены современные сведения о структуре, принципах работы и эволюции межнейронных контактов (синапсов) различного вида. Подробно разбирается морфология синапсов. Особое внимание уделено вопросам молекулярных механизмов работы синапсов и их эволюции в процессе становления и развития нервной системы у животных.
нервная система, нейрон, синапс, медиаторы, синаптические рецепторы
Таким образом, в конце ХХ – века окончательно сложилась « нейронная теория строения нервной системы », основными положениями которой стали тезисы о том, что нейрон – это элементарная единица нервной ткани и что нейроны соединяются друг с другом или с другими клетками организма посредством специальных межнейронных прерывистых контактов – синапсов .
Классифицировать синапсы можно исходя из разных их параметров и свойств.
— По способу передачи межнейронные контакты можно разделить на: химические, электрические и смешанные (электро-химические).
— По механизму действия на соседние нейроны синапсы разделяются на возбуждающие или тормозные ( см. ниже ).
Химические синапсы (строение, принцип работы, свойства). Этот вид межнейронных контактов широко представлен в нервной системе всех животных – от низших беспозвоночных до высших млекопитающих и человека. У высших животных (как позвоночных, так и беспозвоночных) они преобладают в нервной системе.
Принципиальная схема строения химического синапса едина: та часть нервной клетки (аксон, дендрит или участок тела нейрона), которая образует контакт, называется пресинапсом , затем идет синаптическая щель (шириной от 10 до 500 нм), а та часть, на которой оканчивается контакт – постсинапсом . (Рис. 1).

Рис.1 Электронная фотография химического синапса .
Обозначения: 1 – пресинапс, 2 – постсинапс, 3 – скопления синаптических пузырьков, 4 – пресинаптическое утолщение (активная зона синапса), 5 – постсинаптическое утолщение, 6 –митохондрия в пресинапсе. Масштаб 200 нм.
Основной принцип работы химического синапса – использование специальных химических веществ (медиаторов, трансмиттеров) для передачи сигнала от одного нейрона другому. Медиаторы находятся в пресинапсе в специальных мембранных структурах – синаптических пузырьках . Показано, что популяция синаптических пузырьков неоднородна. Большая часть их образует т.н. «резервный пул», который располагается в отдалении от пресинаптической мембраны и «рабочий пул», который используется в работе синапса в первую очередь. Пузырьки претерпевают в процессе работы синапса определенный цикл, по мере которого они используются в работе синапса, а затем вновь заполняются медиатором/ми. Также в пресинапсе находятся элементы цитоскелета и транспортной системы клетки (микротрубочки, актиновые филаменты, миозин), митохондрии, ферментные системы для синтеза медиатора. (Рис. 2).

Рис. 2 Схема строения химического синапса (аксо-шипиковый синапс).
Газообразные посредники имеют ряд свойств, отличающих их от классических медиаторов. Все они легко проникают через мембрану, выделяются из любого участка клетки, не запасаются в синаптических пузырьках и не освобождаются экзоцитозом и являются коротко живущими. Клеточные эффекты газов опосредуются либо через систему внутриклеточных посредников, либо через прямое влияние на субъединицы ионных каналов, белки экзоцитоза, внутриклеточные ферменты. В роли нейромедиаторов и нейромодуляторов газы имеют преимущества перед другими посредниками по скорости синтеза и выделения, степени проницаемости через мембрану и широкому спектру мишеней. Особенности действия газов позволяют предполагать их важную роль в процессах развития нервной системы, формировании кратковременных и долговременных изменений в синаптических структурах, связанных с процессами памяти и обучения.
При этом нейроны могут синтезировать и выделять в своих окончаниях целый набор медиаторов и комедиаторов (например, в ацетилхолиновых синапсах в качестве комедиаторов могут присутствовать: энкефалин, VIP, вещество P, соматостатин или нейротензин).
Вещества, которые претендуют на роль медиатора, должны соответствовать нескольким критериям:
— они должны синтезироваться нейроном и храниться в синапсах;
— при поступлении нервного импульса выделяться в синаптическую щель и избирательно связываться со специфическими рецепторами на постсинаптической мембране другого нейрона;
— вызывать соответствующую физиологическую реакцию;
— при введении в нервную систему извне оказывать такой же физиологический эффект, как и эндогенные медиаторы.
Синтез медиаторов идет в теле клетки, а затем с помощью транспортной системы нейрона пузырьки, заполненные медиатором (или пустые), поставляются по аксону в пресинапс. Часть медиатора синтезируется непосредственно в синапсе и заполняет синаптические пузырьки. Поскольку в одном синапсе может быть несколько медиаторов, то они могут находиться как в отдельных синаптических пузырьках, так и в одном (пузырьковая фракция). Кроме того часть медиатора/ов обнаруживается непосредственно в цитоплазме пресинапса (цитоплазматическая фракция). Обе эти фракции могут обмениваться друг с другом и участвовать в работе синапса. Количество медиатора, находящегося в одном пузырьке, получило наименование «квант». Например, в пузырьке ацетилхолинового синапса содержится примерно 10000 молекул ацетилхолина. Выброс медиатора с синаптическую щель происходит квантами, но для возникновения нервного импульса необходим выброс одновременно множества квантов.
Важно отметить, что термины «нейромедиатор», «нейромодулятор», «нейрогормон» отражают скорее механизм взаимодействия этих соединений с клетками-мишенями, чем их химическую природу. Одно и то же вещество может выступать как в роли медиатора, так и нейрогормона. Многие нейропептиды, например энкефалины и эндорфины, ранее рассматриваемые только как нейрогормоны, выделяются аксонными терминалями и выступают в роли нейромедиаторов. Другие действуют не только через межнейронные сипапсы, но и выделяются нейросекреторными и эндокринными клетками, выступая как типичные гормоны (адреналин, дофамин, серотонин и др.)
Важнейшей частью пресинапса является пресинаптическая мембрана с ее специализированными участками, названными активными зонами синапса , где происходит контакт синаптичеких пузырьков с мембраной и экзоцитоз медиатора в синаптическую щель. Активная зона не занимает всю площадь синаптического контакта и может меняться в зависимости от активности работы нейрона.
Со стороны пре- и постсинаптической мембраны в синаптическую щель выступают молекулы межклеточной адгезии (катенины, кадгерины, нейрексины, нейролигины и др.). Их роль состоит в связывании и закреплении пре- и постсинаптических мембран. Кроме того, в синаптической щели могут находиться ферменты, предназначенные для дезактивации медиатора/ров после выполнения ими своей функции.
Постсинапс является важнейшей частью синаптического контакта и включает несколько компонентов: собственно постсинаптическую мембрану со встроенными в нее постсинаптическими рецепторами и ионными каналами, субмембранный комплекс (постсинатическое утолщение ) и элементы цитоскелета постсинапса. Вся эта структура предназначена для стабилизации постсинаптических рецепторов и ионных каналов в мембране и формирования нового нервного импульса (постсинатическое возбуждение или торможение ).
Важно отметить, что знак работы синапса не зависит от химической природы медиатора, а связан со свойствами постсинаптических рецепторов. Этот важнейший факт был установлен автралийским ученым, лауреатом Нобелевской премии 1963 года Д. К. Экклзом.
Постсинаптические рецепторы являются сложными белковыми комплексами, встроенными в постсинаптическую мембрану. Выделяют три основных вида таких рецепторов: ионотропные, метаботроные и каталитические.
Ионотропные рецепторы представлены мембранными белковыми комплексами, состоящими, как правило, из 4-5 белковых субъединиц с ионным каналом в центре. При связывании медиатора с таким рецептором одновременно открывается ионный канал, происходит перераспределение ионных потоков внутри и снаружи мембраны, вследствие чего изменяется трансмембранный потенциал и возникает нервный импульс (постсинатический потенциал действия). (Рис 3, а).

Рис. 3 Схема строения ионотропного (А) и метаботропного (В) синаптических рецепторов, Б – субъединицы ионотропного никотинового ацетилхолинового (N-Ah) рецептора (объяснения в тексте).
Ионотропные рецепторы открываются почти мгновенно (время реакции
Метаботропные рецепторы представляют собой сложный молекулярный комплекс, где рецепторная часть и ионный канал пространственно разобщены в мембране постсинапса ( Рис. 3,в ). В связи с этим необходим « посредник/и, мессенджер » для связи их друг с другом и дальнейшей передачи сигнала на мембрану или вглубь клетки.
Метаботропные рецепторы сохраняют активированное состояние в течение секунд или минут после связывания с медиатором. Поэтому они имеют более длительные эффекты, чем ионотропные рецепторы.
Как и ионотропный никотиновый рецептор ( n AhR), метаботропный мускариновый ацетилхолиновый рецептор ( m AhR) имеет несколько разновидностей, различающихся по вторичным посредникам и типам ионных каналов ( Табл. 1).



