Хлоропласты
Хлоропласты являются одним из видов пластид. Хлоропласты имеют зеленый цвет за счет преобладающего в них пигмента хлорофилла. Основная их функция — фотосинтез.
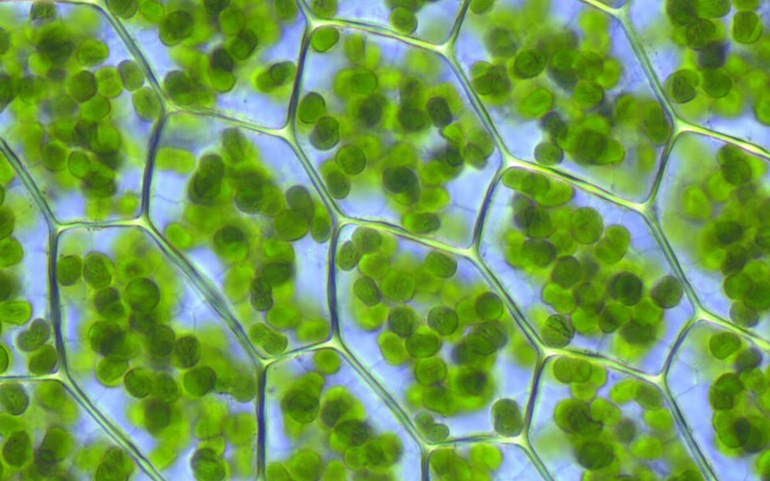
Количество данных органоидов в клетке варьирует. У некоторых водорослей в клетках содержится одни большой хлоропласт, часто причудливой формы. У высших растений их множество, особенно в мезофильной ткани листьев, где количество может достигать сотни штук на клетку.
У высших растений размер органоида около 5 мкм, форма округлая слегка вытянутая в одном направлении.
Хлоропласты в клетках развиваются из пропластид или путем деления надвое ранее существующих.
Строение хлоропласта
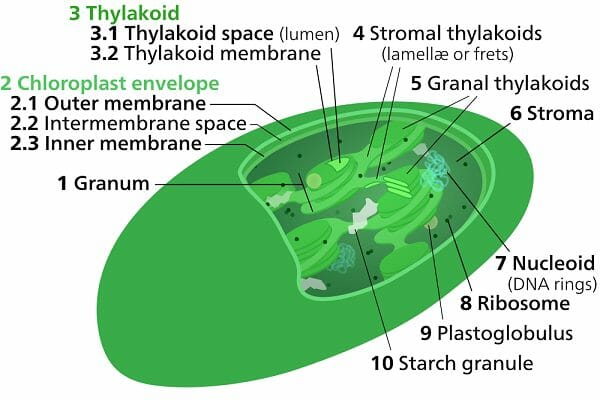
В строении хлоропластов выделяют внешнюю и внутреннюю мембраны, межмембранное пространство, строму, тилакоиды, граны, ламеллы, люмен.
Тилакоид представляет собой ограниченное мембраной пространство в форме приплюснутого диска. Тилакоиды в хлоропластах объединяются в стопки, которые называют гранами. Граны связаны между собой удлиненными тилакоидами — ламеллами.
Полужидкое содержимое хлоропласта называется стромой. В ней находятся его ДНК и РНК, рибосомы, обеспечивающие полуавтономность органоида (см. Симбиогенез).
Также в строме находятся зерна крахмала. Они образуются при избытке углеводов, образовавшихся при фотосинтетической активности. Жировые капли обычно формируются из мембран разрушающихся тилакоидов.
Функции хлоропластов
Основная функция хлоропластов — это фотосинтез — синтез глюкозы из углекислого газа и воды за счет солнечной энергии, которая улавливается хлорофиллом. В качестве побочного продукта фотосинтеза выделяется кислород. Однако процесс этот сложный и многоступенчатый, при котором синтезируются и побочные продукты, использующиеся как в самом хлоропласте, так и в остальных частях клетки.
Основным фотосинтетическим пигментом является хлорофилл. Он существует в нескольких разных формах. Кроме хлорофилла в фотосинтезе принимают участие пигменты каротиноиды.
Пигменты локализованы в мембранах тилакоидов, здесь протекают световые реакции фотосинтеза. Кроме пигментов здесь присутствуют ферменты и переносчики электронов. Хлоропласты стараются расположиться в клетке так, чтобы их тилакоидные мембраны находились под прямым углом к солнечному свету.
Хлорофилл состоит из длинного углеводного кольца и порфириновой головки. Хвост гидрофобен и погружен в липидный слой мембран тилакоидов. Головка гидрофильна и обращена к строме. Энергия света поглощается именно головкой, что приводит к возбуждению электронов.
Электрон отделяется от молекулы хлорофилла, который после этого становится электроположительным, т. е. оказывается в окисленной форме. Электрон принимается переносчиком, которые передает его на другое вещество.
Разные виды хлорофилла отличаются между собой несколько различным спектром поглощения солнечного света. Больше всего в растениях хлорофилла А.
В строме хлоропласта происходят темновые реакции фотосинтеза. Здесь находятся ферменты цикла Кальвина и другие.
Хлоропласты: роль в процессе фотосинтеза и структура

Фотосинтез происходит в эукариотических клеточных структурах, называемых хлоропластами. Хлоропласт – это тип органеллы растительных клеток, известный как зеленые пластиды. Пластиды помогают хранить и собирать необходимые вещества для производства энергии. Хлоропласт содержит зеленый пигмент, называемый хлорофиллом, который поглощает световую энергию для процесса фотосинтеза. Следовательно, название хлоропласт указывает на то, что эти органеллы представляют собой хлорофиллсодержащие пластиды.
Подобно митохондриям, хлоропласты имеют свою собственную ДНК, ответственны за производство энергии и воспроизводятся независимо от остальной части клетки посредством процесса деления, подобного бактериальному бинарному делению. Они также ответственны за производство аминокислот и липидных компонентов, необходимых для производства хлоропластов. Хлоропласты также встречаются в клетках других фотосинтезирующих организмах, таких как водоросли.
Хлоропласт: структура
Хлоропласты обычно встречаются в охранных клетках, расположенных в листьях растений. Охранные клетки окружают крошечные поры, называемые устьицами, открывая и закрывая их, чтобы обеспечить необходимый для фотосинтеза газообмен. Хлоропласты и другие пластиды развиваются из клеток, называемых пропластидами, которые являются незрелыми, недифференцированными клетками, развивающимися в разные типы пластид. Пропластид, развивающийся в хлоропласт, осуществляет этот процесс только при свете. Хлоропласты содержат несколько различных структур, каждая из которых имеет специализированные функции. Основные структуры хлоропласта включают:
Хлоропласт: фотосинтез

При фотосинтезе энергия солнечного света преобразуется в химическую энергию. Химическая энергия хранится в виде глюкозы (сахара). Двуокись углерода, вода и солнечный свет используются для производства глюкозы, кислорода и воды. Фотосинтез происходит в два этапа: световая фаза и темновая фаза.
Световая фаза фотосинтеза протекает только при наличии света и происходит внутри хлоропластовой граны. Первичным пигментом, используемым для преобразования световой энергии в химическую, является хлорофилл а. Другие пигменты, участвующие в поглощении света, включают хлорофилл b, ксантофилл и каротин. Во время световой фазы, солнечный свет преобразуется в химическую энергию в виде АТФ (молекулы, содержащей свободную энергию) и НАДФ (молекула, несущая электроны высокой энергии).
И АТФ, и НАДФ используются во время темновой фазы для получения сахара. Темновая фаза фотосинтеза, также известная как этап фиксации углерода или цикл Кальвина. Реакции на этой стадии возникают в строме. Строма содержит ферменты, которые облегчают серию реакций, использующих АТФ, НАДФ и углекислый газ для получения сахара. Сахар может храниться в виде крахмала, используемого во время дыхания или при производстве целлюлозы.
Функциональные особенности
Строение хлоропласта изучается школьниками в 6 классе на уроках биологии. К особенностям клеток относится наличие в строме рибосомы, ДНК, РНК. В мембране присутствует вещество, способное придать растениям соответствующий цвет. Для хлорофилла характерен зеленый оттенок, а для каротиноида:

Значение хлорофилла для растений заключается в возможности осуществления процесса фотосинтеза. С учётом строения биологи выделяют 4 типа хлорофилла: a, b, c, d. Первые два содержатся в растениях на суше и зеленых водорослях. Типы a и c считаются растительными компонентами диатомовых, d и a — красных водорослей.
Для хлорофилла характерно поглощение солнечной энергии с последующей передачей иным молекулам. Разрушение зеленого вещества наблюдается в конце жизненного цикла органоида в результате резкого изменения светового дня и значения температуры. Часть хлоропластов превращается в хромопласты. Это приводит к изменению внутренней информации, пожелтению и опадению листьев.
Принципы классификации
Пластиды делятся на три вида: лейкопласты (бесцветные), хлоропласты (окрашенные в зеленый цвет), хромопласты (имеют разные оттенки). На протяжении жизни клетки способны превращаться друг в друга. Лейкопластам свойственно переходить в хлоропласты, а последние за счёт появления бурых и прочих пигментов — в хромопласты, пластоглобулы.
Внешне зеленые вещества покрыты липидной и белковой мембранами. Полужидкая строма с тилакоидами (компартменты, ограниченные мембраной) считается основным веществом, в состав которого входят граны с каналами. Первые компоненты представлены в виде плоских круглых мешочков, расположенных перпендикулярно поверхности двухмембранных органоидов (ДО).
Уникальность их структуры заключается в хранении зеленого пигмента (хлорофилл). Главная функция хлоропластов связана с участием в фотосинтетическом явлении. В их состав входят жиры, зерна (митохондрия, пропластида), крахмал.
На долю липидов приходится до 30%. Они представлены тремя группами:
К другим компонентам, входящим в состав хлоропласта, относятся углеводы. Они представлены в виде продуктов фотосинтеза. До 25% приходится на долю минералов. Ферменты могут выполнять двойную функцию: катализацию различных реакций, обеспечение биосинтеза белков.
Внутренняя структурированность хлоропластов зависит от функциональных нагрузок, физиологического состояния. Молодые клетки размножаются за счет деления, а зрелые обладают выраженной системой гран. Если они стареют, происходит разрыв тилакоидов, распадается хлорофилл. Осенью деградация приводит к появлению хромопластов.
Главная роль хлоропластов в фотосинтезе обеспечена их способностью пассивно двигаться в клетках, увлекаемых током цитоплазмы. Веществу свойственно собирать свет и активно перемещаться с одного места на другое. При интенсивном свете оно поворачивается ребром к яркому солнцу, выстраиваясь вдоль стенок, которые параллельны лучам.
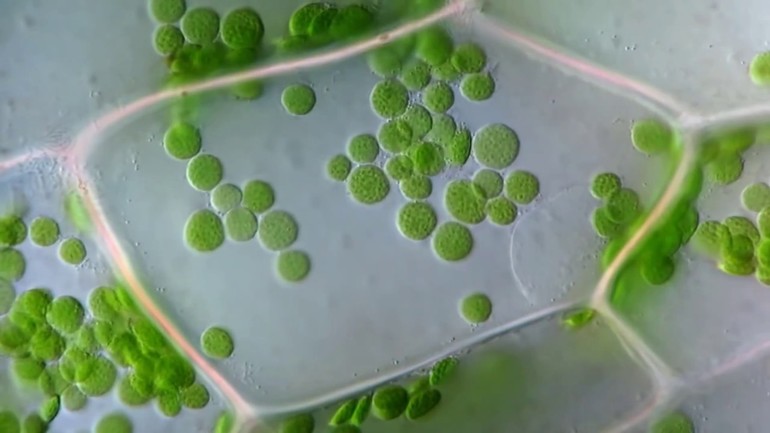
Если освещение слабое, схема движения хлоропластов следующая: они перемещаются на стенки, обращённые к солнцу, поворачиваясь наибольшей поверхностью. Когда освещение среднее, клетки занимают соответствующее положение. От условий освещения зависит то, какие пигменты хлоропластов появятся.
Для пластид и митохондрий свойственна полуавтономная степень. Кроме фотосинтеза, в первых компонентах происходит биосинтез белка. Так как они содержат в себе ДНК, поэтому принимают активное участие в наследственном комплексе: передача признаков, цитоплазматические свойства.
Описание хромопластов
К пластидам высших растений относятся хромопласты. Они имеют незначительные размеры. Для внутриклеточных органелл характерен разный окрас: красный, желтый, коричневый. Он придает соответствующий цвет осенью, плодам и цветкам, что необходимо для привлечения опылителей и животных, разносящих семена продолжительные расстояния.
Структура ткани похожа на иные пластиды. Внутренняя оболочка развита слабее внешней. У некоторых представителей она может отсутствовать. В каротиноидах (жирорастворимые пигменты) происходит накапливание кристаллов. Для определения точных функций вещества изучается таблица с формами хромопластов:

Их роль в жизни растений до конца не выяснена. Ученые предполагают, что пигменты участвуют в окислительных и восстановительных процессах, необходимых для размножения и физиологического развития клеток.
Строение лейкопластов
В органоидах этого типа накапливаются питательные компоненты. Лейкопласты имеют 2 оболочки: внутреннюю и внешнюю. На свету им свойственно превращаться в хлоропласты, но в привычном состоянии органоиды бесцветны. Основная их форма — шаровидная. Размещены они в мягких частях растений:
С учетом накапливаемого вещества лейкопласты классифицируются на следующие виды: амилопласты, элайопласты, протеинопласты. В первую группу входят органоиды с крахмалом, находящиеся в каждом растении. Если лейкопласт полностью заполнен крахмалом, он называется крахмальным зерном. Для элайопластов характерно продуцирование и запас жиров, а для протеинопластов — скопление белковых веществ.
Лейкопласты обладают ферментной субстанцией, что способствует ускоренному протеканию химических реакций. В отрицательном жизненном периоде, когда не происходит фотосинтез, они расщепляют полисахариды на простые углеводы. Так как в луковицах содержится много органоидов, поэтому им свойственно переносить длительную засуху, жару, низкую температуру. После выполнения своих функций они становятся хромопластами.
Симбиотическая теория
Чтобы выяснить механизм появления пластид, митохондрий и других органоидов, рассматривается теория эндосимбиоза. Ее суть заключается в совместной и взаимовыгодной жизни органеллы с клеткой. Впервые теорию предложил Шимпер в 1883 году. В 1867 ученые работали над двойственной природой лишайников.
Биолог Фамицын, учитывая теорию Шимпера, предположил, что хлоропласты, как лишайники и водоросли, относятся к симбионтам. Ученые доказали, что митохондрии — аэробные бактерии, которые не размножаются за пределами клеток. Общие свойства, характерные для митохондрий и пластид:
В ДНК пластид и митохондрий, в отличие от аналогичных структур прокариот, нет интронов. А в ДНК хлоропластов закодирована информация о некоторых белках, остальные данные находятся в ядре клетки. В результате эволюции часть генетического материала из генома перешло в ядро, поэтому хлоропласты и митохондрии не размножаются независимо.
Археи и бактерии не склонны к фагоцитозу. Они питаются только осмотрофно. Множественные биологические и химические исследования указывают на химерную сущность бактерий. Ученые не выяснили, как сливаются организмы из нескольких доменов. В условиях современности выявлены организмы, которые содержат в себе другие клетки в качестве эндосимбионтов. Они отличаются от первичных эукариотов тем, что не интегрируются в одно целое, не имеют своей индивидуальности.
Интересным организмом считается Mixotricha paradoxa. Чтобы двигаться, она использует 250 000 бактерий, которые фиксируются на ее поверхности. Митохондрии у этого организма вторично потеряны. Внутри находятся сферические аэробные микроорганизмы, которые заменяют органеллы.
Строма – определение и функция
Определение стромы
Строма обычно относится к заполненному жидкостью внутреннему пространству хлоропластов, окружающих тилакоиды и граны. Первоначально считалось, что строма просто обеспечивает поддержку пигментированных тилакоидов. Однако теперь известно, что строма содержит крахмал, хлоропласт ДНК и рибосомы, а также все ферменты, необходимые для светозависимых реакций фотосинтез также известный как Цикл Кальвина.
Происходя от греческого слова для слоя или покрытия кровати, строма может также относиться к другим поддерживающим структурам, таким как соединительные ткани в органах или грибке ткань это несет споры.
Строение хлоропласта строма
Микроскопическое исследование хлоропласта выявляет некоторые очевидные особенности. Он состоит из внешней мембраны и сложной сети внутренних мембран, образующих стопки дискообразных структур, называемых гранами. Различные граны связаны друг с другом через мембранные отростки.
Хлоропласты произошли от свободно живущих прокариот, которые сформировали эндосимбиотические отношения с некоторыми эукариотическими клетками. Следовательно, строма продолжает содержать ДНК и рибосомы для осуществления синтеза белка. Эти белки включают те, которые важны в светозависимых реакциях фотосинтеза, а также в реакциях, которые фиксируют неорганические минералы, такие как нитраты, в органических молекулах.
Функция Хлоропласт Строма
Функция в фотосинтезе
Строма сначала начинает играть роль в фотосинтезе, когда энергия света, захваченная молекулами пигмента, преобразуется в химическую энергию через цепь переноса электронов.

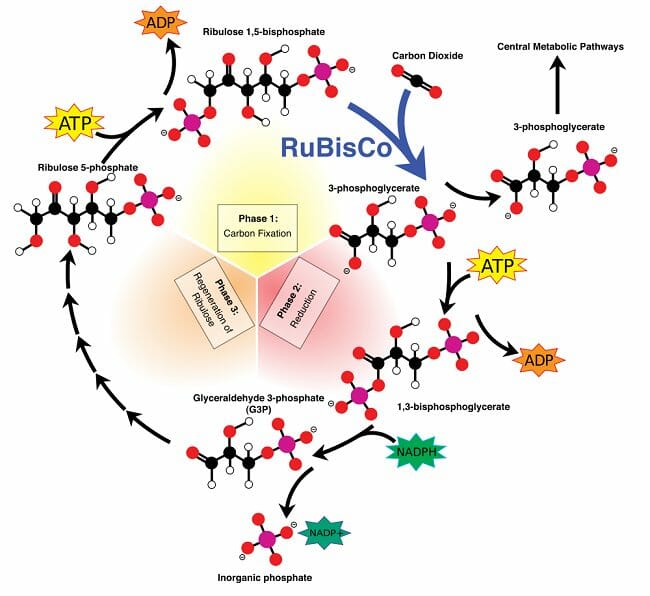
Наиболее важным ферментом в реакциях, не зависящих от света, или цикла Кальвина, является RuBisCO или рибулозо-1,5-бисфосфат (RuBP) карбоксилаза. Этот фермент катализирует первую стадию светозависимых реакций, включающих фиксацию углерода. RuBisCO улавливает двуокись углерода в атмосфере, которая диффундирует в строму хлоропласта, и фиксирует ее в форме органического вещества. молекула, Каждая молекула СО2 объединяется с одной молекулой RuBP, содержащей пять атомов углерода, в результате чего образуются две молекулы фосфоглицерата, которые являются трехуглеродными молекулами.
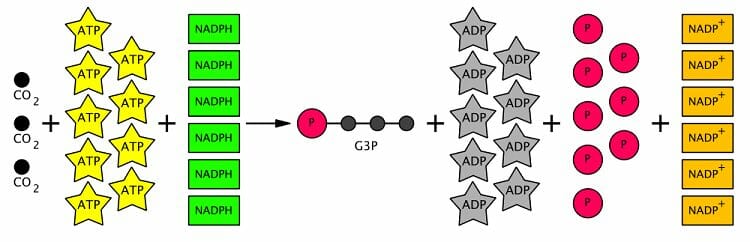
Цикл Кальвина имеет еще две ступени, которые происходят в строме – восстановление фосфоглицерата и регенерация RuBP. Эти шаги включают использование ATP и NADPH. В целом, свето-независимые реакции используют две молекулы НАДФН и три молекулы АТФ, чтобы зафиксировать одну молекулу атмосферного CO2.
Функция в внутриорганной сигнализации
Хлоропласты являются полуавтономными, поскольку они содержат свой собственный геном, но также импортируют ряд белков и небольших молекул из цитоплазма клетки. Хотя изначально они были свободноживущими автотрофами, с течением времени некоторые их гены были перенесены в ядро хозяина. Эти гены слегка модифицированы, так что белки нацелены на хлоропласт и, по-видимому, находятся под комбинированной регуляцией как ядра, так и хлоропласта. Передача сигналов от ядра к пластиде называется антероградной передачей сигналов, а сигналы, идущие к ядру, называются ретроградными сигналами. Оба этих сигнала, по-видимому, опосредованы через стромулы, которые также играют роль в коммуникации между двумя пластидами.
Calvin Cycle: свет-независимые реакции
Строма является местом для трех этапов цикла Кальвина – фиксации, восстановления и регенерации углерода.
Фиксация углерода начинается с реакции между одной молекулой CO2 и одной молекулой RuBP. Эти шесть атомов углерода и две фосфатные группы собираются вместе, образуя две молекулы фосфоглицерата, трехуглеродную молекулу, содержащую один фосфатная группа, Эту реакцию повторяют трижды, чтобы получить шесть молекул фосфоглицерата.
На следующем этапе фосфоглицерат принимает электроны с образованием глицеральдегид-3-фосфата (G3P). Движущей силой этой реакции восстановления является преобразование NADPH в NADP + и ATP в ADP. Таким образом, ADP и NADP + регенерируются для использования в светозависимых реакциях.
Это оставляет один последний шаг – регенерация RuBP. Из шести молекул G3P, полученных на предыдущем этапе, пять используются в реформировании RuBP, а шестая экспортируется из хлоропласта для получения глюкозы.
Примеры животных Строма
В то время как строма в каждой ткани или органе выполняет некоторые общие функции, такие как транспортировка топлива и метаболитов, а также структурная поддержка, в некоторых органах они выполняют специфические функции. Строма в эндокринные железы поддерживают образование гормонов в фолликулах и долях органа. В тимусе строма влияет на дифференцировку Т-клеток посредством положительного или отрицательного отбора. Органы, которые должны быстро реагировать на меняющиеся требования организм, такие как костный мозг или радужная оболочка глаза, также нужна специализированная строма.
Строма костного мозга
Строма костного мозга не участвует напрямую в кроветворение, но создает микросреду, которая усиливает активность клеток, участвующих в формировании кровь, Строма продуцирует факторы роста, содержит клетки, участвующие в метаболизме костей, содержит жировые клетки, а также макрофаги. Макрофаги особенно важны, потому что они участвуют в обмене эритроцитов и обеспечивают железо, необходимое для производства гемоглобина.
Строма ириса
Человеческая радужка начинает формироваться в первом триместре беременности и является одним из немногих внутренних органов тела, которые можно легко наблюдать. Радужная оболочка состоит из пигментированного эпителия вместе с мышцами, необходимыми для сжатия или расширения зрачка. Эти клетки выполняют основную функцию радужной оболочки и поддерживаются сосудистой стромой с высоким содержанием сосудов, рыхлым и прерывистым слоем соединительной ткани, содержащим связки и пигментообразующие клетки. Наличие пигмента экранирует падающий на глаз свет и позволяет лишь некоторым из них проходить через зрачок, образуя изображение на сетчатке. Эта пигментация определяется плотностью и наличием меланина глубоко в строме с коричневым глаза возникающие в результате сильной пигментации, и люди, имеющие синие ирисы, производят очень мало пигмента.
викторина
1. Из чего сделаны граны в хлоропластах?A. Внешние мембраносвязанные белкиB. Стеки внутренних мембранных структурC. хлорофиллD. Все вышеперечисленное
Ответ на вопрос № 1
В верно. Грана состоит из стеков тилакоидов, которые образуются при инвагинациях внутренней мембраны в хлоропластах. В то время как хлорофилл существует как часть фотосистем, использующих световую энергию, грана – это больше, чем просто совокупность пигментов.
2. Что из этого НЕ является функцией стромы хлоропласта?A. Реакция на стрессB. Внутриклеточная сигнализацияC. Высвобождение электронов высоких энергий при взаимодействии с фотонамиD. Все вышеперечисленное
Ответ на вопрос № 2
С верно. Выделение электронов высоких энергий является чисто функцией реакционных центров в тилакоидах. Хотя строма участвует в обеспечении протонов во время цепи переноса электронов, они не участвуют напрямую в высвобождении электронов высокой энергии. Строма была вовлечена как в реакцию на стресс, так и во внутриклеточную передачу сигналов посредством образования пальцевидных выпячиваний.
3. Что из этого НЕ является функцией стромы в тканях животных?A. Структурная поддержкаB. Кислородный транспортC. Секреция факторов ростаD. Ни один из вышеперечисленных
Ответ на вопрос № 3
D верно. Строма в тканях животных поддерживает орган различными способами, включая обеспечение структурной поддержки, транспортировку кислорода через кровеносные сосуды и удаление продуктов метаболизма. Часто они также выделяют цитокины и другие факторы роста. Следовательно, все три варианта являются функциями стромы в тканях животных.
Хлоропласт


Слово хлоропласт происходит от греческих слов хлорос (χλωρός), что означает зеленый, и пластов (πλάστης), что означает «тот, кто формирует». [4]
Родительская группа: Цианобактерии
Первичный эндосимбиоз

Глаукофита
Rhodophyceae (красные водоросли)
Хлоропластида (зеленые водоросли и растения)

Большинство хлоропластов, изображенных в этой статье, являются зелеными хлоропластами.
Паулинелла хроматофора
Вторичный и третичный эндосимбиоз
Возможная кладограмма эволюции хлоропластов [18] [19] [34]
Круги представляют собой эндосимбиотические события. Для ясности опущены третичные эндосимбиозы динофитов и многие нефотосинтетические клоны.


Хлоропласты, полученные из зеленых водорослей
Эвгленофиты
Хлорарахниофиты
Динофитный хлоропласт, производный от празинофитов
Хлоропласты, полученные из красных водорослей
Криптофиты
Криптофиты, возможно, сыграли ключевую роль в распространении хлоропластов на основе красных водорослей. [49] [50]
Гаптофиты
Heterokontophytes (stramenopiles)
Apicomplexans, хромериды и динофиты
Альвеолаты являются основной кладой одноклеточных эукариот как автотрофных, так и гетеротрофных членов. Наиболее заметной общей характеристикой является наличие кортикальных (внешних) альвеол (мешочков). Это сплюснутые пузырьки (мешочки), упакованные в непрерывный слой непосредственно под мембраной и поддерживающие ее, обычно образующие гибкую пленку (тонкую кожу). У динофлагеллят они часто образуют панцирные пластины. Многие члены содержат пластиду, полученную из красных водорослей. Одной из примечательных характеристик этой разнообразной группы является частая потеря фотосинтеза. Однако большинство этих гетеротрофов продолжают обрабатывать нефотосинтетические пластиды. [51]
Хлоропласт перидинина, как полагают, является «исходным» хлоропластом динофитов [48], который был утерян, уменьшен, заменен или имеет компанию в нескольких других линиях динофитов. [34]
Фукоксантин-содержащие (происходящие от гаптофита) хлоропласты динофитов
Хлоропласты, содержащие фукоксантин, характеризуются наличием пигмента фукоксантина (на самом деле 19′-гексаноилоксифукоксантин и / или 19′-бутаноилоксифукоксантина ) и отсутствием перидинина. Фукоксантин также обнаружен в хлоропластах гаптофитов, что свидетельствует о его происхождении. [48]
Хлоропласты диатомовых водорослей
Клептопластиды
Динофитный хлоропласт, полученный из криптофитов
Молекулярная структура
![]()
Хотя обычно их считают круговой молекулой, есть некоторые свидетельства того, что молекулы хлоропластной ДНК чаще принимают линейную форму. [64] [66]
Инвертированные повторы
Инвертированные повторы сильно различаются по длине, от 4000 до 25000 пар оснований каждый и содержат от четырех до более 150 генов. [64] Инвертированные повторы у растений, как правило, находятся в верхнем конце этого диапазона, каждый из которых имеет длину 20 000–25 000 пар оснований. [44] [67]
Области инвертированных повторов высоко консервативны среди наземных растений и накапливают мало мутаций. [44] [67] Подобные инвертированные повторы существуют в геномах цианобактерий и двух других клонов хлоропластов ( glaucophyta и rhodophyceae ), что позволяет предположить, что они предшествовали хлоропласту, [64] хотя некоторые ДНК хлоропластов с тех пор утрачены [67] [68] или перевернул перевернутые повторы (сделав их прямыми повторами). [64] Возможно, что инвертированные повторы помогают стабилизировать остальную часть генома хлоропластов, поскольку ДНК хлоропластов, которые потеряли некоторые из сегментов инвертированных повторов, имеют тенденцию больше перестраиваться. [68]
Нуклеоиды
Ремонт ДНК
Ведущая модель репликации хпДНК

Механизм репликации хлоропластной ДНК (хпДНК) окончательно не определен, но были предложены две основные модели. Ученые пытались наблюдать репликацию хлоропластов с помощью электронной микроскопии с 1970-х годов. [73] [74] Результаты экспериментов с микроскопией привели к идее, что ДНК хлоропластов реплицируется с использованием петли двойного смещения (D-петля). По мере того, как D-петля движется по кольцевой ДНК, она принимает промежуточную форму тета, также известную как промежуточное звено репликации Кэрнса, и завершает репликацию с помощью механизма катящегося круга. [73] [66] Транскрипция начинается в определенных точках происхождения. Открываются множественные репликационные вилки, позволяя репликационному аппарату транскрибировать ДНК. По мере продолжения репликации вилки разрастаются и в конечном итоге сходятся. Новые структуры хпДНК разделяются, образуя дочерние хромосомы хпДНК.

Дезаминирование
В хпДНК существует несколько градиентов дезаминирования A → G. ДНК становится восприимчивой к событиям дезаминирования, когда она является одноцепочечной. Когда образуются репликационные вилки, не копируемая нить является однонитевой и, следовательно, подвержена риску дезаминирования A → G. Следовательно, градиенты дезаминирования указывают на то, что вилки репликации, скорее всего, присутствовали, и направление, в котором они изначально открывались (самый высокий градиент, скорее всего, находится ближе всего к месту старта, потому что он был одноцепочечным в течение самого длительного периода времени). [73] Этот механизм до сих пор является ведущей теорией; однако вторая теория предполагает, что большая часть хпДНК на самом деле линейна и реплицируется посредством гомологичной рекомбинации. Далее он утверждает, что только меньшая часть генетического материала хранится в кольцевых хромосомах, тогда как остальная часть находится в разветвленных, линейных или других сложных структурах. [73] [66]
Альтернативная модель репликации
Содержание генов и синтез белка
Среди наземных растений состав генома хлоропластов довольно схож. [44]
Редукция генома хлоропласта и перенос генов
Синтез белка
Нацеливание на белок и импорт

В большинстве, но не во всех случаях, белки хлоропластов, кодируемые ядром, транслируются с помощью расщепляемого транзитного пептида, который добавляется к N-концу предшественника белка. Иногда транзитная последовательность обнаруживается на С-конце белка [87] или в функциональной части белка. [85]
Транспортные белки и мембранные транслоконы



Наружная хлоропластная мембрана
Межмембранное пространство и пептидогликановая стенка
Внутренняя мембрана хлоропласта
Периферический ретикулум
Строма
Рибосомы хлоропластов

Пластоглобулы
Гранулы крахмала
RuBisCO

Пиреноиды
Thylakoid system
Thylakoids (sometimes spelled thylakoïds), [125] are small interconnected sacks which contain the membranes that the light reactions of photosynthesis take place on. The word thylakoid comes from the Greek word thylakos which means «sack». [126]
Suspended within the chloroplast stroma is the thylakoid system, a highly dynamic collection of membranous sacks called thylakoids where chlorophyll is found and the light reactions of photosynthesis happen. [15] In most vascular plant chloroplasts, the thylakoids are arranged in stacks called grana, [127] though in certain C4 plant chloroplasts [119] and some algal chloroplasts, the thylakoids are free floating. [16]
Thylakoid structure
Using a light microscope, it is just barely possible to see tiny green granules—which were named grana. [106] With electron microscopy, it became possible to see the thylakoid system in more detail, revealing it to consist of stacks of flat thylakoids which made up the grana, and long interconnecting stromal thylakoids which linked different grana. [106] In the transmission electron microscope, thylakoid membranes appear as alternating light-and-dark bands, 8.5 nanometers thick. [106]
For a long time, the three-dimensional structure of the thylakoid membrane system had been unknown or disputed. Many models have been proposed, the most prevalent being the helical model, in which granum stacks of thylakoids are wrapped by helical stromal thylakoids. [128] Another model known as the ‘bifurcation model’, which was based on the first electron tomography study of plant thylakoid membranes, depicts the stromal membranes as wide lamellar sheets perpendicular to the grana columns which bifurcates into multiple parallel discs forming the granum-stroma assembly. [129] The helical model was supported by several additional works, [127] [130] but ultimately it was determined in 2019 that features from both the helical and bifurcation models are consolidated by newly-discovered left-handed helical membrane junctions. [124] Likely for ease, the thylakoid system is still commonly depicted by older «hub and spoke» models where the grana are connected to each other by tubes of stromal thylakoids. [131]
Grana consist of a stacks of flattened circular granal thylakoids that resemble pancakes. Each granum can contain anywhere from two to a hundred thylakoids, [106] though grana with 10–20 thylakoids are most common. [127] Wrapped around the grana are multiple parallel right-handed helical stromal thylakoids, also known as frets or lamellar thylakoids. The helices ascend at an angle of
20°, connecting to each granal thylakoid at a bridge-like slit junction. [127] [130] [124]
The stroma lamellae extend as large sheets perpendicular to the grana columns. These sheets are connected to the right-handed helices either directly or through bifurcations that form left-handed helical membrane surfaces. [124] The left-handed helical surfaces have a similar tilt angle to the right-handed helices (
20°), but ¼ the pitch. Approximately 4 left-handed helical junctions are present per granum, resulting in a pitch-balanced array of right- and left-handed helical membrane surfaces of different radii and pitch that consolidate the network with minimal surface and bending energies. [124] While different parts of the thylakoid system contain different membrane proteins, the thylakoid membranes are continuous and the thylakoid space they enclose form a single continuous labyrinth. [127]
Thylakoid composition
Embedded in the thylakoid membranes are important protein complexes which carry out the light reactions of photosynthesis. Photosystem II and photosystem I contain light-harvesting complexes with chlorophyll and carotenoids that absorb light energy and use it to energize electrons. Molecules in the thylakoid membrane use the energized electrons to pump hydrogen ions into the thylakoid space, decreasing the pH and turning it acidic. ATP synthase is a large protein complex that harnesses the concentration gradient of the hydrogen ions in the thylakoid space to generate ATP energy as the hydrogen ions flow back out into the stroma—much like a dam turbine. [99]
There are two types of thylakoids—granal thylakoids, which are arranged in grana, and stromal thylakoids, which are in contact with the stroma. Granal thylakoids are pancake-shaped circular disks about 300–600 nanometers in diameter. Stromal thylakoids are helicoid sheets that spiral around grana. [127] The flat tops and bottoms of granal thylakoids contain only the relatively flat photosystem II protein complex. This allows them to stack tightly, forming grana with many layers of tightly appressed membrane, called granal membrane, increasing stability and surface area for light capture. [127]
In contrast, photosystem I and ATP synthase are large protein complexes which jut out into the stroma. They can’t fit in the appressed granal membranes, and so are found in the stromal thylakoid membrane—the edges of the granal thylakoid disks and the stromal thylakoids. These large protein complexes may act as spacers between the sheets of stromal thylakoids. [127]
The number of thylakoids and the total thylakoid area of a chloroplast is influenced by light exposure. Shaded chloroplasts contain larger and more grana with more thylakoid membrane area than chloroplasts exposed to bright light, which have smaller and fewer grana and less thylakoid area. Thylakoid extent can change within minutes of light exposure or removal. [108]
Pigments and chloroplast colors
Inside the photosystems embedded in chloroplast thylakoid membranes are various photosynthetic pigments, which absorb and transfer light energy. The types of pigments found are different in various groups of chloroplasts, and are responsible for a wide variety of chloroplast colorations.
Chlorophylls
Chlorophyll a is found in all chloroplasts, as well as their cyanobacterial ancestors. Chlorophyll a is a blue-green pigment [132] partially responsible for giving most cyanobacteria and chloroplasts their color. Other forms of chlorophyll exist, such as the accessory pigments chlorophyll b, chlorophyll c, chlorophyll d, [16] and chlorophyll f.
Chlorophyll b is an olive green pigment found only in the chloroplasts of plants, green algae, any secondary chloroplasts obtained through the secondary endosymbiosis of a green alga, and a few cyanobacteria. [16] It is the chlorophylls a and b together that make most plant and green algal chloroplasts green. [132]
Chlorophyll c is mainly found in secondary endosymbiotic chloroplasts that originated from a red alga, although it is not found in chloroplasts of red algae themselves. Chlorophyll c is also found in some green algae and cyanobacteria. [16]
Chlorophylls d and f are pigments found only in some cyanobacteria. [16] [133]
Carotenoids
In addition to chlorophylls, another group of yellow–orange [132] pigments called carotenoids are also found in the photosystems. There are about thirty photosynthetic carotenoids. [134] They help transfer and dissipate excess energy, [16] and their bright colors sometimes override the chlorophyll green, like during the fall, when the leaves of some land plants change color. [135] β-carotene is a bright red-orange carotenoid found in nearly all chloroplasts, like chlorophyll a. [16] Xanthophylls, especially the orange-red zeaxanthin, are also common. [134] Many other forms of carotenoids exist that are only found in certain groups of chloroplasts. [16]
Phycobilins
Phycobilins are a third group of pigments found in cyanobacteria, and glaucophyte, red algal, and cryptophyte chloroplasts. [16] [136] Phycobilins come in all colors, though phycoerytherin is one of the pigments that makes many red algae red. [137] Phycobilins often organize into relatively large protein complexes about 40 nanometers across called phycobilisomes. [16] Like photosystem I and ATP synthase, phycobilisomes jut into the stroma, preventing thylakoid stacking in red algal chloroplasts. [16] Cryptophyte chloroplasts and some cyanobacteria don’t have their phycobilin pigments organized into phycobilisomes, and keep them in their thylakoid space instead. [16]
| Photosynthetic pigments Table of the presence of various pigments across chloroplast groups. Colored cells represent pigment presence. [16] [134] [136] | |||||||||
| Chlorophyll a | Chlorophyll b | Chlorophyll c | Chlorophyll d and f | Xanthophylls | α-carotene | β-carotene | Phycobilins | ||
| Land plants | |||||||||
| Green algae | |||||||||
| Euglenophytes and Chlorarachniophytes | |||||||||
| Multicellular red algae | |||||||||
| Unicellular red algae | |||||||||
| Haptophytes and Dinophytes | |||||||||
| Cryptophytes | |||||||||
| Glaucophytes | |||||||||
| Cyanobacteria | |||||||||
Specialized chloroplasts in C4 plants

To fix carbon dioxide into sugar molecules in the process of photosynthesis, chloroplasts use an enzyme called RuBisCO. RuBisCO has a problem—it has trouble distinguishing between carbon dioxide and oxygen, so at high oxygen concentrations, RuBisCO starts accidentally adding oxygen to sugar precursors. This has the end result of ATP energy being wasted and CO
2 being released, all with no sugar being produced. This is a big problem, since O2 is produced by the initial light reactions of photosynthesis, causing issues down the line in the Calvin cycle which uses RuBisCO. [138]
C4 plants evolved a way to solve this—by spatially separating the light reactions and the Calvin cycle. The light reactions, which store light energy in ATP and NADPH, are done in the mesophyll cells of a C4 leaf. The Calvin cycle, which uses the stored energy to make sugar using RuBisCO, is done in the bundle sheath cells, a layer of cells surrounding a vein in a leaf. [138]
As a result, chloroplasts in C4 mesophyll cells and bundle sheath cells are specialized for each stage of photosynthesis. In mesophyll cells, chloroplasts are specialized for the light reactions, so they lack RuBisCO, and have normal grana and thylakoids, [119] which they use to make ATP and NADPH, as well as oxygen. They store CO
2 in a four-carbon compound, which is why the process is called C4 photosynthesis. The four-carbon compound is then transported to the bundle sheath chloroplasts, where it drops off CO
2 and returns to the mesophyll. Bundle sheath chloroplasts do not carry out the light reactions, preventing oxygen from building up in them and disrupting RuBisCO activity. [138] Because of this, they lack thylakoids organized into grana stacks—though bundle sheath chloroplasts still have free-floating thylakoids in the stroma where they still carry out cyclic electron flow, a light-driven method of synthesizing ATP to power the Calvin cycle without generating oxygen. They lack photosystem II, and only have photosystem I—the only protein complex needed for cyclic electron flow. [119] [138] Because the job of bundle sheath chloroplasts is to carry out the Calvin cycle and make sugar, they often contain large starch grains. [119]
Both types of chloroplast contain large amounts of chloroplast peripheral reticulum, [119] which they use to get more surface area to transport stuff in and out of them. [107] [108] Mesophyll chloroplasts have a little more peripheral reticulum than bundle sheath chloroplasts. [139]
Distribution in a plant
Not all cells in a multicellular plant contain chloroplasts. All green parts of a plant contain chloroplasts—the chloroplasts, or more specifically, the chlorophyll in them are what make the photosynthetic parts of a plant green. [15] The plant cells which contain chloroplasts are usually parenchyma cells, though chloroplasts can also be found in collenchyma tissue. [140] A plant cell which contains chloroplasts is known as a chlorenchyma cell. A typical chlorenchyma cell of a land plant contains about 10 to 100 chloroplasts.

In some plants such as cacti, chloroplasts are found in the stems, [141] though in most plants, chloroplasts are concentrated in the leaves. One square millimeter of leaf tissue can contain half a million chloroplasts. [15] Within a leaf, chloroplasts are mainly found in the mesophyll layers of a leaf, and the guard cells of stomata. Palisade mesophyll cells can contain 30–70 chloroplasts per cell, while stomatal guard cells contain only around 8–15 per cell, as well as much less chlorophyll. Chloroplasts can also be found in the bundle sheath cells of a leaf, especially in C4 plants, which carry out the Calvin cycle in their bundle sheath cells. They are often absent from the epidermis of a leaf. [142]
Cellular location
Chloroplast movement

The chloroplasts of plant and algal cells can orient themselves to best suit the available light. In low-light conditions, they will spread out in a sheet—maximizing the surface area to absorb light. Under intense light, they will seek shelter by aligning in vertical columns along the plant cell’s cell wall or turning sideways so that light strikes them edge-on. This reduces exposure and protects them from photooxidative damage. [143] This ability to distribute chloroplasts so that they can take shelter behind each other or spread out may be the reason why land plants evolved to have many small chloroplasts instead of a few big ones. [144] Chloroplast movement is considered one of the most closely regulated stimulus-response systems that can be found in plants. [145] Mitochondria have also been observed to follow chloroplasts as they move. [146]
In higher plants, chloroplast movement is run by phototropins, blue light photoreceptors also responsible for plant phototropism. In some algae, mosses, ferns, and flowering plants, chloroplast movement is influenced by red light in addition to blue light, [143] though very long red wavelengths inhibit movement rather than speeding it up. Blue light generally causes chloroplasts to seek shelter, while red light draws them out to maximize light absorption. [146]
Studies of Vallisneria gigantea, an aquatic flowering plant, have shown that chloroplasts can get moving within five minutes of light exposure, though they don’t initially show any net directionality. They may move along microfilament tracks, and the fact that the microfilament mesh changes shape to form a honeycomb structure surrounding the chloroplasts after they have moved suggests that microfilaments may help to anchor chloroplasts in place. [145] [146]
Guard cell chloroplasts
Unlike most epidermal cells, the guard cells of plant stomata contain relatively well-developed chloroplasts. [142] However, exactly what they do is controversial. [147]
Plant innate immunity
Plants lack specialized immune cells—all plant cells participate in the plant immune response. Chloroplasts, along with the nucleus, cell membrane, and endoplasmic reticulum, [148] are key players in pathogen defense. Due to its role in a plant cell’s immune response, pathogens frequently target the chloroplast. [148]
Plants have two main immune responses—the hypersensitive response, in which infected cells seal themselves off and undergo programmed cell death, and systemic acquired resistance, where infected cells release signals warning the rest of the plant of a pathogen’s presence. Chloroplasts stimulate both responses by purposely damaging their photosynthetic system, producing reactive oxygen species. High levels of reactive oxygen species will cause the hypersensitive response. The reactive oxygen species also directly kill any pathogens within the cell. Lower levels of reactive oxygen species initiate systemic acquired resistance, triggering defense-molecule production in the rest of the plant. [148]
In some plants, chloroplasts are known to move closer to the infection site and the nucleus during an infection. [148]
Chloroplasts can serve as cellular sensors. After detecting stress in a cell, which might be due to a pathogen, chloroplasts begin producing molecules like salicylic acid, jasmonic acid, nitric oxide and reactive oxygen species which can serve as defense-signals. As cellular signals, reactive oxygen species are unstable molecules, so they probably don’t leave the chloroplast, but instead pass on their signal to an unknown second messenger molecule. All these molecules initiate retrograde signaling—signals from the chloroplast that regulate gene expression in the nucleus. [148]
In addition to defense signaling, chloroplasts, with the help of the peroxisomes, [149] help synthesize an important defense molecule, jasmonate. Chloroplasts synthesize all the fatty acids in a plant cell [148] [150] —linoleic acid, a fatty acid, is a precursor to jasmonate. [148]
Photosynthesis
One of the main functions of the chloroplast is its role in photosynthesis, the process by which light is transformed into chemical energy, to subsequently produce food in the form of sugars. Water (H2O) and carbon dioxide (CO2) are used in photosynthesis, and sugar and oxygen (O2) is made, using light energy. Photosynthesis is divided into two stages—the light reactions, where water is split to produce oxygen, and the dark reactions, or Calvin cycle, which builds sugar molecules from carbon dioxide. The two phases are linked by the energy carriers adenosine triphosphate (ATP) and nicotinamide adenine dinucleotide phosphate (NADP + ). [151] [152]
Light reactions

Energy carriers
ATP is the phosphorylated version of adenosine diphosphate (ADP), which stores energy in a cell and powers most cellular activities. ATP is the energized form, while ADP is the (partially) depleted form. NADP + is an electron carrier which ferries high energy electrons. In the light reactions, it gets reduced, meaning it picks up electrons, becoming NADPH.
Photophosphorylation
Like mitochondria, chloroplasts use the potential energy stored in an H+, or hydrogen ion gradient to generate ATP energy. The two photosystems capture light energy to energize electrons taken from water, and release them down an electron transport chain. The molecules between the photosystems harness the electrons’ energy to pump hydrogen ions into the thylakoid space, creating a concentration gradient, with more hydrogen ions (up to a thousand times as many) [99] inside the thylakoid system than in the stroma. The hydrogen ions in the thylakoid space then diffuse back down their concentration gradient, flowing back out into the stroma through ATP synthase. ATP synthase uses the energy from the flowing hydrogen ions to phosphorylate adenosine diphosphate into adenosine triphosphate, or ATP. [99] [153] Because chloroplast ATP synthase projects out into the stroma, the ATP is synthesized there, in position to be used in the dark reactions. [154]
NADP + reduction
Because NADP + reduction removes electrons from the electron transport chains, they must be replaced—the job of photosystem II, which splits water molecules (H2O) to obtain the electrons from its hydrogen atoms. [99] [151]
Cyclic photophosphorylation
Dark reactions
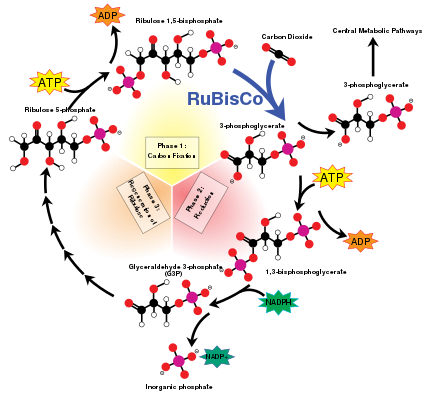
The Calvin cycle, also known as the dark reactions, is a series of biochemical reactions that fixes CO2 into G3P sugar molecules and uses the energy and electrons from the ATP and NADPH made in the light reactions. The Calvin cycle takes place in the stroma of the chloroplast. [138]
While named «the dark reactions», in most plants, they take place in the light, since the dark reactions are dependent on the products of the light reactions. [15]
Carbon fixation and G3P synthesis
The Calvin cycle starts by using the enzyme RuBisCO to fix CO2 into five-carbon Ribulose bisphosphate (RuBP) molecules. The result is unstable six-carbon molecules that immediately break down into three-carbon molecules called 3-phosphoglyceric acid, or 3-PGA. The ATP and NADPH made in the light reactions is used to convert the 3-PGA into glyceraldehyde-3-phosphate, or G3P sugar molecules. Most of the G3P molecules are recycled back into RuBP using energy from more ATP, but one out of every six produced leaves the cycle—the end product of the dark reactions. [138]
Sugars and starches

Glyceraldehyde-3-phosphate can double up to form larger sugar molecules like glucose and fructose. These molecules are processed, and from them, the still larger sucrose, a disaccharide commonly known as table sugar, is made, though this process takes place outside of the chloroplast, in the cytoplasm. [155]
Alternatively, glucose monomers in the chloroplast can be linked together to make starch, which accumulates into the starch grains found in the chloroplast. [155] Under conditions such as high atmospheric CO2 concentrations, these starch grains may grow very large, distorting the grana and thylakoids. The starch granules displace the thylakoids, but leave them intact. [156] Waterlogged roots can also cause starch buildup in the chloroplasts, possibly due to less sucrose being exported out of the chloroplast (or more accurately, the plant cell). This depletes a plant’s free phosphate supply, which indirectly stimulates chloroplast starch synthesis. [156] While linked to low photosynthesis rates, the starch grains themselves may not necessarily interfere significantly with the efficiency of photosynthesis, [157] and might simply be a side effect of another photosynthesis-depressing factor. [156]
Photorespiration
Photorespiration can occur when the oxygen concentration is too high. RuBisCO cannot distinguish between oxygen and carbon dioxide very well, so it can accidentally add O2 instead of CO2 to RuBP. This process reduces the efficiency of photosynthesis—it consumes ATP and oxygen, releases CO2, and produces no sugar. It can waste up to half the carbon fixed by the Calvin cycle. [151] Several mechanisms have evolved in different lineages that raise the carbon dioxide concentration relative to oxygen within the chloroplast, increasing the efficiency of photosynthesis. These mechanisms are called carbon dioxide concentrating mechanisms, or CCMs. These include Crassulacean acid metabolism, C4 carbon fixation, [151] and pyrenoids. Chloroplasts in C4 plants are notable as they exhibit a distinct chloroplast dimorphism.
Because of the H+ gradient across the thylakoid membrane, the interior of the thylakoid is acidic, with a pH around 4, [158] while the stroma is slightly basic, with a pH of around 8. [159] The optimal stroma pH for the Calvin cycle is 8.1, with the reaction nearly stopping when the pH falls below 7.3. [160]
CO2 in water can form carbonic acid, which can disturb the pH of isolated chloroplasts, interfering with photosynthesis, even though CO2 is used in photosynthesis. However, chloroplasts in living plant cells are not affected by this as much. [159]
Chloroplasts can pump K+ and H + ions in and out of themselves using a poorly understood light-driven transport system. [159]
In the presence of light, the pH of the thylakoid lumen can drop up to 1.5 pH units, while the pH of the stroma can rise by nearly one pH unit. [160]
Amino acid synthesis
Chloroplasts alone make almost all of a plant cell’s amino acids in their stroma [161] except the sulfur-containing ones like cysteine and methionine. [162] [163] Cysteine is made in the chloroplast (the proplastid too) but it is also synthesized in the cytosol and mitochondria, probably because it has trouble crossing membranes to get to where it is needed. [163] The chloroplast is known to make the precursors to methionine but it is unclear whether the organelle carries out the last leg of the pathway or if it happens in the cytosol. [164]
Other nitrogen compounds
Chloroplasts make all of a cell’s purines and pyrimidines—the nitrogenous bases found in DNA and RNA. [161] They also convert nitrite (NO2 − ) into ammonia (NH3) which supplies the plant with nitrogen to make its amino acids and nucleotides. [161]
Other chemical products
The plastid is the site of diverse and complex lipid synthesis in plants. [165] [166] The carbon used to form the majority of the lipid is from acetyl-CoA, which is the decarboxylation product of pyruvate. [165] Pyruvate may enter the plastid from the cytosol by passive diffusion through the membrane after production in glycolysis. [167] Pyruvate is also made in the plastid from phosphoenolpyruvate, a metabolite made in the cytosol from pyruvate or PGA. [165] Acetate in the cytosol is unavailable for lipid biosynthesis in the plastid. [168] The typical length of fatty acids produced in the plastid are 16 or 18 carbons, with 0-3 cis double bonds. [169]
The biosynthesis of fatty acids from acetyl-CoA primarily requires two enzymes. Acetyl-CoA carboxylase creates malonyl-CoA, used in both the first step and the extension steps of synthesis. Fatty acid synthase (FAS) is a large complex of enzymes and cofactors including acyl carrier protein (ACP) which holds the acyl chain as it is synthesized. The initiation of synthesis begins with the condensation of malonyl-ACP with acetyl-CoA to produce ketobutyryl-ACP. 2 reductions involving the use of NADPH and one dehydration creates butyryl-ACP. Extension of the fatty acid comes from repeated cycles of malonyl-ACP condensation, reduction, and dehydration. [165]
Other lipids are derived from the methyl-erythritol phosphate (MEP) pathway and consist of gibberelins, sterols, abscisic acid, phytol, and innumerable secondary metabolites. [165]

Chloroplasts are a special type of a plant cell organelle called a plastid, though the two terms are sometimes used interchangeably. There are many other types of plastids, which carry out various functions. All chloroplasts in a plant are descended from undifferentiated proplastids found in the zygote, [161] or fertilized egg. Proplastids are commonly found in an adult plant’s apical meristems. Chloroplasts do not normally develop from proplastids in root tip meristems [170] —instead, the formation of starch-storing amyloplasts is more common. [161]
In shoots, proplastids from shoot apical meristems can gradually develop into chloroplasts in photosynthetic leaf tissues as the leaf matures, if exposed to the required light. [12] This process involves invaginations of the inner plastid membrane, forming sheets of membrane that project into the internal stroma. These membrane sheets then fold to form thylakoids and grana. [171]
If angiosperm shoots are not exposed to the required light for chloroplast formation, proplastids may develop into an etioplast stage before becoming chloroplasts. An etioplast is a plastid that lacks chlorophyll, and has inner membrane invaginations that form a lattice of tubes in their stroma, called a prolamellar body. While etioplasts lack chlorophyll, they have a yellow chlorophyll precursor stocked. [12] Within a few minutes of light exposure, the prolamellar body begins to reorganize into stacks of thylakoids, and chlorophyll starts to be produced. This process, where the etioplast becomes a chloroplast, takes several hours. [171] Gymnosperms do not require light to form chloroplasts. [171]
Light, however, does not guarantee that a proplastid will develop into a chloroplast. Whether a proplastid develops into a chloroplast some other kind of plastid is mostly controlled by the nucleus [12] and is largely influenced by the kind of cell it resides in. [161]

Plastid interconversion
Plastid differentiation is not permanent, in fact many interconversions are possible. Chloroplasts may be converted to chromoplasts, which are pigment-filled plastids responsible for the bright colors seen in flowers and ripe fruit. Starch storing amyloplasts can also be converted to chromoplasts, and it is possible for proplastids to develop straight into chromoplasts. Chromoplasts and amyloplasts can also become chloroplasts, like what happens when a carrot or a potato is illuminated. If a plant is injured, or something else causes a plant cell to revert to a meristematic state, chloroplasts and other plastids can turn back into proplastids. Chloroplast, amyloplast, chromoplast, proplast, etc., are not absolute states—intermediate forms are common. [161]
Division
Most chloroplasts in a photosynthetic cell do not develop directly from proplastids or etioplasts. In fact, a typical shoot meristematic plant cell contains only 7–20 proplastids. These proplastids differentiate into chloroplasts, which divide to create the 30–70 chloroplasts found in a mature photosynthetic plant cell. If the cell divides, chloroplast division provides the additional chloroplasts to partition between the two daughter cells. [172]
In single-celled algae, chloroplast division is the only way new chloroplasts are formed. There is no proplastid differentiation—when an algal cell divides, its chloroplast divides along with it, and each daughter cell receives a mature chloroplast. [171]
Almost all chloroplasts in a cell divide, rather than a small group of rapidly dividing chloroplasts. [173] Chloroplasts have no definite S-phase—their DNA replication is not synchronized or limited to that of their host cells. [174] Much of what we know about chloroplast division comes from studying organisms like Arabidopsis and the red alga Cyanidioschyzon merolæ. [144]

The division process starts when the proteins FtsZ1 and FtsZ2 assemble into filaments, and with the help of a protein ARC6, form a structure called a Z-ring within the chloroplast’s stroma. [144] [175] The Min system manages the placement of the Z-ring, ensuring that the chloroplast is cleaved more or less evenly. The protein MinD prevents FtsZ from linking up and forming filaments. Another protein ARC3 may also be involved, but it is not very well understood. These proteins are active at the poles of the chloroplast, preventing Z-ring formation there, but near the center of the chloroplast, MinE inhibits them, allowing the Z-ring to form. [144]
Next, the two plastid-dividing rings, or PD rings form. The inner plastid-dividing ring is located in the inner side of the chloroplast’s inner membrane, and is formed first. [144] The outer plastid-dividing ring is found wrapped around the outer chloroplast membrane. It consists of filaments about 5 nanometers across, [144] arranged in rows 6.4 nanometers apart, and shrinks to squeeze the chloroplast. This is when chloroplast constriction begins. [175]
In a few species like Cyanidioschyzon merolæ, chloroplasts have a third plastid-dividing ring located in the chloroplast’s intermembrane space. [144] [175]
Late into the constriction phase, dynamin proteins assemble around the outer plastid-dividing ring, [175] helping provide force to squeeze the chloroplast. [144] Meanwhile, the Z-ring and the inner plastid-dividing ring break down. [175] During this stage, the many chloroplast DNA plasmids floating around in the stroma are partitioned and distributed to the two forming daughter chloroplasts. [176]
Later, the dynamins migrate under the outer plastid dividing ring, into direct contact with the chloroplast’s outer membrane, [175] to cleave the chloroplast in two daughter chloroplasts. [144]
A remnant of the outer plastid dividing ring remains floating between the two daughter chloroplasts, and a remnant of the dynamin ring remains attached to one of the daughter chloroplasts. [175]
Of the five or six rings involved in chloroplast division, only the outer plastid-dividing ring is present for the entire constriction and division phase—while the Z-ring forms first, constriction does not begin until the outer plastid-dividing ring forms. [175]

Regulation
In species of algae that contain a single chloroplast, regulation of chloroplast division is extremely important to ensure that each daughter cell receives a chloroplast—chloroplasts can’t be made from scratch. [77] [144] In organisms like plants, whose cells contain multiple chloroplasts, coordination is looser and less important. It is likely that chloroplast and cell division are somewhat synchronized, though the mechanisms for it are mostly unknown. [144]
Light has been shown to be a requirement for chloroplast division. Chloroplasts can grow and progress through some of the constriction stages under poor quality green light, but are slow to complete division—they require exposure to bright white light to complete division. Spinach leaves grown under green light have been observed to contain many large dumbbell-shaped chloroplasts. Exposure to white light can stimulate these chloroplasts to divide and reduce the population of dumbbell-shaped chloroplasts. [173] [176]
Chloroplast inheritance
Like mitochondria, chloroplasts are usually inherited from a single parent. Biparental chloroplast inheritance—where plastid genes are inherited from both parent plants—occurs in very low levels in some flowering plants. [177]
Many mechanisms prevent biparental chloroplast DNA inheritance, including selective destruction of chloroplasts or their genes within the gamete or zygote, and chloroplasts from one parent being excluded from the embryo. Parental chloroplasts can be sorted so that only one type is present in each offspring. [178]
Gymnosperms, such as pine trees, mostly pass on chloroplasts paternally, [179] while flowering plants often inherit chloroplasts maternally. [180] [181] Flowering plants were once thought to only inherit chloroplasts maternally. However, there are now many documented cases of angiosperms inheriting chloroplasts paternally. [177]
Angiosperms, which pass on chloroplasts maternally, have many ways to prevent paternal inheritance. Most of them produce sperm cells that do not contain any plastids. There are many other documented mechanisms that prevent paternal inheritance in these flowering plants, such as different rates of chloroplast replication within the embryo. [177]
Among angiosperms, paternal chloroplast inheritance is observed more often in hybrids than in offspring from parents of the same species. This suggests that incompatible hybrid genes might interfere with the mechanisms that prevent paternal inheritance. [177]
Transplastomic plants
Recently, chloroplasts have caught attention by developers of genetically modified crops. Since, in most flowering plants, chloroplasts are not inherited from the male parent, transgenes in these plastids cannot be disseminated by pollen. This makes plastid transformation a valuable tool for the creation and cultivation of genetically modified plants that are biologically contained, thus posing significantly lower environmental risks. This biological containment strategy is therefore suitable for establishing the coexistence of conventional and organic agriculture. While the reliability of this mechanism has not yet been studied for all relevant crop species, recent results in tobacco plants are promising, showing a failed containment rate of transplastomic plants at 3 in 1,000,000. [181]



