







СТРУКТУРНАЯ И ФУНКЦИОНАЛЬНАЯ РОЛЬ ГЕТЕРОХРОМАТИНА
В отличие от безуспешных попыток понять фенотипические проявления полиморфизма хромосом, молекулярная природа вариабельных участков достаточно хорошо изучена. Главное, что отличает различные сегменты хромосом — это богатая по проявлениям неоднородность их структурной и функциональной организации. Так, общим феноменом является наличие в каждой хромосоме двух типов хроматина — эухроматина и гетерохроматина. В данной работе я рассмотрю именно гетерохроматин.
Актуальность темы: Данную тему я считаю очень актуальной, так как гетерохроматин является участком хроматина, в составе которого происходит реализация генетической информации, а также репликация и репарация ДНК.
Цель: Изучить структурную и функциональную роль гетерохроматина.
Задачи: Задачи работы следующие:
1. Дать понятие гетерохроматина.
2. Изучить историю открытия гетерохроматина.
3. Рассмотреть факультативный и конститутивный гетерохроматин.
4. Выяснить особенности структуры и состава гетерохроматина.
Глава 1. Понятие «Гетерохроматин»
В составе хромосом выделяют участки гетерохроматина и эухроматин.
Гетерохроматин — участки хроматина, находящиеся в течение клеточного цикла в конденсированном (компактном) состоянии. Гетерохроматин расположен в районе центромеры, теломер и внутри плеч хромосом – интеркалярный гетерохроматин. Гетерохроматиновые участки значительно дольше представлены в клеточном цикле в виде плотно спирализованных фрагментов [4]. Они деспирализуются значительно позже, чем эухроматин или совсем не деспирализуются, сохраняясь в интерфазном ядре в виде плотно окрашенных глыбок – хромоцентров. Гетерохроматиновые районы хромосом могут ассоциироваться друг с другом – эктопическая конъюгация. В результате здесь возможна повышенная частота хромосомных перестроек. Еще одной особенностью гетерохроматина является варьирование его количества в геноме[3].
Глава 2. История открытия гетерохроматина
В 1907 году немецкий цитолог С. Гутхерц обнаружил, что некоторые фрагменты хромосом или хромосомы целиком во время клеточного деления интенсивно окрашиваются и выглядят более конденсированными по сравнению со слабоокрашенными участками. В ядрах клеток, находящихся в интерфазе, были обнаружены участки, интенсивно окрашиваемые красителями, связывающимися с хроматином, такие участки были названы хромоцентрами. С. Гутхерц показал, что гетеропикнотические сегменты хромосом становятся заметными в начале профазы, то есть в начале конденсации хромосом, отличаясь от «нормальных» участков более интенсивной окраской; различия в интенсивности окраски по мере конденсации снижаются и становятся практически неразличимыми в конце метафазы.
Другой немецкий цитолог Эмиль Хайц пришёл к выводу, что хромоцентры, обнаруживаемые в интерфазе, ассоциируются с сильно конденсированными и интенсивно окрашиваемыми гетеропикнотическими участками хромосом, наблюдаемыми в течение митотического цикла, то есть хромоцентры и гетеропикнотические участки являются одними и теми же участками хромосом, которые не подвергаются деконденсации в телофазе.
В 1928 году Хайц предложил термины «эухроматин» для участков хромосом, претерпевающих процесс компактизации-декомпактизации в процессе митоза и «гетерохроматин» для участков, постоянно остающихся в конденсированном состоянии. Хайц считал, что гетерохроматиновые участки хромосом являются генетически инертными [3].
Глава 3. Факультативный и конститутивный гетерохроматин
Гетерохроматин в хромосомах человека в свою очередь подразделяется на структурный (конститутивный) и факультативный гетерохроматин.
Конститутивный (структурный) гетерохроматин образует постоянные структурные элементы в парах гомологичных хромосом и располагается преимущественно в околоцентромерных районах, а также дистальном отделе длинного плеча Y-хромосомы. Конститутивный гетерохроматин человека состоит из сателлитной ДНК I–III классов, а также из сателлитной ДНК α-, β- и γ-типов.
Термин «факультативный гетерохроматин» относится к гетерохроматинизированному эухроматину, который присутствует не в обеих, а в одной из двух гомологичных хромосом. Если конститутивному гетерохроматину присуще неизменное, стабильное конденсированное состояние, то факультативный гетерохроматин свойственен какой-либо определенной стадии развития или типу клеток. Классическим примером факультативного гетерохроматина является функционально неактивная (инактивированная) Х-хромосома, представленная в интерфазных соматических клетках, имеющих две Х-хромосомы, в виде так называемого тельца полового хроматина (тельце Барра). Факультативный гетерохроматин, в отличие от конститутивного хроматина, содержащего различные классы сателлитной ДНК, обогащен протяженными повторяющимися последовательностями типа LINE, которые способствуют конденсации хроматина. Наличие LINE повторов в G-сегментах сближает эти районы метафазных хромосом с факультативным гетерохроматином. Гетерохроматиновые районы поздно реплицируются, метилированы и содержат гипоацетилированные гистоны, вследствие чего эти сегменты хромосом обычно транскрипционно инертны или обладают пониженной транскрипционной активностью [1].
Глава 4. Особенности структуры и состава гетерохроматина
Хроматин является нуклеопротеидом — комплексом ДНК с гистонами. Конденсация хроматина в гетерохроматин сопровождается как модификацией гистонов, так и усложнением состава нуклеопротеидного комплекса за счёт участия в нём белков гетерохроматина HP1 (Heterochromatin Protein 1). Гистоны гетерохроматинового комплекса характеризуются низкой степенью ацетилированности по лизиновым остаткам, что увеличивает их основные свойства и, соответственно, связывание с кислыми фосфатными группами ДНК, что способствует компактификации комплекса. [6]. Другой особенностью, ведущей к образованию гетерохроматина, является метилирование 27-го лизинового остатка гистона H3 белками Polycomb-комплекса 2 (PRC2) и 9-го лизинового остатка гистона H3 гистоновой метилтрансферазой Suv39h. Метилирование 9-го лизинового остатка гистона H3 ведёт к образованию высокоаффинного сайта связывания гистона H3 и белка гетерохроматина HP1. У дрозофил метилтрансфераза Suv39h функционально ассоциирована с гистондеацетилазой таким образом, что ацетилированное и метилированное состояние 9-го лизинового остатка гистона H3 являются взаимоисключающими, то есть обеспечивается единый механизм деацетилирования и метилирования гистона H3, ведущий к усилению связывания с гистоном как ДНК, так и белка гетерохроматина HP1.
Заключение
Хотя в исторической ретроспективе он изучен хуже, чем эухроматин, новые открытия заставляют считать, что гетерохроматин играет критически важную роль в организации и правильном функционировании геномов, начиная с дрожжей и кончая человеком (хотя у S. cerevisiae особая форма гетерохроматина) [2]. Его потенциальное значение подчеркивается тем фактом, что 96% генома млекопитающих состоит из некодирующих и повторяющихся последовательностей.
Список используемой литературы
1. Баранов В.С., Кузнецова Т. В. Цитогенетика эмбрионального развития человека: Научно-практические аспекты. [Текст]: СПб: Издательство Н-Л,2007.
2. Различие между эухроматином и гетерохроматином [Электронный ресурс]: Режим доступа: http://humbio.ru/humbio/epihumblu/001c4577.htm (Дата обращения 23.12.2017).
3. Коряков Д. Е., Жимулев И. Ф. Хромосомы. Структура и функции / Под ред. д.б.н. Л. В. Высоцкой. — Новосибирск: Изд-во СО РАН, 2009. — 258 с.
4. Новости науки [Электронный ресурс]: Режим доступа: http://elementy.ru/novosti_nauki/431182 (Дата обращения 23.12.2017).
5. Генетика человека. Бочков Н.П. с.90
6. Жимулев И. Ф. Общая и молекулярная генетика. — 1. — Новосибирск: Издательство Новосибирского университета, 2002. — 459 с.
Гетерохроматин


Гетерохроматин — участки хроматина, находящиеся в течение клеточного цикла в конденсированном (компактном) состоянии. Особенностью гетерохроматиновой ДНК является крайне низкая транскрибируемость.
Содержание
История открытия
Другой немецкий цитолог Эмиль Хайц, проанализировав соотношение числа хромоцентров и гетеропикнотических участков хромосом, наблюдаемых при митозе в клетках мха, пришёл к выводу, что хромоцентры, обнаруживаемые в интерфазе, ассоциируются с сильно конденсированными и интенсивно окрашиваемыми гетеропикнотическими участками хромосом, наблюдаемыми в течение митотического цикла, то есть хромоцентры и гетеропикнотические участки являются одними и теми же участками хромосом, которые не подвергаются деконденсации в телофазе.
Факультативный и конститутивный (структурный) гетерохроматин
Основное функциональное отличие факультативного гетерохроматина от конститутивного — возможность перехода в эухроматиновое состояние, при котором ДНК становится транскрипционно активной и, соответственно, происходит экспрессия генов, локализованных на данном участке хромосомы.
Факультативный гетерохроматин содержит кодирующую и, в силу этого, относительно консервативную ДНК; ДНК конститутивного гететерохроматина является преимущественно некодирующей и в силу этого высокополиморфна и вариабельна.
На ранних стадиях онтогенеза во многих случаях содержание гетерохроматина в метафазных хромосомах значительно ниже, чем на более поздних стадиях и в клетках взрослого организма — метафазные хромосомы бластомеров многих позвоночных сильно декомпактизированы, в интерфазных ядрах гетерохроматиновые образования не обнаруживаются.
Факультативный и конститутивный гетерохроматины также выявляются по различию в окрашивании: если факультативный гетерохроматин подвергается G-окрашиванию по Романовскому — Гимзе в стандартных условиях, то окраска этим же красителем после денатурации-ренатурации ДНК избирательно окрашивает конститутивный гетерохроматин. Такой селективный метод получил название окраски на конститутивный (C) гетерохроматин, или C-окрашивание.
Факультативный гетерохроматин
Обычно факультативные гетерохроматиновые участки присутствуют в только одной из гомологичных хромосом. Типичным примером факультативного гетерохроматина является неактивная половая хромосома при гомогаметном кариотипе, например, неактивная X-хромосома у женских особей млекопитающих, деактивирующаяся в конденсированное гетерохроматиновое состояние; такая гетерохроматиновая X-хромосома наблюдается в интерфазе как тельце Барра. Вместе с тем, при гаметогенезе и на ранних стадиях эмбриогенеза обе X-хромосомы являются эухроматиновыми и транскрипционно активными.
Другим примером образования факультативного гетерохроматина является пахитенная стадия мейотического деления гетерогаметного гаметоцита, сопровождающаяся у млекопитающих при сперматогенезе образованием гетерохроматинового комплекса XY-хромосом — полового пузырька. Образование такого гетерохроматинового комплекса является временным и обратимым, необходимым для деактивации половых хромосом на этой стадии мейоза: в случае, когда Х и Y хромосомы остаются активными на этой стадии, происходит нарушение баланса между продуктами экспрессии аутосом и половых хромосом, что приводит к гибели клеток.
Факультативный гетерохроматин обуславливает также «молчание» тканеспецифичных генов, переходящих в эухроматиновое состояние и экспрессирующихся только в дифференцированных клетках определённых тканей: в таких клетках активны
10 % генов — остальные гены являются инактивированными и находятся в составе факультативного гетерохроматина.
Конститутивный гетерохроматин
Конститутивный (структурный) гетерохроматин содержится в обеих гомологичных хромосомах и локализован преимущественно в экспонированных участках — центромере, теломерах, ядрышковом организаторе. ДНК конституивного гетерохроматина является преимущественно сателлитной ДНК, состоящей из тандемных повторов (например, HS1 (Human Satellite 1), HS2, HS3, альфа-сателлит и другие сателлиты человека). В интерфазном ядре конститутивный гетерохроматин образует хромоцентры с внутренней стороны ядерной мембраны, а также в районах ядрышковых организаторов. Вопрос о функциональной роли структурного гетерохроматина в эукариотической клетке остается открытым.
Особенности структуры и состава гетерохроматина
Хроматин является нуклеопротеидом — комплексом ДНК с гистонами. Конденсация хроматина в гетерохроматин сопровождается как модификацией гистонов, так и усложнением состава нуклеопротеидного комплекса за счёт участия в нём белков гетерохроматина HP1 (Heterochromatin Protein 1).
Гистоны гетерохроматинового комплекса характеризуются низкой степенью ацетилированности по лизиновым остаткам, что увеличивает их основные свойства и, соответственно, связывание с кислыми фосфатными группами ДНК, что способствует компактификации комплекса. Другой особенностью, ведущей к образованию гетерохроматина, является метилирование 27-го лизинового остатка гистона H3 белками Polycomb-комплекса 2 (PRC2) и 9-го лизинового остатка гистона H3 гистоновой метилтрансферазой Suv39h. Метилирование 9-го лизинового остатка гистона H3 ведёт к образованию высокоаффинного сайта связывания гистона H3 и белка гетерохроматина HP1. У дрозофил метилтрансфераза Suv39h функционально ассоциирована с гистондеацетилазой таким образом, что ацетилированное и метилированное состояние 9-го лизинового остатка гистона H3 являются взаимоисключающими, то есть обеспечивается единый механизм деацетилирования и метилирования гистона H3, ведущий к усилению связывания с гистоном как ДНК, так и белка гетерохроматина HP1.
Что такое структурный гетерохроматин
Слово «хромосома» означает «окрашенное тело» (окрашивается более эффективно, чем остальные компоненты клетки). Каждая хромосома состоит из очень длинной молекулы ДНК, связанной с РНК и белками, образующей вещество, известное как хроматин. Во время интерфазы клеточного цикла (когда клетка не делится) хромосомы распределены по всему ядру, в то время как при митозе и мейозе они уплотняются, располагаясь ближе друг к другу.
ДНК содержится именно в хромосомах, вероятно, потому, что это облегчает разделение полного набора генов между дочерними клетками на стадии митоза, а также её упаковку в головки сперматозоидов после мейоза.
Способность к окрашиванию используют в диагностических целях для визуального исследования хромосом, их идентификации и выявления патологий. Различают слабоокрашенные части (эухроматин) и области с более выраженной окраской (гетерохроматин).
Генетическая информация (геном) заключена в молекуле ДНК и кодирована определённой последовательностью азотистых оснований в ДНК. Основная часть этой информации находится в составе хромосом в ядре клетки, однако небольшое количество ДНК в виде свободных цепочек локализовано в митохондриях цитоплазмы. Ядра присутствуют практически во всех клетках организма человека, за исключением эритроцитов и клеток хрусталика.
Обычно в каждом ядре содержится двухметровая цепь ДНК, разделённая между 23 парами хромосом (примерно по 4 см на хромосому). Однако непосредственно перед делением она уплотняется до 5 um (0,005 мм) посредством сложных процессов спирализации и уплотнения.
Структура хроматина хромосом
В каждой хромосоме цепь ДНК дважды обмотана вокруг октамерного комплекса, состоящего из восьми «белков хранения» (гистонов), формирующих нуклеосомы, при этом образованная структура напоминает «бусы на нити». Кора (сердцевина) нуклеосомы состоит из двух молекул каждого из четырех гистонов— Н2А, Н2В, НЗ и Н4. Гистоны заряжены положительно, а потому могут образовывать ионные связи с отрицательно заряженными фосфатными группами ДНК.
Последовательность аминокислот в гистонах у всех видов совпадает практически на 100%, указывая на огромную значимость данных элементов в поддержании структуры и функций хроматина. Каждая нуклеосома вмещает порядка 200 пар оснований молекулы ДНК и укорачивает длину цепи ДНК на одну десятую.
Затем подобная бусам нить спирализуется в соленоид, или зигзагообразную спираль (фибрилла длиной 30 нм), состоящую из 5—6 нуклеосом. При этом её структура поддерживается одной молекулой гистона HI на каждую нуклео-сому. Коэффициент упаковки при образовании соленоида примерно равен 5, что в свою очередь даёт общий коэффициент упаковки 50. Существует предположение, что в таком состоянии эухроматин находится во время интерфазы в местах, где гены не экспрессируются.
Во время митоза и мейоза хромосомы ещё больше уплотняются, при этом коэффициент упаковки по отношению к предыдущим уровням составляет 100, а общий коэффициент может достигать 5000. Считают, что фибриллы хроматина складываются в вытянутые петли, в основании которых расположен скаффолд (остов) из негистоновых хромосомных белков, которые прикрепляются к определённым последовательностям оснований, рассредоточенным вдоль молекулы ДНК. Сжатие этих негистоновых хромосомных белков предположительно обусловливает уплотнение хромосом.
Один из ключевых белков скаффолда — топоизомераза II, фермент, который расщепляет двойные цепи ДНК, переносит их через разрыв и вновь соединяет, что в свою очередь необходимо для релаксации суперспиралей ДНК во время репликации или транскрипции. Топоизомераза II прикрепляется к участкам связывания со скаффолдом, которые богаты А- и Т-основаниями (более 65% оснований составляют А и Т, см. главу 4). Считают, что каждая петля выступает в роли независимого функционального домена во время репликации или транскрипции ДНК.
На следующем этапе петли фибрилл хроматина спирализуются, в результате чего возникает наиболее уплотнённый гетерохроматин, входящий в состав хромосомы во время деления клеток.
Дифференциальное окрашивание хромосом
Наиболее уплотнённые части хромосом хорошо окрашиваются красителем Гимзы (G-окраска). При этом отчётливо видны сильно уплотнённые маленькие петли (G-полосы), так как участки связывания со скаффолдом тесно прилегают друг к другу. Их репликация происходит во время S-периода, они не активны во время транскрипции. Полосы, которые плохо окрашиваются раствором Гимзы, или R-полосы, состоят из более свободных петель, богатых Г- и Ц-основаниями, они отличаются высокой активностью во время транскрипции. В основе идентификации хромосом лежат различия в их окраске.
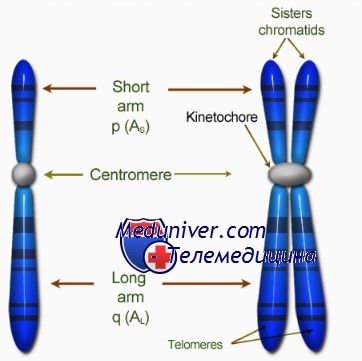
Центромера хромосом
Видимые на ранних стадиях митоза хромосомы состоят из двух абсолютно одинаковых структур, называемых сестринскими хроматидами, которые соединяет первичная перетяжка. Она представляет собой недуплицированный участок ДНК, или центромеру, которая дуплицируется в начале анафазы митоза.
Кинетохор — органелла, которая локализована на каждой стороне каждой центромеры в период ранней профазы митоза и облегчает полимеризацию димеров тубулина, необходимую для образования микротрубочек митотического веретена (веретена деления).
Тепомера хромосом
Теломерой называют специализированный участок конца хромосомы. К ней прикрепляются теломерспецифические белки, образующие «шапочку» (cap) для защиты конца хромосомы.
Теломеры предположительно препятствуют патологическому слиянию концов хромосом конец в конец, обеспечивают полноту репликации, участвуют в образовании пар хромосом во время мейоза и помогают восстанавливать внутреннюю структуру ядра на стадии интерфазы путём присоединения хромосом к ядерной мембране.
Эухроматин и гетерохроматин хромосом
Эухроматин находится в уплотнённом состоянии во время деления клеток и распаковывается на стадии интерфазы. Он обусловливает бледное окрашивание R-полос в уплотнённых хромосомах и содержит большинство структурных генов.
Гетерохроматин очень сильно уплотнён во время деления клетки и остаётся в таком состоянии даже на стадии интерфазы. Он локализован в основном на периферии ядра и около ядрышка и не активен во время транскрипции. Конструктивный (структурный) гетерохроматин одинаков во всех клетках организма, в то время как факультативный гетерохроматин различен в зависимости от типа генов, экспрессируемых в дифференцированных клетках определённых тканей.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Научная электронная библиотека

Юров И. Ю., Ворсанова С. Г., Воинова В. Ю., Чурносов М. И., Юров Ю. Б.,
3.2. Гетерохроматин и эухроматин
Каждая хромосома человека дифференцирована на два типа различных участков: так называемые, эухроматиновые и гетерохроматиновые районы хромосом. Существует два основных типа хроматина: гетерохроматин и эухроматин. Данная классификация основана на ранних изучениях свойств хроматина хромосом человека в интерфазных ядрах, содержащих деконденсированный хроматин (эухроматин), распределенный диффузно и слабо окрашиваемый при использовании специфических гистологических красителей, а также конденсированные темно-окрашенные блоки хроматина (гетерохроматин). В эухроматиновых участках хромосом локализовано подавляющее большинство генов, в то время как гетерохроматин содержит мало генов, которые, по-видимому, обычно не активны. Выделяют два основных типа гетерохроматина: конститутивный и факультативный. Конститутивный гетерохроматин, который практически на протяжении всего клеточного цикла конденсирован, генетически не активен. Вероятно, последовательности ДНК, из которых состоит конститутивный гетерохроматин, практически не содержат генов. Этот тип гетерохроматина сформирован особыми классами высокоповторяющихся (сателлитных) последовательностей ДНК и расположен в участках центромер всех хромосом, а также в некоторых других ее участках. Факультативный гетерохроматин представляет собой инактивированные и конденсированные участки хромосом, которые могут обратимо переходить из эухроматинового в гетерохроматиновое состояние в зависимости от типа клетки, стадии клеточного цикла или особенности одной из гомологичных хромосом. Как правило, этот термин применяется для описания процесса инактивации хромосомы Х в клетках, которые содержат две и более хромосомы Х.
Что такое структурный гетерохроматин
• Гетерохроматин формируется на специфических последовательностях, и неактивная структура распространяется по фибрилле хроматина
• Гены в области гетерохроматина находятся в неактивном состоянии
• Поскольку длина неактивного региона варьирует от клетки к клетке, инактивация генов в этой области обусловливает эффект положения мозаичного типа
• Аналогичные эффекты распространения проявляются на теломерах и на молчащих кассетах, контролирующих тип спаривания у дрожжей
Интерфазное ядро содержит как эу-, так и гетерохроматин. Степень конденсации гетерохроматина почти такая же, как в митотических хромосомах. Гетерохроматин не проявляет активности. В интерфазе он остается в конденсированном состоянии, транскрипция в нем подавлена, репликация приходится на позднюю S-фазу, и чаще всего он локализован на периферии ядра. Гетерохроматин центромеры обычно содержит сателлитную ДНК.
Однако образование гетерохроматина не зависит от первичной структуры ДНК. Когда ген перемещается за счет хромосомной транслокации или трансфекции и интегрируется в другое положение, ближе к гетерохроматину, в результате попадания на новое место он может инактивироваться. При этом предполагается, что он приобретает черты гетерохроматинового гена.
 Мозаичный эффект положения, проявляющийся в изменении цвета глаз,
Мозаичный эффект положения, проявляющийся в изменении цвета глаз,
возникает, когда белый ген интегрируется рядом с гетерохроматиновым участком.
Клетки, в которых белый ген неактивен, образуют бляшки белоглазости, а клетки с активным белым геном образуют красные бляшки.
Выраженность эффекта определяется близостью интегрированного гена к гетерохроматиновой области.
Такая инактивация является результатом эпигенетического эффекта. Для отдельных клеток эукариот он может проявляться по-разному и приводить к эффекту положения мозаичного типа (PEV), при котором генетически идентичные клетки обладают различным фенотипом. Этот эффект хорошо изучен у Drosophila. иллюстрирует эффект положения на примере окраски глаза дрозофилы, некоторые фасетки которого не имеют окраски, в то время как другие окрашены в красный цвет.
Это объясняется тем, что белая окраска вызвана геном, который в некоторых клетках инактивировался под действием примыкающего гетерохроматина, в то время как в других остался активным.
На рисунке ниже представлено схематическое объяснение этого эффекта. Инактивация распространяется на разное расстояние от гетерохроматина в примыкающие области. В одних клетках она заходит достаточно далеко для того, чтобы инактивировать соседний ген, в других клетках этого не наблюдается. Это происходит в определенный момент эмбрионального развития, после чего состояние гена передается всем клеткам-потомкам. Потомки, произошедшие от клеток с инактивированным геном, образуют фасетки с фенотипом, отражающим выпадение признака (в данном случае неокрашенные, т. е. белого цвета).

Чем ближе располагается ген к гетерохроматиновой области, тем выше вероятность того, что он будет инактивирован. Это позволяет предполагать, что образование гетерохроматина представляет собой двухэтапный процесс: вначале на специфической последовательности происходит нуклеация, а затем неактивное состояние распространяется по фибрилле хроматина.
Расстояние, на которое оно распространяется, точно не известно; скорее всего, процесс имеет стохастическую природу, и на него влияют такие параметры, как количество белковых компонентов, играющих ограничительную роль. К числу факторов, влияющих на процесс распространения, относится активация промоторов в определенной области; активный промотор может ингибировать распространение гетерохроматина.
Гены, расположенные поблизости от гетерохроматиновых областей, находятся в неактивном состоянии и поэтому в большинстве клеток неактивны. Эта модель предполагает, что границы гетерохроматиновой области могут меняться в зависимости от присутствия необходимых белков.
У дрожжей известен т. н. эффект теломерного молчания, аналогичный эффекту положения мозаичного типа у Drosophila: гены, локализованные в области тело-меры, в различной степени инактивированы. Это объясняется эффектом распространения гетерохроматина от теломеры.
Для дрожжей характерна еще одна форма эффекта молчания. Тип спаривания у них определяется активностью одного локуса (МАТ), однако в геноме содержатся две копии таких последовательностей (HML и HMR), которые находятся в неактивной конфигурации. Молчащие локусы HML и HMR имеют много общего с гетерохроматином, и их можно рассматривать как его небольшие области.
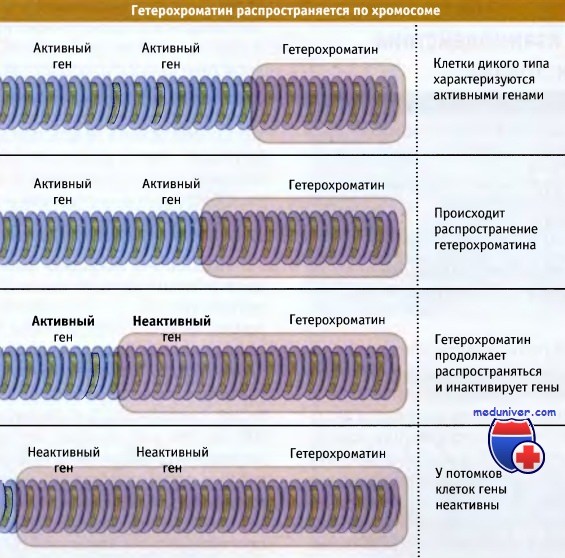 Распространение участка гетерохроматина приводит к инактивации генов.
Распространение участка гетерохроматина приводит к инактивации генов.
Вероятность инактивации гена определяется его расстоянием от гетерохроматиновой области.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021



