На пути к разгадке тайны мейоза

Появление мейоза – особого варианта клеточного деления, в результате которого число хромосом сокращается вдвое – было одним из важнейших эволюционных «достижений» первых эукариот. Механизм мейоза сложился из комбинации готовых «блоков»: механизмов митоза, рекомбинации и репарации ДНК. Ключевым событием стало формирование синаптонемного комплекса – особой белковой структуры, обеспечивающей попарное соединение и точное «выравнивание» хромосом.
Появление эукариотической клетки было важнейшим эволюционным преобразованием (ароморфозом) в истории земной жизни (см. обзор «Происхождение эукариот» ). Одним из главных «достижений» древних эукариотических организмов стало возникновение настоящего полового процесса, то есть слияния двух гаплоидных (содержащих одинарный набор хромосом) клеток – гамет в диплоидную (содержащую двойной набор хромосом) клетку – зиготу.
Чтобы жизненный цикл эукариот, обладающих половым процессом, мог продолжаться, должен был развиться механизм, посредством которого из диплоидных клеток снова могли образовываться гаплоидные. Таким механизмом стал мейоз – особый вид клеточного деления, при котором число хромосом в дочерних клетках уменьшается вдвое по сравнению с родительской клеткой.
В статье Ю.Ф.Богданова, крупнейшего специалиста по эволюции мейоза, рассматриваются современные представления о происхождении механизмов мейотического деления у эукариот.
Общепризнано, что мейоз произошел из митоза – «обычного» клеточного деления эукариот, в результате которого число хромосом остается прежним. Происхождение митоза само по себе было замечательным ароморфозом. Мейоз, по сути дела, является модифицированной версией митоза.

В ходе митоза (а также в ходе второго деления мейоза) хромосомы вступают в метафазу поодиночке, и нити веретена деления присоединяются к каждой хромосоме с двух сторон.
Именно благодаря указанным особенностям первого деления мейоза и обеспечивается уменьшение числа хромосом: к полюсам клетки расходятся не сестринские хроматиды, а гомологичные хромосомы, по одной из каждой пары (см. схему).
Эти ключевые особенности первого деления мейоза развились на основе одного и того же ароморфоза – возникновения так называемого синаптонемного комплекса (СК).
Механизм мейоза был «собран» из готовых блоков (как это часто бывает в эволюции) – разумеется, с некоторыми модификациями и новшествами.
Одним из этих исходных блоков стал уже имевшийся к тому времени у эукариот механизм митоза, другим – механизм репарации (починки повреждений) ДНК, основанный на гомологичной рекомбинации (см.: Глазер В.М. Гомологичная генетическая рекомбинация ). Суть процесса в том, что поврежденный участок одной молекулы ДНК заменяется его неповрежденной копией, взятой из другой (гомологичной) молекулы ДНК.
Чтобы «развести» гомологичные хромосомы к разным полюсам клетки, в ходе мейоза используется веретено деления, «унаследованное» от митоза. Но чтобы распределение хромосом по дочерним клеткам прошло без ошибок, гомологичные хромосомы необходимо сначала сгруппировать попарно. Для этого используется механизм гомологичной рекомбинации, точнее, одна из деталей этого механизма – образование так называемого соединения Холлидея.

Починка разрывов ДНК – одна из основ мейоза
Механизмы репарации и рекомбинации ДНК – очень древние, они должны были возникнуть еще на заре жизни.
Становление мейоза в ходе эволюции, так же как и сам мейоз в жизненном цикле современных организмов, начинается с репарации повреждений (разрывов) ДНК. Разрывы эти в начале мейоза возникают не сами собой – их создает специальный фермент, эндонуклеаза SPO11. Этот фермент является модификацией другого белка – ДНК-топоизомеразы VI, унаследованной эукариотами от своих прокариотических предков – архей.
Клетка начинает «чинить» разорванные молекулы ДНК при помощи древнего механизма гомологичной рекомбинации. А для этого нужно объединить попарно гомологичные молекулы ДНК (чтобы использовать неповрежденные участки одной молекулы в качестве «матрицы» для исправления повреждений в другой молекуле). На этом и основано попарное объединение хромосом в профазе I мейоза.
Белковые оси хромосом
Эукариотический геном значительно больше прокариотического и содержит больше «избыточной» ДНК (различных некодирующих последовательностей, мобильных генетических элементов и др.) Большие размеры генома стимулировали «распадение» его на отдельные хромосомы (линейные, в отличие от единственной кольцевой хромосомы прокариот). Как следствие, «оказалась полезной новация в структуре хромосом – белковые оси, к которым стали крепиться петли гигантской молекулы ДНК – эукариотической хромосомы».
В клетках эукариот, делящихся путем митоза, сестринские хроматиды (образовавшиеся в результате репликации исходной молекулы ДНК) соединены посредством белков-когезинов. Чтобы хроматиды могли разойтись к полюсам клетки (в анафазе митоза), когезины должны гидролизоваться и исчезнуть из межхроматидного пространства.
Один из когезинов (Rec8) «оказался подходящим субстратом для наслоения на него других мейоз-специфичных белков», из которых в начале мейоза строятся более сложные белковые оси, или «линейные элементы», к которым ДНК крепится петлями, приобретая вид «ершика». Каждая петля содержит несколько тысяч пар нуклеотидов и крепится к линейному элементу только в одном месте, «кнопочно». Такая организация хромосомы позволяет хромосомным локусам легче находить гомологичные им локусы в хромосоме-партнере. Так обстоит дело у некоторых низших эукариот, у которых нет «классического» мейоза, характерного для более высокоорганизованных форм.
Синаптонемные комплексы. Ароморфоз.
У эукариот с классическим мейозом «бывшие линейные элементы хромосом соединяются попарно с помощью белковой «застежки-молнии» в двухосевую структуру, именуемую синаптонемным комплексом (СК)». Это обеспечивает тесное соединение гомологичных хромосом. Ю.Ф.Богданов считает появление СК важным ароморфозом. СК существует ограниченное время в течение профазы I мейоза и затем распадается. СК обнаружен у многих сотен видов – от одноклеточных водорослей, грибов и протистов до многоклеточных. Предполагается несколько возможных функций СК:
1) Организация профазной мейотической хромосомы. Благодаря СК возникает билатеральная организация пары соединившихся гомологичных хромосом. Петли хроматина располагаются по обе стороны от СК. Это позволяет точно сопоставить петли хроматина в трехмерном пространстве клеточного ядра. «По-видимому, это простейший, если не единственный, способ внести порядок и точность в процесс взаимного узнавания локусов гомологичных хромосом и обеспечить достаточную точность (гомологию) рекомбинации в мейозе.»
2) СК не только соединяет гомологичные хромосомы, но и не дает им «склеиться», удерживая их на расстоянии 70-120 нм друг от друга. В конце профазы I (стадия диплотены) СК распадается во всех локусах, кроме локусов хиазм (перекрещивания хроматид). Гомологичные хромосомы, взаимно оттолкнувшиеся во всех локусах, кроме локусов хиазм, выстраиваются на экваторе веретена деления в метафазе I. Они готовы разойтись к полюсам, как только освободятся от хиазм.
3) СК необходим для формирования хиазм. СК также не позволяет хиазмам располагаться слишком близко друг от друга. Иными словами, благодаря СК хроматиды могут «рваться» и обмениваться участками лишь в ограниченном количестве мест. Ю.Ф.Богданов оценивает это как положительное явление, поскольку «чрезмерная частота кроссинговера – не столько «благо» комбинаторики, сколько угроза стабильности адаптационно выгодных фенотипов».
Происхождение белков СК
У разных групп эукариот СК строится по единому «плану», но при этом используются совершенно разные (структурно негомологичные) белки. Это похоже на постройку домов по сходному плану (стены, крыша, окна), но из совершенно разных материалов. Материал не важен – «важно, чтобы СК выравнивал параллельно лежащие гомологичные хромосомы, сохранял между ними пространство, в котором происходит рекомбинация ДНК, и сохранял бы эти условия столь долго, сколько необходимо для завершения рекомбинации и формирования хиазм».


Сравнительный анализ структуры белков СК показал, что они возникли независимо у предков современных эукариот, давших начало разным крупным таксонам (растений, грибов, животных).
Ю.Ф.Богданов приводит убедительные доводы и экспериментальные факты, свидетельствующие о том, что структуры СК формируются путем самосборки из белковых молекул, подобно многим другим сложным молекулярным комплексам. Автор отмечает, что «самоорганизующиеся структуры могут состоять из разных белковых субъединиц и нуклеиновых кислот, как это наблюдается в отношении вирусов и рибосом». Важно, что белки, сходные по размерам и трехмерной организации, могут служить компонентами одинаковых структур у далеких друг от друга организмов даже в тех случаях, когда первичная структура (аминокислотная последовательность) этих белков сильно различается.
Почему в первом делении мейоза сестринские хроматиды не расходятся?
Итак, причина нерасхождения хроматид в мейозе I – сохранение соединяющей их когезиновой оси. В результате после первого деления мейоза «программа» клеточного деления остается как бы «не выполненной». Эта программа – в основе которой лежит древняя программа митоза – после завершения мейоза I, не останавливаясь, продолжает работать (хроматиды не разошлись, когезиновые оси не гидролизовались – значит, нужно делиться дальше). И поэтому клетки сразу же, без интерфазы и без синтеза ДНК, вступают в новое деление (мейоз II), идущее уже в точности по схеме классического митоза.
В заключительнй части статьи Ю.Ф.Богданов рассматривает ряд интересных случаев отхода некоторых организмов от схемы «классического» мейоза. Например, у самцов дрозофил (в отличие от самок) СК не образуется, и мейоз идет по примитивной схеме, характерной для некоторых архаичных эукариот (дрожжи S. pombe ). Таким образом «генетическая программа» примитивного мейоза у высших организмов не исчезает (вероятно, она просто входит в состав более сложной программы «классического» мейоза) и может вновь оказаться востребованной.
Ю. Ф. Богданов. Белковые механизмы мейоза (популярная статья в журнале «Природа»).
С фактами и теориями, касающимися происхождения мейоза, можно ознакомиться в статьях Ю.Ф.Богданова и его коллег:
Научная электронная библиотека

Юров И. Ю., Ворсанова С. Г., Воинова В. Ю., Чурносов М. И., Юров Ю. Б.,
1.1. Хромосомы человека
Хромосомный набор человека, определяемый как кариотип – совокупность данных о структуре, размерах и количестве митотических хромосом, – установлен в начале 60-х годов прошлого века. Ещё в 1888 году Г. Вальдеер (H. Waldeyer, 1836–1921 гг.) ввёл термин «хромосома» для обозначения окрашенных нитевидных структур, видимых в ходе стадий деления клетки (митоза). Характерные особенности строения каждой хромосомы человека определяются, как известно, положением в ней центромеры – важнейшей структуры, которая в делении клетки (митозе) соединяется с нитями веретена и определяет расхождение сестринских хроматид к противоположным полюсам клетки. Метафазная хромосома состоит из двух хроматид (сестринские хроматиды) и центромеры, при помощи которой они соединяются. В районе центромеры хромосома сужена, две её хроматиды сближены, и этот район в теле хромосомы образует первичную перетяжку. Центромера делит хромосому на два плеча (короткое и длинное). По положению центромеры и первичной перетяжки среди хромосом человека различают метацентрические хромосомы, у которых центромера расположена в середине хромосомы (медианно) и делит её на два равных по длине плеча; субметацентрические хромосомы, в которых центромера расположена субмедианно и делит хромосому на два плеча неравной длины; и акроцентрические хромосомы, у которых центромера расположена почти на конце хромосомы (терминально), отделяя от длинного очень короткое плечо. У некоторых хромосом на коротком плече двух хроматид на красящейся тонкой нити располагаются маленькие хроматические тельца – спутники. Участки на концах хромосомы называются теломерами. Структуры в виде точек прикрепления нитей митотического веретена к центромерам называются кинетохорами. Плечи некоторых хромосом содержат перетяжки, называемые «вторичными» (например, хромосомы 1, 9, 16). Диплоидный набор человека, состоящий из 46 хромосом, составлен из 23 пар гомологичных хромосом – гомологов (отцовского и материнского происхождения): 22 пары аутосом и плюс половые хромосомы (гоносомы) – ХХ у женщин или ХY у мужчин. Гомологичные хромосомы, как правило, сходны между собой в размерах и строении, хотя могут встречаться некоторые отклонения от каждого показателя, и это носит название «гетероморфизм хромосом». Термин «кариотип» рекомендуется применять к систематизированному набору хромосом отдельной клетки человека. Существует также термин «идиограмма», который сохраняется для представления кариотипа в виде схемы, построенной на основании измерений хромосом большого числа клеток. Хромосомы пронумерованы серийно от 1 до 22 в соответствии с их длиной, а также с другими особенностями их строения, допускающими идентификацию. Половые хромосомы (гоносомы) не имеют номеров и обозначаются как Х и Y. Следует отметить, что термины и «кариотип», и «идиограмма», получившие международное признание и распространение, принадлежат русским цитологам: «идиограмма» – С.Г. Навашину (1857–1930 гг.) в 1921 году и «кариотип» – Г.А. Левитскому (1878–1942 гг.) в 1924 году. В фазах деления – метафазах и прометафазах (см ниже) – хромосомы можно увидеть в световом микроскопе как дискретные удлинённые структуры длиной от 2 до 11 мкм. На рисунках 1 и 2 представлены мужской и женский кариотипы человека.

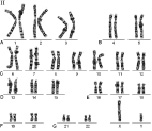
Рис. 1. Мужской кариотип: I – метафазная пластинка; II – классификация по группам и нумерация хромосом
Как указано выше, кариотип человека состоит из 46 хромосом, которые нумеруются от 1 до 22 (аутосомы) и делятся на 7 групп, – A, B, C, D, E, F, G и половые хромосомы (гоносомы) X и Y (рис. 1 и 2).
К первой группе А относятся хромосомы 1, 2 и 3, которые хорошо отличаются друг от друга. Хромосома 1 (размер – 11 мкм) – метацентрическая, содержит вторичную перетяжку в околоцентромерном участке длинного плеча. Хромосома 2 (10,8 мкм) по размерам почти равна хромосоме 1 и является субметацентрической. Хромосома 3 (размер – 8,3 мкм) – практически метацентрическая.
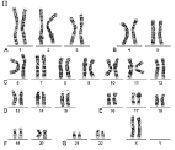
Рис. 2. Женский кариотип: I – метафазная пластинка; II – классификация по группам и нумерация хромосом
К группе В относятся хромосомы 4 и 5 (размер – 7,7 мкм каждая) – это крупные субметацентрические хромосомы, которые не отличаются друг от друга при рутинном окрашивании ни размером, ни положением центромер.
К группе С относятся хромосомы с 6 по 12 и Х. В основном, это субметацентрические хромосомы крупных и средних размеров. Наиболее крупные хромосомы из группы С – 6, 7 и Х (6,8–7,2 мкм). Хромосома Х является половой хромосомой (гоносомой). Хромосома 7 более метацентрична, чем хромосома 6. Хромосомы 8 и 9 – практически одинаковы по размеру (5,8 мкм). Хромосома 8 метацентричнее хромосомы 9, которая характеризуется регулярной вторичной перетяжкой в прицентромерном районе длинного плеча.
К группе D относятся хромосомы 13, 14 и 15 (4,2 мкм) – средних размеров акроцентрические хромосомы с почти терминальным расположением центромеры. Эти хромосомы между собой не различаются ни по размерам, ни морфологически после рутинного окрашивания. Короткое плечо всех трёх пар хромосом может формировать спутники (рис. 1, 2 и 5).
К группе Е относятся хромосомы 16, 17 и 18. Хромосома 16 (размер – 3,6 мкм) – сравнительно небольшая метацентрическая хромосома, содержащая вторичную перетяжку в длинном плече. Хромосома 17 (размер – 3,5 мкм) – сравнительно короткая субметацентрическая хромосома. Хромосома 18 (размер – 3,2 мкм) – самая короткая субметацентрическая хромосома.
Группа F представлена хромосомами 19 и 20 (размеры – 2,9 мкм). Это короткие метацентрические хромосомы, которые не отличаются между собой без дифференциального окрашивания по длине.
Хромосомы 21 и 22 (2,8 мкм) относятся к группе G. Это наиболее короткие акроцентрические хромосомы в кариотипе, которые обладают способностью формировать спутники на коротком плече (рис. 1, 2 и 5).
Хромосома Y (2,3 мкм) является маленькой акроцентрической хромосомой, сравнимой по размерам с хромосомами 21 и 22, но не имеющая спутников.
Важнейшая работа по созданию общей системы обозначения и классификации хромосом человека, представляемая в виде отдельной книги, проводилась регулярно из года в год, куда в последние десятилетия включались и результаты использования современных молекулярно-цитогенетических методов. Первые результаты работы учёных по номенклатуре и классификации хромосом человека были представлены в 1960 году в Денвере, в Университете штата Колорадо (США). Работа комиссии была проведена при поддержке американского онкологического общества. В состав комиссии входили известнейшие учёные: 14 учёных-цитологов и 3 учёных-генетика. Результаты были опубликованы в Денвере в том же 1960 году в ряде журналов в виде документа (книги), названного как «Стандартная система номенклатуры митотических хромосом человека». Впоследствии был создан комитет по номенклатуре хромосом человека. Комиссия и комитет периодически собирались для работы и, по мере развития цитогенетических, а в последние десятилетия и молекулярно-цитогенетических исследований, вносили поправки и дополнения в существующую классификацию. Работа комиссий проводилась регулярно в различных странах и городах. Комитет последовательно собирался в Париже, Чикаго, Мехико, Лэйк-Плэсиде, Эдинбурге, Стокгольме, где было решено разработать и опубликовать унифицированный вариант номенклатуры хромосом человека, включающий основные положения первых совещаний (Денвер, Лондон, Чикаго, Париж). Результаты работы комитета публиковались в различных издательствах. Этот важный документ получил название «Международная система номенклатуры хромосом человека» – «An International System for Human Cytogenetic Nomenclature» (ISCN). В дальнейшем поправки и дополнения в документе делались, учитывая новые разработанные технологии, включая молекулярно-цитогенетические, и докладывались на различных конференциях. Последний документ – «ISCN 2016» – An International System for Human Cytogenetic Nomenclature» опубликован в 2016 году. Таким образом, в одном документе даётся полная номенклатура хромосом в норме и при хромосомных синдромах и аномалиях, исходя из цитогенетических и современных молекулярно-цитогенетических технологий (см рекомендуемую литературу).
Следует отметить, что хромосомные синдромы и аномалии связаны с хромосомными (геномными) мутациями (аномалиями) в виде различных структурных перестроек хромосом или с изменением их числа (n). Численные изменения хромосом могут быть двух типов: полиплоидии – умножение хромосомного набора (3n, 4n и т. д.) или генома, кратное гаплоидному числу хромосом; анеуплоидии – увеличение или уменьшение числа хромосом, некратное гаплоидному. Структурные хромосомные (геномные) перестройки классифицируют по принципу линейной последовательности расположения генов: делеции (потеря хромосомных участков), дупликации (удвоение хромосомных участков), инверсии (перевертывание на 180° относительно нормальной последовательности хромосомных участков), инсерции (вставки хромосомных участков) и транслокации (изменение расположения хромосомных участков). Подробная информация по возможным аномалиям хромосом человека представлена в главе 3.5.
Что такое сестринские хроматиды
БУЗ ВО «Вологодская областная клиническая больница», Вологда
Центр лечения бесплодия «ЭКО», Москва
Центр репродукции и генетики «ФертиМед», Москва
Возрастные изменения и риск хромосомных аномалий в ооцитах человека (обзор литературы)
Журнал: Проблемы репродукции. 2019;25(2): 16-26
Смирнова А. А., Зыряева Н. А., Аншина М. Б. Возрастные изменения и риск хромосомных аномалий в ооцитах человека (обзор литературы). Проблемы репродукции. 2019;25(2):16-26.
Smirnova A A, Zyriaeva N A, Anshina M B. Age-related changes and risk of chromosomal incompetence in human oocytes (literature review). Russian Journal of Human Reproduction. 2019;25(2):16-26.
https://doi.org/10.17116/repro20192502116
БУЗ ВО «Вологодская областная клиническая больница», Вологда

В обзоре представлены данные молекулярной биологии об особенностях мейоза у женщин, обусловливающих повышение частоты анеуплоидии в ооцитах с возрастом. Установлено, что анеуплоидными являются около 30% всех зрелых ооцитов и только 1—2% сперматозоидов. Согласно результатам исследований, частота анеуплоидии увеличивается пропорционально возрасту женщины и достигает 82% у женщин старше 42 лет. Основной особенностью оогенеза является длительный период времени между остановкой мейоза во внутриутробном периоде и возобновлением его в каждом цикле, начиная с пубертатного периода и до менопаузы. Рассмотрена ведущая причина возрастной анеуплоидии в ооцитах — ослабление сцепления сестринских хроматид. Показаны независимые от возраста причины анеуплоидии в ооцитах: ошибки контрольных точек формирования веретена в ооцитах, нестабильность веретена деления, сниженная частота и проблемные позиции гомологичной рекомбинации.
БУЗ ВО «Вологодская областная клиническая больница», Вологда
Центр лечения бесплодия «ЭКО», Москва
Центр репродукции и генетики «ФертиМед», Москва
В последние десятилетия наблюдается стойкая тенденция к повышению возраста деторождения. Если 20 лет назад возраст женщины при рождении первого ребенка составлял около 25 лет, то сейчас у многих женщин первые роды происходят между 30 и 35 годами [1]. Например, в 2013 г. в США 43% живорождений пришлось на возраст женщины 30 лет и старше [2]. При этом, по меркам классического акушерства, женщину, рожающую первого ребенка после 28 лет, называли «старородящей» или «пожилой первородящей» и относили ее к группе повышенного риска развития осложнений беременности и родов.
В настоящее время многие женщины рожают детей в возрасте 40 лет и старше. Существует отчетливая тенденция к снижению частоты наступления беременности, повышению риска ее прерывания и увеличению вероятности рождения детей с хромосомной патологией у женщин старшего репродуктивного возраста по сравнению с молодыми [3]. В связи с этим возникает вопрос о том, как именно возраст женщины влияет на качество половых клеток и здоровье будущего ребенка.
Одной из ведущих причин спонтанного прерывания беременности и врожденных пороков развития плода являются генетические нарушения, возникающие вследствие ошибок расхождения хромосом во время мейотического деления ооцитов [4]. Ооциты более подвержены ошибкам, чем сперматоциты и делящиеся путем митоза соматические клетки [5]. Большинство анеуплоидий, обнаруживаемых у эмбрионов человека, происходят из яйцеклетки, а не из сперматозоида [4, 6—10]. Известно, что 88% трисомий по21-й хромосоме имеют материнское происхождение, 8% — отцовское и 4% возникают вследствие ошибок митоза на ранних стадиях развития эмбриона [11]. Установлено, что анеуплоидными являются около 30% ооцитов и только 1—2% сперматозоидов [11], вероятно, из-за значительных различий в процессе мейоза у мужчин и женщин. У мужчин сперматогенез начинается после пубертатного периода и продолжается в течение всей жизни. Напротив, у женщин мейоз инициируется во время внутриутробного развития плода, останавливается на стадии профазы I перед рождением девочки, возобновляется только в процессе овуляции во взрослом возрасте и происходит циклически вплоть до менопаузы [4].
Ограниченный резерв ооцитов формируется еще до рождения девочки, при этом их качество с возрастом снижается, что приводит к снижению фертильности и прогрессивному увеличению числа яйцеклеток с аномальным количеством хромосом [5].
Анеуплоидия — явление, при котором клетки организма содержат число хромосом, некратное гаплоидному (одинарному). Анеуплоидии делят на трисомии (2n+1), тетрасомии (2n+2), двойные трисомии (2n+1+1), моносомии (2n—1) и нуллисомии (2n—2) [12].
Аутосомные моносомии обычно погибают до клинически установленной беременности, трисомии обусловливают тяжелые аномалии развития плода и в 30% случаев приводят к самопроизвольному патологическому прерыванию беременности. Некоторые аутосомные трисомии и анеуплоидии по половым хромосомам совместимы с жизнью, самая частая из них — трисомия по 21-й хромосоме (синдром Дауна) [11, 13]. Частота трисомии эмбриона при клинически установленной беременности остается низкой у женщин в возрасте 20 лет (около 2—3%) и увеличивается примерно до 35% в возрасте 40 лет [14].
Внутриутробное развитие ооцитов
Внутриутробно до 6—7-й недели эмбрионального развития в первичную гонаду из области основания желточного мешка мигрируют половые клетки гоноциты. С окончанием этого периода заканчивается индифферентная стадия развития гонады. Половая дифференцировка индуцируется половыми хромосомами. Формирование гонады женского типа начинается с 8—10-й недели внутриутробного развития. Первичные половые клетки превращаются в оогонии, которые затем размножаются путем митотического деления (рис. 1).  Рис. 1. Развитие ооцита [5]. С началом мейоза оогонии получают название ооцитов (ооцит 1-го порядка). Оогонии содержат диплоидный хромосомный набор (46ХХ), в процессе мейоза происходит его редукция до гаплоидного (23Х). Вокруг ооцитов из клеток мезенхимы образуются первичные гранулезные клетки. На 20-й неделе развития у плода начинается процесс образования примордиальных фолликулов, которые содержат ооцит, плотно окруженный клетками эпителия (см. рис. 1). К рождению девочки число примордиальных фолликулов составляет около одного миллиона [15]. Мейоз начинается со стадии репликации (удвоения) ДНК, которое происходит в интерфазу (периоды G1 — пресинтетический, S — синтетический, G2 — постсинтетический) [16], после чего следуют 2 цикла деления клеток — 1-е и 2-е деления мейоза (мейоз I и мейоз II), в процессе которых происходит редукция числа хромосом и образуются гаплоидные гаметы.
Рис. 1. Развитие ооцита [5]. С началом мейоза оогонии получают название ооцитов (ооцит 1-го порядка). Оогонии содержат диплоидный хромосомный набор (46ХХ), в процессе мейоза происходит его редукция до гаплоидного (23Х). Вокруг ооцитов из клеток мезенхимы образуются первичные гранулезные клетки. На 20-й неделе развития у плода начинается процесс образования примордиальных фолликулов, которые содержат ооцит, плотно окруженный клетками эпителия (см. рис. 1). К рождению девочки число примордиальных фолликулов составляет около одного миллиона [15]. Мейоз начинается со стадии репликации (удвоения) ДНК, которое происходит в интерфазу (периоды G1 — пресинтетический, S — синтетический, G2 — постсинтетический) [16], после чего следуют 2 цикла деления клеток — 1-е и 2-е деления мейоза (мейоз I и мейоз II), в процессе которых происходит редукция числа хромосом и образуются гаплоидные гаметы.
Мейотическое деление состоит из следующих фаз: профаза I (включает 5 стадий: лептотена, зиготена, пахитена, диплотена, диакинез), метафаза I, анафаза I, телофаза I, затем профаза II, метафаза II, анафаза II, телофаза II [16]. Лептотена (стадия тонких нитей) — начало конденсации хромосом. Зиготена (стадия сливающихся нитей) — сближение и начало конъюгации гомологичных хромосом; к концу ее все гомологи объединяются в биваленты (рис. 2).  Рис. 2. Бивалент у женщин молодого и старшего возраста [5]. а — бивалент в ооцитах у молодых женщин. Достаточное количество когезина и шугошина. Гомологичные хромосомы хорошо сцеплены между собой с помощью дистального когезина, сестринские хроматиды также хорошо сцеплены между собой с помощью проксимального когезина. Шугошин защищает проксимальный когезин от гидролиза сепаразой. Кинетохоры сестринских хроматид ориентированы вместе; б — бивалент в ооцитах у женщин старшего возраста. Потеря проксимального когезина и шугошина ведет к разделению сестринских кинетохоров, что может привести к преждевременному разделению сестринских хроматид. Потеря дистального когезина приводит к преждевременному расщеплению бивалента (на униваленты). В пахитене (стадия толстых нитей) происходит кроссинговер. Диплотена (стадия двойных нитей) начинается взаимным отталкиванием гомологов, которые остаются связанными в области хиазм; хромосомы деконденсируются и приобретают вид «ламповых щеток». Это наиболее длительный период профазы I. У человека хромосомы типа «ламповых щеток» существуют 12—50 лет, т. е. остаются в таком состоянии до полового созревания женского организма и далее до менопаузы [12]. В связи с этим выделяют диктиотену — форму диплотены профазы I деления мейоза, характеризующуюся существенно большей продолжительностью в связи с прохождением этапа вителлогенеза (процесса синтеза и накопления питательных веществ в ооцитах на этапе их быстрого роста). На этой стадии происходит остановка развития в профазе I мейоза (см. рис. 1) — 1-й блок мейоза на стадии диктиотены.
Рис. 2. Бивалент у женщин молодого и старшего возраста [5]. а — бивалент в ооцитах у молодых женщин. Достаточное количество когезина и шугошина. Гомологичные хромосомы хорошо сцеплены между собой с помощью дистального когезина, сестринские хроматиды также хорошо сцеплены между собой с помощью проксимального когезина. Шугошин защищает проксимальный когезин от гидролиза сепаразой. Кинетохоры сестринских хроматид ориентированы вместе; б — бивалент в ооцитах у женщин старшего возраста. Потеря проксимального когезина и шугошина ведет к разделению сестринских кинетохоров, что может привести к преждевременному разделению сестринских хроматид. Потеря дистального когезина приводит к преждевременному расщеплению бивалента (на униваленты). В пахитене (стадия толстых нитей) происходит кроссинговер. Диплотена (стадия двойных нитей) начинается взаимным отталкиванием гомологов, которые остаются связанными в области хиазм; хромосомы деконденсируются и приобретают вид «ламповых щеток». Это наиболее длительный период профазы I. У человека хромосомы типа «ламповых щеток» существуют 12—50 лет, т. е. остаются в таком состоянии до полового созревания женского организма и далее до менопаузы [12]. В связи с этим выделяют диктиотену — форму диплотены профазы I деления мейоза, характеризующуюся существенно большей продолжительностью в связи с прохождением этапа вителлогенеза (процесса синтеза и накопления питательных веществ в ооцитах на этапе их быстрого роста). На этой стадии происходит остановка развития в профазе I мейоза (см. рис. 1) — 1-й блок мейоза на стадии диктиотены.
В профазе I происходит конъюгация гомологичных хромосом (синапсис) и рекомбинация (обмен ДНК между гомологичными хромосомами) [4]. Этот процесс называют гомологичной рекомбинацией (см. рис. 1) [17]. Связи между гомологичными хромосомами устанавливаются на ранних стадиях развития ооцита во время роста плода женского пола [17]. Материнская и отцовская хромосомы соединяются с помощью белкового синаптонемного комплекса, в результате чего происходит кроссинговер — процесс обмена гомологичными участками ДНК между гомологичными хромосомами [5, 18]. После завершения кроссинговера комплекс исчезает, гомологичные хромосомы прочно связываются перекрещенными молекулами ДНК (хиазмами). Синаптонемный комплекс состоит из центрального элемента и двух латеральных элементов. Основа протяженных латеральных элементов синаптонемного комплекса — структура из 4 белков когезинов (от англ. cohesion — сцепление) [18].
Накануне мейоза в хромосомах появляется специфический белок когезин REC8, к нему присоединяются 3 других белка когезина. Этот когезиновый комплекс располагается внутри хромосомы между двумя сестринскими хроматидами, удерживая их вместе. С комплексом когезинов связываются мейоз-специфичные белки, которые становятся главными белками хромосомных осей и превращают эти оси в латеральные элементы синаптонемного комплекса [18—20].
Когезин склеивает сестринские хроматиды между собой в области центромер (кинетохоров) и в области мест кроссинговера (проксимальный когезин) (см. рис. 2) [5, 18—20]. Когезин дистальнее мест кроссинговера (дистальный когезин) связывает гомологичные хромосомы [5, 19, 20]. В районе кинетохора когезины защищены белком шугошином (от японского слова — защита), который препятствует гидролизу когезина REC8 сепаразой [5, 18, 21, 22]. Протеины шугошина локализуются в обеих перицентромерных областях сестринских хроматид [5, 23].
Когезины — белки, которые играют ключевую роль в процессе сцепления и расхождения хромосом во время деления клетки, регулируют процесс разделения сестринских хроматид. Центромера — участок хромосомы, связывающий сестринские хроматиды. Кинетохор — белковая структура на хромосоме, к которой крепятся волокна веретена деления, формируется в области центромеры (см. рис. 2). Кинетохоры сестринских хроматид должны функционировать как единый кинетохор, чтобы обе сестринские хроматиды гомологичной хромосомы отошли к одному полюсу в процессе мейоза I.
Таким образом, в профазе I образуется структура из двух связанных гомологичных хромосом, называемая бивалентом (см. рис. 1, 2). Затем ооцит вступает в состояние клеточного покоя, происходит остановка развития — 1-й блок мейоза на стадии диктиотены, которая у человека может длиться десятки лет [5].
Развитие ооцитов от пубертатного периода до менопаузы
Ооцит 2-го порядка останавливается на стадии метафазы II, в таком состоянии он во время овуляции выходит из зрелого фолликула в брюшную полость, затем транспортируется в маточную трубу, где происходит встреча со сперматозоидом [5]. Оплодотворение снимает 2-й блок мейоза; 2-е мейотическое деление завершается после проникновения сперматозоида в ооцит [24].
В процессе анафазы II и телофазы II один набор сестринских хроматид остается в яйцеклетке, другой набор уходит во второе полярное тело (ПТ2). ДНК яйцеклетки и сперматозоида формируется в женский и мужской пронуклеусы, которые затем сливаются друг с другом перед первым митотическим делением эмбриона (см. рис. 1) [24, 25]. Данный процесс происходит регулярно в каждом овуляторном менструальном цикле (до остановки развития ооцита на стадии метафазы II, а завершается лишь в случае оплодотворения).
Во время 1-го деления мейоза один набор гомологичных хромосом остается в ооците, а второй уходит в ПТ1. При этом сестринские хроматиды остаются связанными когезином проксимально у центромеры, а когезин защищен протеинами шугошина от воздействия сепаразы. Во 2-м делении мейоза протеины шугошина больше не защищают когезин от воздействия сепаразы, которая гидролизует когезин REC8, поэтому сестринские хроматиды разделяются и расходятся в разные стороны [23]. Таким образом, в процессе 1-го деления мейоза расходятся гомологичные хромосомы, а во время 2-го деления мейоза — сестринские хроматиды.
Причины нарушения расхождения хромосом
В качестве причин нарушений расхождения хромосом в мейозе рассматривают 2 основных механизма: нерасхождение и преждевременное разделение сестринских хроматид (рис. 3)  Рис. 3. Ошибки расхождения хромосом [4, 5]. а — нормальное расхождение хромосом (б, в, г, д — материнские ошибки в первом делении мейоза; е — материнские ошибки во втором делении мейоза); б — ошибочное прикрепление кинетохоров сестринских хроматид к микротрубочкам веретена деления. Полуперевернутый бивалент: одна пара сестринских хроматид прикрепляется к микротрубочкам противоположных полюсов веретена; в — ошибочное прикрепление кинетохоров сестринских хроматид к микротрубочкам веретена деления. Обе пары сестринских хроматид прикрепляются к микротрубочкам одного полюса веретена, что обусловливает нерасхождение гомологичных хромосом в первом делении мейоза; г — полностью перевернутый бивалент: обе пары сестринских хроматид прикрепляются к микротрубочкам противоположных полюсов веретена. В процессе второго деления мейоза возможно реципрокное расхождение; д — бивалент распадается на униваленты, которые расходятся независимо друг от друга. Возможно реципрокное расхождение; е — нормальное расхождение в первом делении мейоза. Ошибочное прикрепление сестринских хроматид к микротрубочкам одного полюса веретена во втором делении мейоза, в результате чего сестринские хроматиды не расходятся. ПРСХ — преждевременное разделение сестринских хроматид; ПТ1 — первое полярное тело; ПТ2 — второе полярное тело. [22, 26—30].
Рис. 3. Ошибки расхождения хромосом [4, 5]. а — нормальное расхождение хромосом (б, в, г, д — материнские ошибки в первом делении мейоза; е — материнские ошибки во втором делении мейоза); б — ошибочное прикрепление кинетохоров сестринских хроматид к микротрубочкам веретена деления. Полуперевернутый бивалент: одна пара сестринских хроматид прикрепляется к микротрубочкам противоположных полюсов веретена; в — ошибочное прикрепление кинетохоров сестринских хроматид к микротрубочкам веретена деления. Обе пары сестринских хроматид прикрепляются к микротрубочкам одного полюса веретена, что обусловливает нерасхождение гомологичных хромосом в первом делении мейоза; г — полностью перевернутый бивалент: обе пары сестринских хроматид прикрепляются к микротрубочкам противоположных полюсов веретена. В процессе второго деления мейоза возможно реципрокное расхождение; д — бивалент распадается на униваленты, которые расходятся независимо друг от друга. Возможно реципрокное расхождение; е — нормальное расхождение в первом делении мейоза. Ошибочное прикрепление сестринских хроматид к микротрубочкам одного полюса веретена во втором делении мейоза, в результате чего сестринские хроматиды не расходятся. ПРСХ — преждевременное разделение сестринских хроматид; ПТ1 — первое полярное тело; ПТ2 — второе полярное тело. [22, 26—30].
При нерасхождении не происходит расхождения гомологичных хромосом или сестринских хроматид в соответствующих фазах мейоза. При преждевременном разделении пары сестринских хроматид отделяются одна от другой, чтобы независимо друг от друга случайным образом, часто неправильно, разойтись во время анафазы 1-го мейотического деления. Результаты недавних цитогенетических исследований полярных тел позволяют предположить, что ошибки, обусловленные преждевременным разделением сестринских хроматид, встречаются чаще, чем обусловленные их нерасхождением [31—33].
Ошибки расхождения происходят с одинаковой частотой в 1-м и 2-м делениях мейоза, хотя некоторые исследователи сообщают о более высокой частоте ошибок в мейозе II [31, 33, 34]. Это можно объяснить тем, что часто ошибки в мейозе I проявляются только в мейозе II (см. рис. 3). Например, преждевременно разделенные сестринские хроматиды могут разойтись правильно в мейозе I, но дать ошибку позже, в мейозе II. Интересно, что ошибки, обусловленные преждевременным разделением сестринских хроматид, в мейозе I могут быть скорректированы «сбалансированной ошибкой» в мейозе II: если в ПТ1 и ПТ2 есть реципрокные ошибки, то полученный эмбрион будет содержать правильный набор хромосом [31].
Феномен «реципрокного расхождения» — формирование нормальных гаплоидных ооцитов после преждевременного разделения сестринских хроматид в мейозе I и коррекции путем расхождения гомологов в мейозе II (см. рис. 3). Реципрокное расхождение происходит, когда сестринские хроматиды, а не гомологичные хромосомы, расходятся в мейозе I. Они остаются несвязанными после мейоза I, что может вызвать проблемы в метафазе II. Реципрокное расхождение чаще встречается у женщин старшего возраста [35].
По мнению T. Chiang и соавт., большинство трисомий материнского происхождения возникают вследствие ошибок в мейозе I [4]. На материнское происхождение хромосомных аномалий в процессе мейоза I также указывают T. Hassold и соавт. [36, 37].
Анализ полярных тел 20 986 ооцитов методом FISH на 5 хромосом (13, 16, 18, 21, 22), проведенный A. Kuliev и соавт. [34], показал, что анеуплоидии с равной частотой возникают в процессе мейоза I и мейоза II. Изолированные нарушения в процессе мейоза I выявлены в 30,4%, в процессе мейоза II — в 39,8%, в то время как у 29,8% ооцитов аномальный хромосомный набор возник вследствие последовательных ошибок в мейозе I и в мейозе II. Это означает, что почти треть ошибок в мейозе II связана с предшествующими ошибками в мейозе I. Результаты показали, что частота анеуплоидии по 5 исследованным хромосомам увеличивается пропорционально возрасту женщины и составляет 20% в 35 лет и более 40% в 40 лет [34]. Анеуплоидии, происходящие вследствие последовательных ошибок в мейозе I и мейозе II, зависят от возраста.
Частота анеуплоидии, по результатам исследований ПТ1 и ПТ2 (соответственно, после мейоза I и II) методом CGH, составила 45,2% у женщин в возрасте 37 лет и младше, 65,5% у женщин 38—41 года и 82,2% у женщин 42 лет и старше [33]. Следовательно, при исследовании 24 хромосом частота анеуплоидии в ооцитах значительно выше, чем при исследовании 5 хромосом (рис. 4).  Рис. 4. Частота анеуплоидии в ооцитах в зависимости от возраста женщины (по данным A. Kuliev и соавт., и E. Fraguoli и соавт.).
Рис. 4. Частота анеуплоидии в ооцитах в зависимости от возраста женщины (по данным A. Kuliev и соавт., и E. Fraguoli и соавт.).
L. Gianaroli и соавт. исследовали ПТ1 ооцитов методом флюоресцентной гибридизации in situ (FISH) на 13, 15, 16, 18, 21 и 22-й хромосоме и выявили, что доля ооцитов с ошибками в мейозе I составила 39% у женщин моложе 38 лет и 58% у женщин 44 лет [38]. Таким образом, наличие длительного временного интервала между остановкой мейоза у плода и его возобновлением в период овуляции у взрослой женщины создает предпосылки для повышения частоты анеуплоидии в ооцитах у женщин старшего возраста [4].
Следует отметить, что исследование бластомеров эмбрионов на стадии дробления выявило анеуплоидии в 67,2% случаев у женщин до 38 лет и в 86,3% случаев у женщин 38 лет и старше [39, 40], при этом у женщин старше 45 лет эуплоидные эмбрионы вовсе не обнаружены [41].
Зависимые от возраста причины анеуплоидии в ооцитах
Для точного расхождения хромосом важна их правильная интеграция. С возрастом в ооцитах женщин чаще возникают следующие структурные дефекты [42, 43]:
1) сестринские кинетохоры отделяются на большие расстояния, что способствует неправильному расположению бивалентов в мейотическом веретене в мейозе I [44];
2) биваленты со сниженным дистальным сцеплением часто преждевременно распадаются на отдельные хромосомы, называемые униваленты [42].
Для обоих дефектов возможно реципрокное расхождение [5].
Частота образования унивалентов увеличивается с возрастом: у женщин 30—35 лет униваленты определяются в 10% ооцитов, а у женщин старше 35 лет — в 40% [45]. При реципрокном расхождении сестринские хроматиды уже разделены и не могут правильно расположиться в веретене метафазы II (см. рис. 3) [5]. Предполагают, что ухудшение сцепления хроматид является ведущей причиной возрастной анеуплоидии в ооцитах [4, 42]. Сцепление сестринских хроматид в ооцитах человека ослабевает с возрастом, что может привести к неправильному расположению бивалентов во время мейоза I (см. рис. 3) [44].
В норме дистальный когезин связывает вместе гомологичные хромосомы, проксимальный когезин связывает кинетохоры сестринских хроматид, а протеины шугошина защищают проксимальный когезин от удаления в анафазу I. В ооцитах женщин старшего возраста дистальный когезин может быть потерян, и гомологичные хромосомы разделены (см. рис. 2). Проксимальный когезин ослабевает, и сестринские кинетохоры отдаляются на большее расстояние [44]. Защитное влияние шугошина на когезин снижается в перицентромерных областях. В полуперевернутых бивалентах одна пара сестринских хроматид прикрепляется к микротрубочкам противоположных полюсов веретена, что приводит к несбалансированному расхождению. В полностью перевернутых бивалентах обе пары сестринских хроматид прикрепляются к микротрубочкам противоположных полюсов веретена, и это может привести к реципрокному расхождению (см. рис. 3). Также возможно нарушение соединения кинетохоров с микротрубочками [46]. Перевернутые биваленты чаще обнаруживают в ооцитах женщин старшего возраста, и это коррелирует с увеличенным расстоянием между сестринскими кинетохорами [42].
Поскольку сцепление хромосом должно сохраняться в течение довольно длительного периода времени (около 50 лет), оно может быть подвержено возрастным изменениям, приводящим к формированию анеуплоидии. В норме сцепление хромосом должно прекращаться только с началом анафазы. Дефект сцепления дистальнее мест кроссинговера может привести к смещению местоположения хиазмы (сдвиг хиазмы) или к преждевременному расхождению бивалентов в мейозе I. Уменьшение сцепления центромер может привести к преждевременному разделению сестринских хроматид в мейозе II [4]. Преобладание преждевременно разделившихся сестринских хроматид в ооцитах человека также говорит о том, что у женщин старшего возраста ослабевает или теряется их сцепление [4].
Исследования на трансгенных мышах показали, что протеины сцепления (когезины) устанавливаются на хромосомы внутриутробно в фазу S (синтетический период интерфазы) и остаются функциональными, пока возобновляется мейоз. Выявлено, что когезины формируются в избытке, с возрастом их количество снижается, и при достижении некоторого порогового уровня начинает расти частота анеуплоидий [4, 44, 47]. Иммуннофлюоресцентное окрашивание ооцитов на срезах яичников показало, что уровень мейоз-специфических когезинов REC8 и SMC1ß в диктиотене снижен у женщин старшего возраста [11, 48], при этом достоверно увеличивается расстояние между кинетохорами, чаще происходят ошибки расхождения хромосом [42, 44]. Установлена отрицательная линейная корреляция между уровнем когезинов, потерей сцепления и возрастом женщины [48]. Частота ошибок расхождения хромосом возрастает экспоненциально у женщин после 35 лет [14]. Причины падения уровня когезинов на сегодняшний день точно не установлены [11].
Независимые от возраста причины анеуплоидии в ооцитах
Хромосомные нарушения обнаруживают в 3—61% ооцитов у женщин даже моложе 30 лет [49, 50]. Предполагают следующие независимые от возраста причины анеуплоидии:
1. Ошибки контрольных точек формирования веретена (ТФВ) в ооцитах. Функция ТФВ в мейозе I — задерживать анафазу, пока все кинетохоры не прикрепились к микротрубочкам веретена [51]. Однако ооциты вступают в анафазу, несмотря на ошибки расположения хромосом [45]. Установлено, что ТФВ в ооцитах молодых мышей и мышей старшего возраста функционируют одинаково [4].
2. Наличие нескольких полюсов и нестабильность веретена деления затрудняют прикрепление хромосом к микротрубочкам и обусловливают неправильное расхождение сестринских хроматид или гомологичных хромосом [52].
3. Гомологичная рекомбинация влияет на сцепление хромосом в биваленте [11, 53]. У женщин отмечено больше локусов кроссинговера в хромосомах, чем у мужчин. Более длинные хромосомы формируют больше кроссинговеров, чем короткие. Дистальные и проксимальные кроссинговеры могут влиять на силу сцепления гомологичных хромосом и сестринских хроматид [35, 53, 54]. Синапсис и рекомбинация — две первые критические ступени мейоза — происходят в профазе I во время внутриутробного развития плода и поэтому не зависят от возраста. Возможное объяснение связи между ошибками рекомбинации и зависимой от возраста анеуплоидии может заключаться в том, что сниженная частота рекомбинаций или проблемные позиции рекомбинаций (слишком близко к теломерам или слишком близко к центромере) делают конкретные хромосомы более уязвимыми для зависимого от возраста нарушения других процессов годы спустя [4].
Поскольку ооциты млекопитающих проходят профазу мейоза на стадии внутриутробного развития, важно, чтобы сформировался по крайней мере один кроссинговер для рекомбинации между каждой парой гомологичных хромосом с целью обеспечения их сбалансированного расхождения в мейозе I [11]. При синдроме Дауна в 30% случаев материнского происхождения ошибка расхождения гомологичных хромосом возникает вследствие отсутствия кроссинговера [11]. В некоторых случаях наблюдается низкая частота рекомбинации во всем геноме независимо от возраста матери. Например, у сиблингов детей с синдромом Дауна, возникшим в результате отсутствия кроссинговера в 21 паре хромосом, обнаружена низкая частота рекомбинаций во всем геноме независимо от возраста матери на момент их рождения [11].
Регуляция частоты кроссинговера, возможно, менее строго контролируется у женщин, что потенциально может влиять на более высокую частоту анеуплоидий материнского происхождения [11]. Одним из материнских факторов могут быть варианты гена RNF212, обусловливающие низкую частоту кроссинговеров в ооцитах [11]. Для начала мейотической рекомбинации создаются разрывы двойной нити ДНК, затем набираются протеины для репарации и формируются локусы рекомбинации. Мутации или полиморфизм генов этих протеинов могут вызывать нарушение синапсов хромосом. Гены MSH4, MSH5, TEX11, HEI10, RNF212 и TRIP13 являются кандидатами в отношении регуляции материнской рекомбинации у человека и потенциальными факторами риска повышенной частоты анеуплоидии в ооцитах [11]. Позиции кроссинговера также регулируются на многих стадиях мейотической рекомбинации. На частоту и позиции кроссинговера влияют частота и позиции формирования разрыва двойной нити ДНК. Формирование кроссинговера в определенных локусах делает более вероятным нерасхождение хромосом [11].
Таким образом, можно предположить существование генетической предрасположенности к нарушениям процессов мейотического деления ооцитов, проявляющимся независимо от возраста женщины.
Вывод
1. Наличие длительного временного интервала между остановкой мейоза у плода женского пола и его возобновлением в период овуляции у взрослой женщины создает предпосылки для повышения частоты анеуплоидии в ооцитах женщин старшего возраста.
2. Ведущей причиной возрастной анеуплоидии в ооцитах считается ухудшение сцепления хроматид.
3. Независимые от возраста причины анеуплоидий в ооцитах включают ошибки контрольных точек формирования веретена в ооцитах, нестабильность веретена деления, снижение частоты кроссинговера и проблемные позиции гомологичной рекомбинации.
Концепция и дизайн — М.А., А.С., Н.З.
Написание текста — Н.З., А.С.
Редактирование — А.С., М.А.
Авторы заявляют об отсутствии конфликта интересов.
The authors declare no conflict of interest.



